
小麦抗穗发芽基因挖掘及分子育种进展
2021-05-20 03:20黄义文代旭冉刘宏伟买春艳于立强刘朝辉李洪杰张宏军
麦类作物学报 2021年2期
黄义文,代旭冉,刘宏伟,杨 丽,买春艳,,于立强, 刘朝辉,李洪杰,周 阳,张宏军
(1.中国农业科学院作物科学研究所/作物分子育种国家工程实验室,北京 100081; 2.新乡矮败小麦育种技术创新中心, 河南新乡 453731; 3.石家庄市农林科学研究院赵县试验基地,河北赵县 051530)
小麦是世界第一大粮食作物,为人类提供约20%的热量和蛋白质[1-2]。中国是世界上最大的小麦生产和消费国[3]。中国产业信息网数据显示,2019年中国生产小麦1.33亿t,消费小麦 1.28亿t。据统计,从1962―2012年,中国人口增长了1倍,而小麦的消费量却增长了6倍[4]。由此可见,人口的不断增长对小麦产量的提高提出了严峻的挑战。
穗发芽是影响小麦产量和品质的重要因素。世界上许多小麦主产国如中国、美国、加拿大、法国和澳大利亚等均受到不同程度穗发芽的影响[5]。穗发芽导致小麦的价格下降20%~50%,全世界每年由于小麦穗发芽引起的经济损失高达10亿美元[6]。在中国,83%的小麦受到不同程度穗发芽的影响[7]。穗发芽对小麦的危害主要体现在三个方面:一是穗发芽降低了面团的延展性和弹性,使面团的保水能力降低,淀粉的糊化、凝胶化和回生特性改变,从而显著影响面条、蛋糕、面包的品质;二是穗发芽后籽粒中的储藏物不断被水解、消耗,导致容重和千粒重下降,严重影响小麦产量[8-10];三是穗发芽导致种子发芽势减弱,苗小苗弱,甚至造成出苗率严重下降,给农户和种子经营者造成重大经济损失。
为避免或减少小麦收获后籽粒晾晒的人工成本,近年来我国越来越多的地区要等到小麦籽粒含水量降到入库标准时才开始收获,导致收获期推迟,大大增加了穗发芽风险。因此,我国小麦生产对品种的穗发芽抗性必将提出更高的要求。本文从小麦穗发芽抗性QTLs/基因发掘、功能标记开发、穗发芽主要抗源以及抗穗发芽分子育种方面进行综述,并对今后小麦穗发芽抗性研究重点及育种思路进行讨论,以期为更有效地开展穗发芽遗传研究和指导穗发芽抗性育种提供参考。
1 已发掘的小麦穗发芽抗性位点
小麦穗发芽抗性是由多基因控制的数量性状。目前,定位到的与小麦穗发芽抗性相关的QTLs有42个,分布在除1D、4D和7D之外的18条染色体上(图1),可解释4.9%~48.3%的表型变异[5,10],其中,主效QTLs集中在3AS和4AL染色体上[5,10]。
Mares[11]应用非整倍体最早在小麦3AS染色体上发现一个与籽粒休眠有关的QTL。此后,Osa等[12]利用Zenkoujikomugi(强休眠性)/中国春(弱休眠性)RIL群体在3AS和3AL染色体上定位到两个与穗发芽抗性有关的主效QTLs (QPhs.ocs-3A.1和QPhs.ocs-3A.2),其中,QPhs.ocs-3A.2与TaVp-1基因相同。 Shao等[13]研究发现,QPhs.ocs-3A.1能够在多个环境下稳定表达,可解释23%~38%的表型变异,且在不同的抗穗发芽小麦品种中均能被检测到。Nakamura等[14]和Liu等[15]分别在红粒和白粒品种中克隆了QPhs.ocs-3A.1的候选基因TaMFT-3A(TaPHS1)。
Torada等[16]在4AL染色体上发现另一个与籽粒休眠有关的QTL,被命名为phs1。Kato等[17]利用AC Domain(强休眠性)/小麦Haruyutaka(弱休眠性)双单倍体(doubled haploid,DH)群体,在4AL染色体上定位到一个主效QTL(QPhs.ocs-4A.1),该位点在不同国家的抗穗发芽小麦品种AUS1408(南非)、SW95-50213(中国)和AUS1490(澳大利亚)中都存在,可解释30%~38%表型变异[18-19]。Torada等[16]图位克隆了phs1位点的候选基因TaMKK3-A。
2 小麦抗穗发芽基因克隆及功能标记开发
目前,已经克隆了6个与小麦穗发芽抗性相关的基因,包括TaSdr、TaMFT-3A、TaDFR、Tamyb10、TaVp-1和TaMKK3-A。检测穗发芽抗性基因不同等位基因的功能标记详细信息见表1。
2.1 TaSdr基因
Zhang等[20]在小麦2A、2B和2D染色体上克隆到3个与水稻OsSdr4同源的基因,分别命名为TaSdr-A1、TaSdr-B1和TaSdr-D1。TaSdr-A1基因在编码区+643 bp位置存在一个SNP(G/A),利用这个SNP开发的CAPS标记Sdr2A可区分TaSdr-A1a等位基因(抗穗发芽)和TaSdr-A1b等位基因(感穗发芽)[20]。在TaSdr-B1基因起始密码子上游-11 bp的位置存在一个SNP(A/G),由其开发的CAPS分子标记Sdr2B可检测等位基因TaSdr-B1a(抗穗发芽)和TaSdr-B1b(感穗发芽)[21]。TaSdr-D1基因在抗、感材料间没有序列差异。
2.2 TaMFT-3A基因
在拟南芥种子萌发过程中,MFT(MOTHER OF FT AND TFL1)基因通过调控脱落酸(ABA)和赤霉素(GA)的信号通路来调节种子的休眠[22]。Nakamura等[14]利用Zenkoujikomugi(红粒、强休眠性)/中国春(白粒、弱休眠性)F2群体将TaMFT-3A基因定位于3A染色体上,并且克隆了该基因。Liu等[15]在白粒抗穗发芽品种Rio Blanco中重复定位到QPhs.ocs-3A.1,并图位克隆其候选基因TaPHS1,还证实TaPHS1与TaMFT-3A为同一基因。TaMFT-3A基因在启动子区-222 bp处存在一个SNP(T/C),据此开发了CAPS标记MFT-3A,用来检测抗、感两个等位基因[14]。此外,Jiang等[23]在TaMFT-3A基因启动子-194 bp位置发现有一个33个碱基的插入或缺失,并与穗发芽抗性密切相关,据此开发了分子标记MFT-A2,可用来检测等位基因TaMFT-3Aa(抗穗发芽)和TaMFT-3Ab(感穗发芽)。Liu等[15]在TaPHS1编码区发现两个SNP,分别位于+646和+666 bp位置,利用这两个SNP开发了两个STS标记,并证实携带不同等位基因的材料间发芽率呈显著差异。Lei等[24]还发现TaMFT-3A基因编码区有一个12 bp的插入,且与穗发芽抗性有关,并据此开发了分子标记MFT-A1,用来检测等位基因MFT-A1a(抗穗发芽)和MFT-A1b(感穗发芽)。

加粗字体表示小麦主要抗穗发芽基因。
2.3 TaDFR基因
TaDFR基因是控制普通小麦种皮颜色的关键基因。普通小麦TaDFR有3个同源基因TaDFR-A、TaDFR-B和TaDFR-D,分别位于3A、3B和3D染色体长臂远端[25-26]。不同抗性材料间TaDFR-A、TaDFR-B和TaDFR-D基因序列比对发现,TaDFR-A和TaDFR-D基因没有等位变异,而TaDFR-B基因则存在等位变异[27]。进一步分析发现,TaDFR-B基因启动子区域有一处8 bp的插入,且与穗发芽抗性密切相关,据此开发了分子标记DFR-3B,用来检测等位基因TaDFR-Ba/c(抗穗发芽)和TaDFR-Bb(感穗发芽)[27]。
2.4 Tamyb10基因
myb10是重要的调节基因,能够激活结构基因的表达,促进色素的合成。Tamyb10基因通过影响小麦胚部对ABA的敏感性来影响小麦的休眠性[28]。Tamyb10基因有Tamyb10-A1、Tamyb10-B1和Tamyb10-D13个同源基因,分别位于3A、3B和3D染色体长臂[29]。通过序列分析发现,Tamyb10-A1和Tamyb10-B1基因分别存在3种和2种等位变异。Tamyb10-A1的3个等位基因分别为Tamyb10-A1a(665 bp)、Tamyb10-A1b(536 bp)和Tamyb10-A1a*(2 750 bp),携带不同等位基因的品种间穗发芽抗性差异不显著[30-31]。Tamyb10-B1基因第二个外显子有一处19 bp的缺失,据此开发了功能标记Tamyb10-B,可用来检测等位基因Tamyb10-B1a(感穗发芽)和Tamyb10-B1b(抗穗发芽)[31]。Wang等[32]通过比较抗、感材料中的Tamyb10-D1基因,开发出一个STS标记Tamyb10-D,并发现Tamyb10-D1a(抗穗发芽)和Tamyb10-D1b(感穗发芽)等位基因间穗发芽抗性存在极显著差异(P<0.01)。
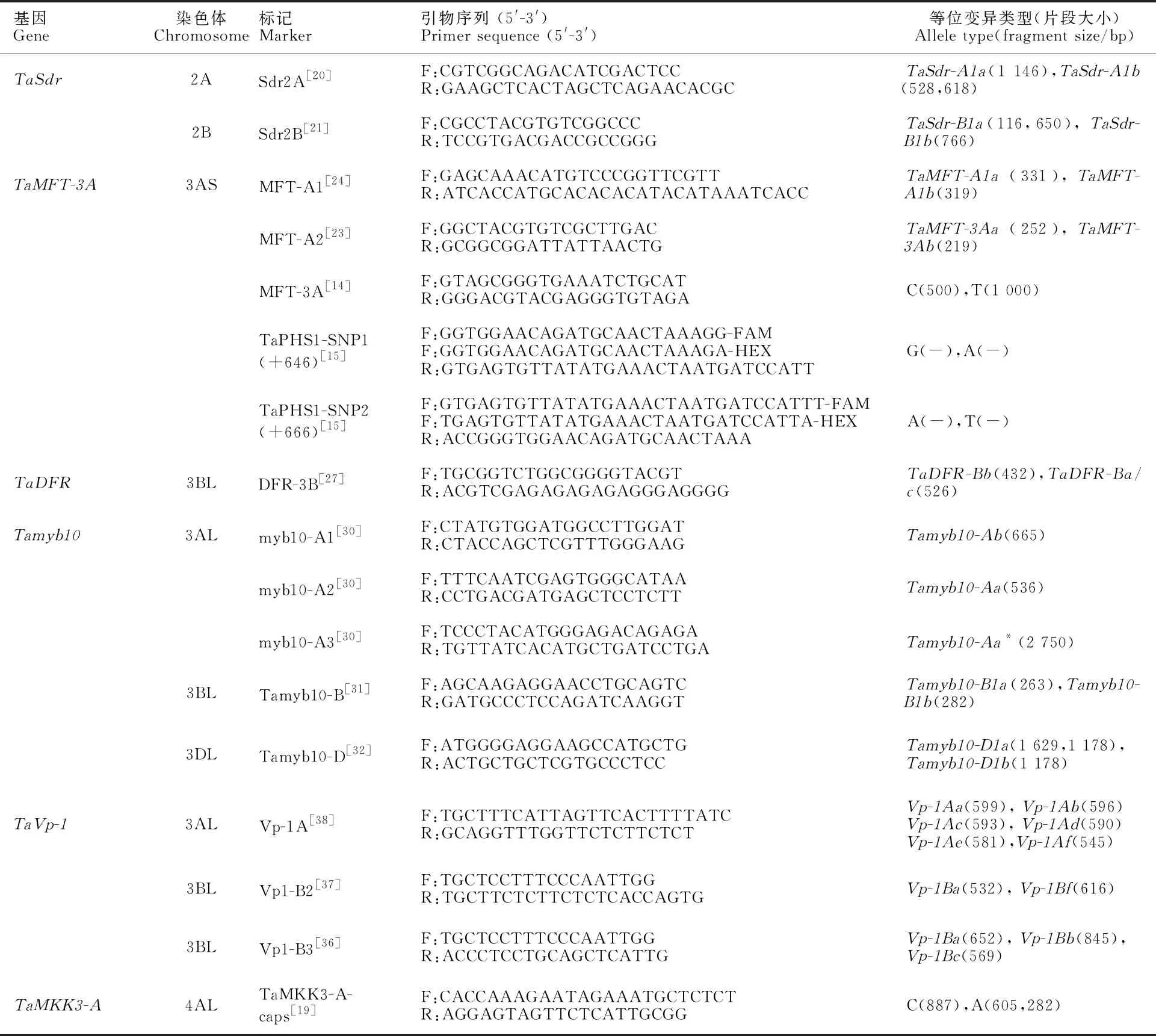
表1 小麦抗穗发芽基因相关功能标记信息Table 1 Information of functional markers for pre-harvest sprouting resistance genes in wheat
2.5 TaVp-1基因
Vp-1基因首先在玉米3号染色体上被发现[33],是促进胚成熟和休眠的主要转录调节因子[34]。小麦TaVp-1基因被分别定位在3A、3B和3D染色体长臂上[35]。目前小麦A和B基因组的Vp-1A和Vp-1B已经开发了功能标记。在中国地方品种永川白麦子、万县白麦子和栽培品种西农979等抗穗发芽种质资源中存在两个新等位基因Vp-1Bb和Vp-1Bc。与野生型Vp-1Ba相比,Vp-1Bb和Vp-1Bc分别在第三个内含子区域存在193 bp的插入和83 bp的缺失,且这些插入和缺失片段均与穗发芽抗性紧密相关[36]。此外,在Vp-1B位点还存在另一个等位基因Vp-1Bf。与野生型Vp-1Ba序列比对,Vp-1Bf在第三个内含子区域存在一个193 bp的插入和109 bp的缺失。表型分析发现,携带Vp-1Bf等位基因材料的平均发芽指数(20%)显著低于野生型(58%)[37]。目前,Vp-1A基因存在6个等位基因,但只有Vp-1Ab(19%)与穗发芽抗性相关[38]。
2.6 TaMKK3-A基因
在小麦4AL染色体上定位到一个控制种子休眠的主效QTL(phs1)[39]。Torada等[40]利用加拿大强休眠性品种Leader和日本弱休眠性品种Haruyokoi杂交,并与Haruyokoi回交构建BC3F2群体,将phs1定位在分子标记Xhbe03和Xbarc170之间的2.9 cM区段;之后又通过图位克隆获得候选基因TaMKK3-A,该基因全长 7 056 bp,包含11个外显子和10个内含子,编码523个氨基酸[16]。MKK3基因编码丝裂原活化蛋白激酶,参与ABA信号转导,而ABA是高等植物种子休眠的重要因素[16]。在TaMKK3-A基因起始密码子下游+665 bp处存在一个SNP (C/A),根据这个SNP开发了功能标记TaMKK3-A-caps,可用于检测穗发芽抗性[16,19]。
3 小麦穗发芽主要抗源
穗发芽抗源是小麦穗发芽抗性育种的基础,也为抗穗发芽基因挖掘提供了宝贵的遗传材料。结合国内外研究结果,发现一些抗穗发芽较好的小麦资源(表2),具体如下:
丰产3号:原西北农学院赵洪璋先生利用丹麦1号与西农6028杂交培育的一个抗穗发芽白粒小麦品种,在连续两年的穗发芽抗性鉴定试验中,发现其平均发芽指数为3.9%[41]。宋 菲[42]利用丰产3号/84-1155 RIL群体,在1B、3A和6B染色体上定位到3个QTLs,其中,位于3A染色体上的QTL与上文介绍的TaMFT-3A(TaPHS1)基因位置相同。笔者在前期的研究中也证实,丰产3号含有TaMFT-3A抗性基因。
百农3217:原河南省百泉农专黄光正先生利用多亲本复交培育成的高产多抗白粒小麦品种。经过多年多点的穗发芽抗性鉴定试验,发现发芽指数均低于8%,并且含有Vp-Bc抗性等位基因[43]。在笔者前期的研究中,发现百农3217含有和丰产3号相同的TaMFT-3A抗性等位基因。丰产3号和百农3217是目前国内较好的抗穗发芽白粒小麦品种,且农艺性状相对较好,可以作为抗穗发芽育种亲本。
万县白麦子:四川万县地方品种,Zhang等[44]发现其平均发芽指数低于8%,并利用万县白麦子/京411和万县白麦子/中优9507两个RIL群体,分别在3AS和3BL染色体上定位到一个主效QTLs,其中,3AS染色体上的QTL可解释25.6%~48.3%的表型变异,与TaMFT-3A(TaPHS1)基因位置相同;3BL染色体上的位点可解释23.5%~37.8%的表型变异,与TaVp-1B基因位置相同。
秃头麦:四川省地方品种,Chen等[45]在5年的穗发芽抗性鉴定试验发现,籽粒发芽指数均低于10%,并利用秃头麦/泗阳936 RIL群体,在4AL染色体上定位到一个主效QTL,可解释 28.3%~30.6%的表型变异,与TaMKK3-A基因共分离。此外,秃头麦也含有TaMFT-3A抗性等位基因[46]。
Danby:美国堪萨斯州立大学通过Trego与KS84063-9-39-3-8w杂交选育的白粒小麦品种,具有较强的穗发芽抗性。Shao等[13]利用Danby/Tiger DH群体在3AS、3BS和5AL染色体上定位到3个QTLs,其中位于3AS染色体上的主效QTL(TaMFT-3A)可解释21.6%~41.0%的表型变异。
Rio Blanco:美国堪萨斯州Agripro生物科学公司培育的白粒冬小麦品种,是抗穗发芽育种的优异亲本[47]。Liu等[48]在3AS染色体定位到一个抗穗发芽的主效QTL(QPhs.pseru-3AS),可解释18.5%~41.0%的表型变异。之后,并进一步对这个主效QTL进行精细定位,并成功克隆了抗穗发芽基因TaPHS1(TaMFT-3A)[15,49]。
Zenkoujikomugi:日本红粒小麦品种,具有极高的种子休眠性和穗发芽抗性。Mori等[50]在3年的穗发芽抗性鉴定试验中,发现其发芽指数均在2%左右,并在3AS染色体上定位到一个与休眠相关的主效QTL,可解释11.6%~44.8%的表型变异。Nakamura等[14]利用Zenkoujikomugi/中国春染色体片段代换系,成功克隆该QTL候选基因TaMFT-3A。

表2 不同小麦穗发芽抗源携带的抗性基因/位点Table 2 Genes/QTLs associated with the pre- harvest sprouting resistance detected in different wheat resistance sources
4 小麦抗穗发芽分子育种研究进展
小麦穗发芽抗性是一个由多基因控制的复杂数量性状,单纯的表型筛选易受环境条件的影响,选择效率较低。小麦穗发芽抗性基因/QTLs的发掘和相关功能标记/连锁标记的开发,对开展穗发芽抗性分子标记辅助育种和提高育种效率奠定了基础。
4.1 分子标记辅助选择育种
在目前已定位和克隆的穗发芽抗性QTLs和基因中,位于3AS染色体上的TaMFT-3A(TaPHS1)基因和4AL染色体上的TaMKK3-A应用效果最好。利用分子标记辅助回交策略,Lin等[46]将含有TaMFT-3A和TaMKK3-A两个抗性基因的白粒小麦品种Tutoumai A和AUS1408分别与美国穗发芽敏感小麦品种NW97S186杂交并连续回交,结果发现,TaMFT-3A抗性等位基因在多个环境下均能够增强穗发芽抗性;TaMKK3-A抗性等位基因也能够增强穗发芽抗性,但易受环境条件影响;两个抗性等位基因同时存在时,穗发芽抗性显著增强。日本红粒抗穗发芽小麦品种Zenkoujikomugi含有TaMFT-3A、R和QPhs-5AL三个抗性基因[14,50],Kottearachchi等[52]利用Zenkoujikomugi的抗性基因改良日本白粒小麦品种Spica的穗发芽抗性,结果发现,后代红粒小麦和白粒小麦中含有TaMFT-3ARILs的穗发芽抗性均明显好于不含有TaMFT-3A的RILs,说明TaMFT-3A能够有效地增强穗发芽抗性。通过回交结合分子标记辅助选择,Kumar等[53]将小麦品种SPR8198(抗性等级为1)的抗性QTL(QPhs.ccsu-3A.1)导入到印度穗发芽敏感小麦品种HD2329(抗性等级为9)中,结果发现,11个BC3F3后代抗性等级为2~6级,其中,有7个抗性等级为2~4级,抗穗发芽能力明显提高。为了证实4AL染色体上主效QTL(QPhs-4AL)的有效性,Singh等[54]利用加拿大60个小麦品种和高代系对QPhs-4AL紧密连锁的分子标记DuPw004、Barc170和Wmc650进行验证,并证实这些标记可用于穗发芽抗性分子标记辅助选择。
国内穗发芽抗性分子标记辅助选择育种进展相对缓慢,因此,亟待加强。Xiao等[55]通过利用携带QPhs.pseru-3AS位点的地方品种万县白麦子作为供体,与不抗穗发芽的京411进行杂交,利用4个连锁标记Xbarc57、Xbarc294、Xbarc310和Xbarc321对后代穗发芽抗性进行选择,培育出小麦新品种中麦911,两年的平均发芽指数为 13.2%,穗发芽抗性比亲本京411明显增强。白粒小麦品种安农0711,在2014年通过安徽省审定,该品种聚合了亲本百农64和矮早781 3BL和3AS染色体上抗穗发芽基因的优异等位变异,因而具有较低的发芽指数、种子发芽率以及整穗发芽率[56]。
4.2 转基因育种
尽管转基因技术在抗除草剂、抗虫等方面取得巨大成功,并有较多报道,但在小麦穗发芽抗性改良方面报道相对较少。Vp-1基因编码的转录因子参与ABA信号转导途径,控制种子休眠,与小麦和其他谷物的穗发芽抗性有关[34]。Huang等[57]将玉米的Vp-1基因包括启动子和编码区转入到小麦品种郑麦9023中,转基因T3、T4和T5代种子的发芽指数与野生型郑麦9023相比,分别降低了79%、80%和82%,表明玉米的Vp-1基因能够有效提高籽粒的休眠性,增强小麦抗穗发芽能力。DOG1是拟南芥中影响种子休眠的一个重要基因[58]。Ashikawa等[59]将小麦的TaDOG1L4和大麦的HvDOG1L1两个DOG1-like基因通过农杆菌介导法转入到小麦品种Fielder中,过表达转基因植株能够明显提高籽粒的休眠性,且TaDOG1L4对小麦穗发芽的抗性改良更 有效。
4.3 抗穗发芽基因编辑育种
CRISPR(clustered regularly interspaced short palindromic repeats)基因编辑技术在过去短短几年内取得飞速发展,已经成为植物功能基因研究和作物遗传改良的重要工具[60-61]。Qsd1基因编码丙氨酸氨基转移酶,是控制大麦籽粒休眠的一个重要基因[62]。2019年,Abe等[63]利用CRISP/Cas9技术对Qsd1小麦同源基因进行编辑,试图通过延长籽粒的休眠性来提高小麦的抗穗发芽能力,结果发现,被编辑后的小麦比野生型休眠期明显延长,穗发芽抗性得到显著提高。除了小麦,该技术同样应用于其他禾谷类作物穗发芽抗性改良,如Jung等[64]通过CRISPR/Cas9技术敲除OsVp1基因后,水稻籽粒的休眠性明显降低。miR156是水稻一个重要产量调节因子,Miao等[65]发现通过基因编辑MIR156家族成员MIR156a、MIR156b、MIR156c、MIR156k和MIR156l,能够抑制GA通路,进而增强籽粒的休眠性,而且编辑这些基因对水稻株型结构和籽粒大小没有不利影响。
5 小麦穗发芽研究的重点和抗性育种思路
虽然小麦穗发芽抗性研究取得了较大进展,且分子标记辅助选择技术在许多国家已被广泛用于小麦穗发芽抗性育种,但今后仍需加强以下几方面的工作:(1)加强穗发芽抗源的收集、鉴定和利用。目前公认的小麦穗发芽抗源十分有限,应加强对地方品种和近缘种的发掘和鉴定工作,同时,对生产上推广品种的穗发芽抗性鉴定工作也需要加强,由于这类品种农艺性状通常较好,一旦鉴定出穗发芽抗性好的品种,可以直接作为抗源用于穗发芽抗性改良。(2)加强穗发芽抗性基因的发掘。已经发现的小麦穗发芽抗性相关的QTLs主要集中在3A和4A染色体上。整体而言,主效位点偏少。因此,在加强种质资源穗发芽抗性评价的基础上,应当积极开展穗发芽抗性基因的发掘。小麦基因组测序工作已经取得突破性进展,为穗发芽抗性基因研究提供了便利条件。(3)加强分子标记辅助及标记聚合育种工作。尽管已有利用基因编辑和转基因技术获得抗穗发芽材料的成功实例,但这类材料很难获得安全证书,只能用于基础研究,难于直接用于生产。分子标记辅助选择育种将是主要的分子育种手段之一。目前国外在利用TaMFT-3A和TaMKK3-A基因改良穗发芽抗性方面取得了一定进展,但是国内利用分子标记辅助选择育成抗穗发芽并大面积推广的品种鲜有报道,聚合不同抗穗发芽基因的分子聚合育种的实例更少。(4)尽快完善穗发芽抗性鉴定评价方法和评价标准。穗发芽籽粒鉴定法费工费时,不适合育种上大量的育种材料筛选,而且该方法是在脱粒以后对穗发芽特性进行鉴定,没有考虑到穗部结构对穗发芽的影响,其鉴定结果与穗部结构的相关性还有待进一步研究;建议利用该方法计算穗发芽指数时,增加相对发芽指数的指标(即待测材料发芽指数与对照品种发芽指数的比值),以便更有效地进行不同年份、不同条件下材料间穗发芽抗性水平的比较。小麦抗穗发芽性鉴定方法(NY/T 1739-2009)在标准操作程序上还有待完善。笔者发现按照该鉴定方法进行穗发芽鉴定时,如果把穗子浸泡4 h,往往造成具有不同穗发芽抗性的白粒小麦品种均表现出较高的发芽指数,难于把具有不同穗发芽抗性的白粒小麦品种区分开来。(5)扩大回交育种群体。由于目前鉴定出的小麦抗穗发芽资源非常有限,抗穗发芽育种经常使用一些农艺性状和适应性较差、但穗发芽抗性较好的材料做亲本。为了快速提高小麦品种对穗发芽的抗性,同时又保持改良后较好的农艺性状和适应性,回交育种结合分子标记辅助选择是抗穗发芽育种最有效的方法之一。结合本课题组在黄淮麦区小麦抗赤霉病分子育种方面取得的进展[66],目前我们正尝试利用相同的研究思路来改良该麦区的小麦穗发芽抗性。在前期工作中,本课题组培育了骨干亲本的矮败小麦近等基因系,比如矮败周麦16、矮败郑麦9023、矮败济麦22、矮败周麦18等,利用这些骨干亲本的矮败近等基因系作为受体亲本,抗穗发芽种质资源作为穗发芽抗性基因的供体亲本,进行杂交和回交,通过分子标记对抗穗发芽基因进行跟踪选择。由于利用矮败小麦做杂交时不需要人工去雄,可以创建大规模的回交育种群体,在较低的回交世代里就有可能获得主要农艺性状基本恢复到轮回亲本类型、同时携带抗穗发芽基因的后代材料,从而加快抗穗发芽育种速度。(6)加强分子标记辅助选择育种技术和表型鉴定技术相结合。在低回交世代,通常通过分子标记跟踪目标基因,高世代主要采取整穗发芽鉴定法对育种材料进行表型鉴定。根据穗发芽鉴定结果,决定田间收获单株的取舍。
猜你喜欢
当代水产(2022年7期)2022-09-20
中国饲料(2021年17期)2021-11-02
智慧健康(2021年17期)2021-07-30
阅读(低年级)(2021年4期)2021-06-15
新课程·下旬(2018年9期)2018-11-14
小天使·五年级语数英综合(2016年4期)2016-04-21
青少年科技博览(中学版)(2015年10期)2015-01-11
发明与创新·中学生(2014年12期)2014-12-16
食品工业科技(2014年13期)2014-03-11
诗潮(2009年10期)2009-12-08
