
CO2浓度升高条件下不同氮素供应对闽楠幼苗光合特性及生长的影响*
2021-05-21 08:07:08韦小丽吴高殷陈胜群
林业科学 2021年4期
王 晓 韦小丽,2 吴高殷 陈胜群
(1.贵州大学林学院 贵阳 550025; 2.贵州省森林资源与环境研究中心 贵阳 550025)
CO2浓度升高是当前全球气候变化的一个主要因素(Landryetal., 2016)。CO2作为温室气体主要成分之一,预计本世纪末将达到700 μmol·mol-1(Aranjueloetal., 2009; IPCC, 2013)。CO2浓度升高在引起温室效应的同时,还影响地球表面其他环境因子变化并直接或间接影响植物生长。大多数情况下,如果没有发生长期的光合作用调节,光合速率将随CO2浓度升高而升高(Quentinetal., 2015; DaMattaetal., 2015)。然而经受长时间高浓度CO2处理的植物则表现出较大的变异性。长期高浓度CO2条件下,多种植物在原先短时间高浓度CO2下初始出现的光合速率增强会逐渐减弱或渐渐消失,有的甚至出现光合作用下调现象(Ainsworthetal., 2003; Seneweera, 2011),被称为光合适应(Alonsoetal., 2009; Gutiérrezetal., 2009),影响植物诸多生理过程,进而影响植物生长。解释植物产生CO2光合适应现象的原因有很多,其中一个重要原因是植物碳氮平衡的破坏(Seneweera, 2011; Bloometal., 2012)。
氮是植物必需的大量营养元素之一,氮素营养对光合作用在高 CO2下的适应影响取决于氮的供应水平,同时受植株整体水平的源库关系调节(Petterssonetal., 1994)。氮素供应不足会限制植物对CO2浓度升高的响应,当氮成为限制因素时,核酮糖1,5-磷酸羧化酶加氧酶(Rubisco)中的氮也可能会重新分配到其他酶类或者其他组织和器官,从而对Rubisco进行调节,这使得植物光合作用更加受到Rubisco的限制(邓亚运等, 2014)。Wujeska-Klause等(2019)的研究表明,CO2浓度升高会限制植物对氮素吸收从而影响植物对CO2浓度升高的积极响应。除了氮素供应量影响植物生长对CO2浓度升高的响应,氮素形态也是重要因素。氮素形态本身影响植物光合特性,不同氮素形态代谢表现不同,相较于铵态氮,CO2浓度升高不利于植物对硝态氮的吸收转化(Hachiyaetal., 2014; Jaureguietal., 2015; Andrewsetal., 2018)。
CO2浓度升高影响闽楠(Phoebebournei)光合作用(韩文军等, 2003),短时CO2体积分数加倍处理使植物的表观光合速率明显增加; 长期处理则明显抑制闽楠的光合作用,出现严重的光合适应现象。结合CO2浓度升高条件下闽楠幼苗出现的光合适应现象以及光合适应出现的碳氮平衡破环原因,本文以1年生闽楠实生苗为试验材料,采用开顶式气室进行CO2浓度处理,施用不同量硝态氮及铵态氮培育闽楠幼苗,旨在研究: 1)闽楠幼苗气体交换参数随CO2浓度升高处理时间延长的变化; 2)不同氮素形态和供应量是否可以调控CO2浓度升高对闽楠幼苗气体交换参数的影响; 3)CO2浓度升高条件下氮素形态及供应量的改变对闽楠幼苗光合生理、有机物积累以及生长的影响; 4)氮素供应如何影响CO2浓度升高条件下闽楠光合作用的生理基础。本研究以期为未来闽楠生长应对CO2浓度升高提供理论依据和技术指导。
1 材料与方法
1.1 试验材料 供试材料为同一母株种子培育的闽楠1年生实生容器苗, 2019年1月选取长势一致的健壮容器苗(平均苗高: 9.42±0.32 cm;地径: 2.73±0.13 mm)移栽至塑料花盆中(口径15.5 cm,高14 cm),盆栽土壤为黄壤,每盆装风干土1.5 kg,栽植幼苗1株,移栽后缓苗4个月,选取长势一致健壮盆栽幼苗(平均苗高: 16.38±1.28 cm;地径: 3.27±0.24 mm)进行试验处理。盆栽土壤养分含量: 全氮(0.84 g·kg-1)、碱解氮(28.3 mg·kg-1)、全磷(0.23 g·kg-1)、有效磷(7.8 mg·kg-1)、全钾(21.6 g·kg-1)、速效钾(207.3 mg·kg-1)。试验所用氮肥分别为硝酸钙、硫酸铵; 磷肥为过磷酸钙; 钾肥为氯化钾。

1.3 指标测定 1) 气体交换参数测定 采用Li-6400便携式光合作用测量系统,每个处理选取3株,每株选取中上部3片完全展开的功能叶并作标记,于晴天9:00—11:00测定叶片净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)叶片气体交换参数,水分利用效率(WUE)由Pn/Tr计算所得。测定时采用 CO2小钢瓶控制aCO2气室测定时叶片叶室CO2浓度350 μmol·mol-1,eCO2气室测定时叶片叶室CO2浓度为700 μmol·mol-1,利用系统自配红蓝光源补光,光量子通量密度设定为1 200 μmol·m-2s-1。
2) Rubisco和RCA活性测定 采用酶联免疫试剂盒(上海科顺生物),每个处理选取5株,每株选取中上部中等成熟的功能叶作为测定材料,按说明书方法测定。
3) 可溶性糖、淀粉、可溶性蛋白含量测定 每个处理每个重复选取5株,每株选取中上部中等成熟的功能叶作为测定材料,采用蒽酮比色法测定可溶糖和淀粉含量,采用考马斯亮蓝染色法测定可溶性蛋白含量(李合生, 2000)。
4) 苗高、地径测定 施肥处理前,每个处理每个重复选取5株进行挂牌编号标记并测定初始苗高、地径,分别用卷尺和游标卡尺测定,CO2处理88天后再次测定标记幼苗苗高、地径。
1.4 数据分析 运用 Excel 2007进行数据的录入和图表的绘制,采用 SPSS 23. 0对数据进行方差分析及差异显著性分析。
2 结果与分析

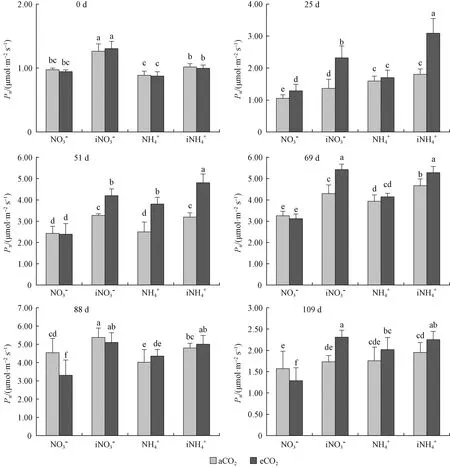
图1 不同处理对闽楠幼苗净光合速率的动态影响Fig. 1 Effect of different treatments on net photosynthetic rate of P. bournei seedlings aCO2: (350±70) μmol·mol-1 CO2; eCO2: (700±10) μmol·mol-1 CO2; 0.8 g N·seedling-1; 1.2 g N·seedling-1. 下同 The same below.
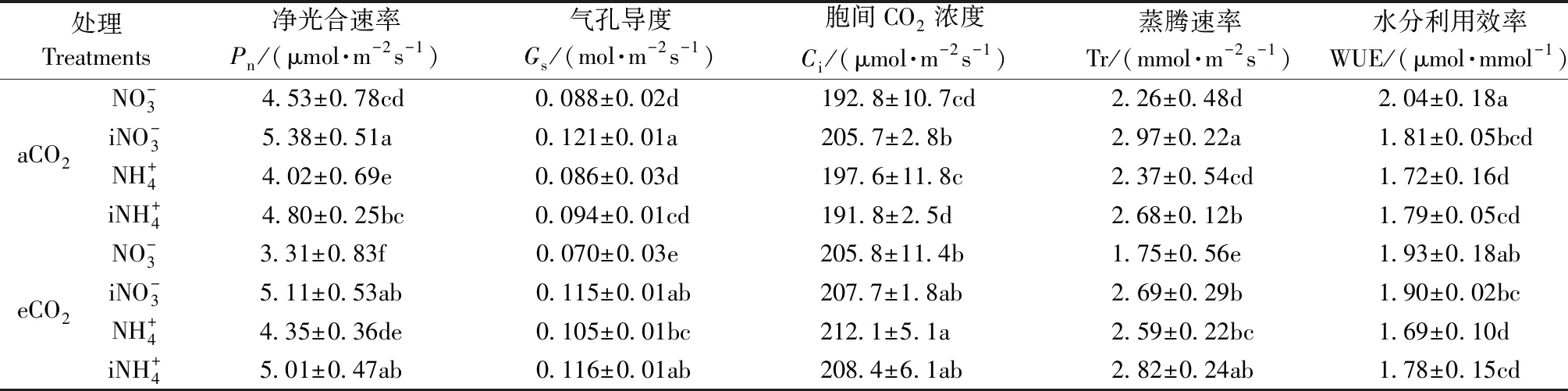
表1 不同处理对闽楠幼苗气体交换参数的影响①Tab.1 Effect of different treatments on gas exchange of P. bournei seedlings
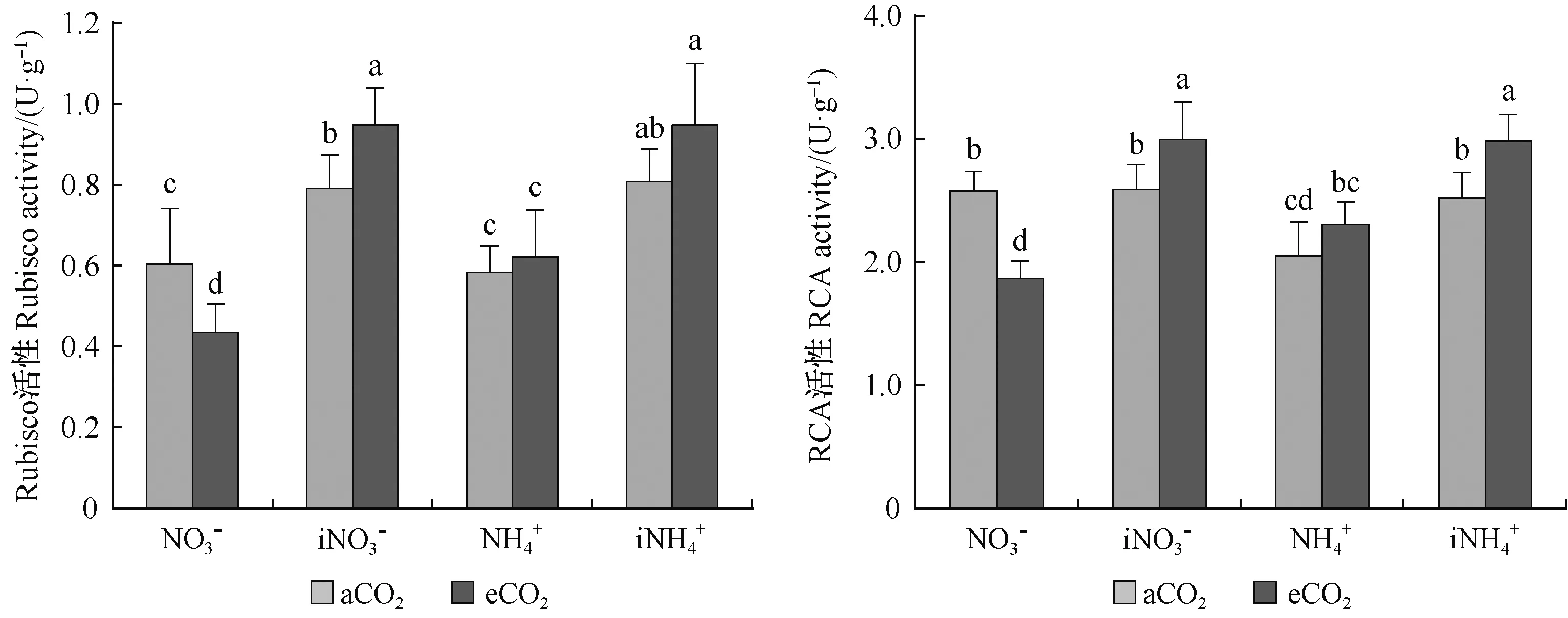
图2 不同处理对闽楠幼苗光合关键酶活性的影响Fig. 2 Effect of different treatments on activity of photosynthetic key enzyme of P. bournei seedlings
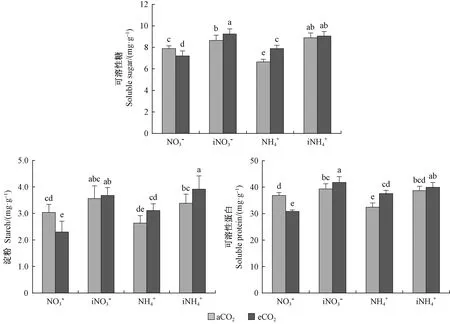
图3 不同处理对闽楠幼苗可溶性糖、淀粉、可溶性蛋白含量的影响Fig. 3 Effect of different treatments on soluble sugar, starch, soluble protein content of P. bournei seedlings

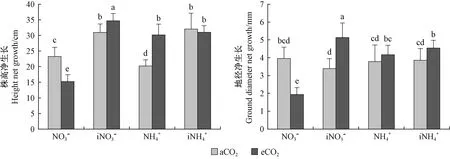
图4 不同处理对闽楠幼苗苗高、地径净生长量的影响Fig. 4 Effects of different treatments on height and ground diameter net growth of P. bournei seedlings
3 讨论
3.1 CO2浓度升高条件下不同氮素供应对闽楠幼苗光合特性的影响 植物苗期对外界环境变化较为敏感,CO2浓度升高尤其是较长时期的高CO2浓度会对苗木的生长生理带来较大影响,进而影响苗木质量(徐胜等, 2015)。闽楠播种当年苗木生长量较低,第2年生长加快,有3次抽梢,因而第2~3年的生长量大小对造林苗木质量影响很大。通常情况下,短时间CO2浓度升高会促进植物光合作用。提高净光合速率,经过短期的促进作用之后,CO2浓度升高对光合效率的积极影响逐渐减弱,甚至出现光合适应现象(Reyetal., 1998)。CO2浓度升高条件下,净光合速率变化是判断植物是否出现光合适应现象的依据之一(Longetal.,1993)。本研究结果表明,CO2浓度升高条件下施用中等量硝态氮闽楠幼苗在处理前期净光合速率逐渐增高且高于自然CO2浓度,说明CO2浓度升高短期内可促进幼苗光合作用,随着处理时间的延长,幼苗净光合速率低于自然CO2浓度条件,且处理后期气温降低,幼苗净光合速率逐渐降低的同时,也低于自然CO2浓度条件下幼苗的净光合速率,说明施用中等量硝态氮闽楠幼苗在CO2浓度升高条件中较长时间后出现光合下调现象,这与韩文军等(2003)的研究结果一致。
增施硝态氮或施用铵态氮闽楠幼苗在整个CO2浓度升高处理过程中净光合速率均高于自然CO2浓度,说明CO2浓度升高条件下增施硝态氮或施用铵态氮有利于闽楠幼苗进行光合作用,这可能是增施硝态氮可弥补碳素供应增多的氮素供应相对不足,施用铵态氮则避免CO2浓度升高对硝态氮吸收代谢的不良影响。植物高浓度CO2条件下出现光合适应现象的原因是相对于碳供应的增多,正常供应量的氮肥就会相对不足,植物碳氮供应不平衡导致光合效率降低(Seneweera, 2011; Bloometal., 2012)。除了氮素供应量的相对减少,CO2浓度升高一定程度上限制了植物对硝态氮的吸收,破坏碳氮平衡,最终导致光合适应现象出现(Smartetal., 1998; Searlesetal., 2003),其原因可能是CO2浓度升高提高Rubisco的羧化能力,抑制光呼吸,阻碍苹果酸的转运最终影响硝态氮的代谢(Rachmilevitchetal., 2004; Bloometal., 2014; Wujeska-Klauseetal., 2019),也有可能是CO2浓度升高影响植物蒸腾速率,进而影响土壤中硝态氮向根系的运输(McGrathetal., 2013; Hachiyaetal., 2014; Jaureguietal., 2015)。以铵态氮为主要氮源的生态系统,例如松林(Finzietal., 2002)、湿地(Drakeetal., 1995),CO2浓度升高使其初级净生产力提高约25%,而在以硝态氮为主要氮源的生态系统,例如草原(Shawetal., 2002),CO2浓度升高则降低其初级净生产力。Silva等(2015)的试验也指出,高浓度CO2处理期间,铵态氮处理的咖啡树(Coffeaarabica)幼苗生长优于硝态氮处理。因此植物的氮素利用相较于其他环境因素更能决定植物对CO2浓度升高的反应(Poorteretal., 2001; Dormannetal., 2002)。
3.2 CO2浓度升高条件下不同氮素供应对闽楠幼苗有机物含量及生长的影响 可溶性糖是光合作用的产物,高CO2浓度条件下植物光合速率提高,制造更多的光合产物,长期CO2浓度升高条件下,植物积累更多的光合产物,最终反馈抑制光合效率导致光合下调(Lastdrageretal., 2014; Smeekensetal., 2010)。本研究中,长期CO2浓度升高条件下出现光合下调的闽楠幼苗其可溶性糖含量较自然CO2浓度降低,说明光合下调的原因并不是光合产物积累抑制的作用,可能是由于光合关键酶活性降低影响光合效率从而影响光合产物的合成。Rubisco和RCA是光合作用的2个关键酶,其中Rubisco是光合作用C3碳反应中重要的羧化酶,是植物叶片中含量最丰富的蛋白质,植物吸收氮素的一大部分用于Rubisco的合成,氮代谢的变化影响Rubisco的活性(Makinoetal., 2003)。RCA是一种可以调节Rubisco活性的酶(Suganamietal., 2018),它能使Rubisco在植株体内条件下达到最大活化程度。光合作用效率的提高需要提高Rubisco的活性,同时也需要提高RCA对Rubisco的活化效率。Vicente等(2019)研究指出,CO2浓度升高条件下,Rubisco活性降低,Takatani等(2014)、Vicente等(2015)均观察到了CO2浓度升高条件下编码Rubisco的2个亚基(rbcS、rbcL)基因表达下调。Bokhari等(2007)研究发现,高浓度CO2条件下RCA的活性降低,同时也有研究发现CO2浓度升高条件下编码RCA的基因表达下调,最终影响光合作用(Leakeyetal., 2009; Kontunen-Soppelaetal., 2010)。本试验中,长期CO2浓度升高条件下施用中等量硝态氮,闽楠幼苗Rubisco以及RCA活性均显著降低,从而降低闽楠幼苗光合效率,影响光合产物可溶性糖、淀粉的含量,硝态氮的供应不足以及长期CO2浓度升高条件对硝态氮吸收代谢的影响使其可溶性蛋白含量降低,最终表现为闽楠幼苗苗高、地径生长的下降。增施硝态氮或施用铵态氮可改善闽楠幼苗光合关键酶活性,从而调节长期CO2浓度升高条件对闽楠幼苗光合作用的影响,最终表现为促进闽楠幼苗生长。
CO2浓度升高对植物生长的影响表现在诸多方面,本文主要从气体交换参数、光合关键酶活性、有机物含量方面研究CO2浓度升高条件下不同氮素形态及供应量对闽楠幼苗生长的调控作用,氮素供应的调控作用及相关机制更应从植物生长的碳氮代谢甚至相关基因表达方面进行研究。除了苗期,今后还应加强长期CO2浓度升高对闽楠幼树、成年树木生长及生理,细根养分周转、养分循环等的影响。另外,CO2浓度升高对植物生长的影响以及氮素对其的调控作用存在物种之间差异的同时也受植物不同生长年龄及库源关系的影响,实际生产中,CO2浓度升高条件下采用氮素对闽楠生长进行调控仍需关注闽楠苗龄以及生长环境等条件。
4 结论
长期CO2浓度升高条件下,施用中等量硝态氮肥的闽楠幼苗光合下调,影响生长;增施硝态氮肥或施用铵态氮肥可通过提高光合关键酶活性等缓解长期CO2浓度升高对闽楠幼苗光合及生长的负面影响。因此,生产上进行1年生闽楠幼苗培育时,可以考虑增加硝态氮肥或施用铵态氮肥以应对未来CO2浓度升高环境。
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
Coco薇(2016年5期)2016-06-03 09:17:41
环境污染与防治(2016年12期)2016-03-13 05:52:12
考试周刊(2016年6期)2016-03-11 08:14:32
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
植物营养与肥料学报(2011年5期)2011-11-06 07:30:52
植物营养与肥料学报(2011年2期)2011-10-26 03:52:10
植物营养与肥料学报(2011年4期)2011-10-26 02:44:56
植物营养与肥料学报(2011年3期)2011-10-24 06:15:00
植物营养与肥料学报(2010年3期)2010-11-16 00:59:24
