
外源抗坏血酸对水稻抗铝生理指标的影响
2021-05-19 10:37周小华李昆志赵峥张小玲程霞冯庆
热带作物学报 2021年3期
周小华 李昆志 赵峥 张小玲 程霞 冯庆
摘 要:为探讨外源抗坏血酸(AsA)对水稻的抗铝性能的影响,以‘滇优35号(杂交稻,粳稻)为试验材料,采用溶液培养法研究外源AsA对铝胁迫下水稻根尖H2O2、内源AsA、GSH、可溶性蛋白、脯氨酸含量及抗氧化酶活性等生理生化指标的影响。结果显示,与对照相比,铝胁迫导致水稻根尖可溶性蛋白和内源AsA含量减少,而MDA和H2O2含量,SOD、POD、APX和CAT活性,GSH及脯氨酸含量增加;外源AsA可增强铝胁迫水稻根尖的脯氨酸及可溶性蛋白等含量,内源AsA、GSH含量及SOD、POD、APX、CAT活性,而降低MDA和H2O2含量。表明外源AsA通过调控抗氧化酶活性,提高渗透调节物质含量和抗氧化物质含量来降低H2O2的积累,从而缓解铝胁迫下水稻质膜氧化程度,增强水稻的抗铝性能。
关键词:水稻;铝胁迫;抗坏血酸;抗氧化酶;渗透调节物质
中图分类号:S511 文献标识码:A
Effects of Exogenous Ascorbic Acid on Physiological Indexes in Rice Under Aluminum Stress
ZHOU Xiaohua1, LI Kunzhi2, ZHAO Zheng1, ZHANG Xiaoling3, CHENG Xia4, FENG Qing1
1. Kunming Cooperative Research Center for Pollution Prevention of Dianchi (Lake), Kunming University, Kunming, Yunnan 650214, China; 2. Faculty of Life Science and Biotechnology, Kunming University of Science and Technology, Kunming, Yunnan 650500, China; 3. Teaching Affairs Office, Kunming University, Kunming, Yunnan 650214, China; 4. Faculty of Agronomy and Life Sciences, Kunming University, Kunming, Yunnan 650214, China
Abstract: In order to investigate the effect of exogenous ascorbic acid (AsA) on aluminum resistance of rice, the ‘Dian Superior 35 (Hybrid, Japonica) was used as the hydroponical test material to study the influence of the physiological and biochemical indicators such as the contents of H2O2, endogenous AsA, GSH, the soluble protein, proline and the activity of the antioxidant enzyme, etc. in the rice root tips. The results showed that compared to the control, the contents of the soluble protein and endogenous AsA in the root were decreased, while the activities of SOD, POD, APX and CAT, the contents of MDA, H2O2, GSH and proline were significantly increased under aluminum stress, respectively, the exogenous AsA migth improve the contents of proline, the soluble protein, endogenous AsA and GSH, the activities of SOD, POD, APX and CAT, respectively, and decreased the contents of MDA and H2O2. The above results showed that exogenous AsA might alleviate the degree of the plasma membrane oxidation and promote aluminum resistance of rice by enhancing the activities of the antioxidant enzyme, the contents of osmoregulation substances and antioxidant substance which might reduce the accumulation of H2O2.
Keywords: rice; aluminum stress; ascorbic acid; antioxidant enzyme; osmoregulation substances
DOI: 10.3969/j.issn.1000-2561.2021.03.023
目前全球约50%的可耕土壤为酸性土壤,我国约有21%的可耕土壤为酸性土壤[1-2],当土壤pH<5.5时,土壤中难溶性铝形态物质就会因土壤的酸性而溶解釋放出Al3+,土壤中微摩尔级Al3+就会影响植物的生长,抑制植物吸收氮、磷等营养素[3],铝胁迫可导致玉米[4]、水稻[5]、甘薯[6]等作物减产。我国约为30%的水稻生长在酸性土壤上,铝胁迫已成为限制水稻生长的主要影响因子之一。
通过外源物质调节铝胁迫下水稻的生长已有相关报道,外源脱落酸[7]、NO[8]等能有效缓解铝胁迫诱导的氧化损伤。抗坏血酸是植物的重要非酶抗氧化剂,也是植物体内的一种多功能代谢物质,可以直接或者间接地清理体内产生的ROS,维持植物体内的氧化还原平衡,增强植物抗逆性[9],其在植物逆境条件下的作用引起了学者的广泛关注。已有研究表明,外源AsA可提高Na2SO4胁迫下燕麦幼苗根长及活力指数[10];能显著增强高温胁迫下水稻[11]、铅胁迫下秋葵[12]及干旱胁迫下玉米[13]的抗氧化酶活性,以缓解环境胁迫对植株的伤害,提高植物的抗逆性。然而,关于外源AsA对水稻抗铝的生理指标的影响研究较少,本研究以4叶龄水稻幼苗为试验材料,探讨外源AsA对铝胁迫水稻幼苗根尖的H2O2含量与脯氨酸等渗透调节物质含量、抗氧化酶活性、AsA和GSH抗氧化物质含量等指标的影响,阐明外源AsA对铝胁迫水稻的生理生化反应机制,为科学评价外源AsA对非生物胁迫下农作物生长及其应用提供理论依据。
1 材料与方法
1.1 材料
供试水稻(Oryza sativa L.)为‘滇优5号,由云南农业大学水稻研究所提供。种子经消毒、浸泡、催芽,将露白的种子转移至营养液于25 ℃、光照[1200 μmol/(m2·s)] 12 h/d的温室中漂浮培养和处理。
试剂:AlCl3,AR;抗坏血酸,AR。
水稻营养液(IRRI,国际水稻研究所):K2SO40.35、KH2PO40.3、NH4NO32.86、CaCl21.0、H3BO32.0×10–2、MnCl2·4H2O 9.0×10–3、CuSO4·5H2O 3.2×10–4、MgSO4·7H2O 1.0、(NH4)6Mo7O24·4H2O 0.39×10–4、Na2SiO3·9H2O 5.0×10–4、Na2EDTA-Fe (Ⅱ) 2.0×10–2、ZnSO4·7H2O 7.7×10–4,单位:mmol/L。
1.2 方法
1.2.1 材料的处理 用含1/4浓度pH 4.5的水稻营养液(IRRI)培养至1叶龄幼苗,放至pH4.5水稻全营养液培养,每2 d更换1次营养液,待水稻长至4叶龄进行铝胁迫(0、50 μmol/L)处理[14]。AsA最佳浓度实验是在含50 μmol/L AlCl3的pH 4.5水稻全营养液中添加不同浓度(0、0.5、1.0、2.0、4.0、8.0 mmol/L)AsA处理24 h,分析根尖材料的H2O2含量,根据H2O2含量确定AsA最佳处理浓度;采用无铝无AsA(–Al–AsA)、有铝无AsA(+Al–AsA)、无铝有AsA(–Al+AsA)和有铝有AsA(+Al+AsA)处理24 h,以无铝无AsA(–Al–AsA)组为对照,铝以AlCl3形式供给,每组3个重复,收集根尖材料备用。
1.2.2 测定指标及方法 参照王学奎等[15]的方法测定MDA、可溶性蛋白质、脯氨酸、可溶性糖含量,根系活力,APX、SOD、POD、CAT活性;参照Nakano等[16]的方法测定APX活性;参照Gay等[17]的方法测定H2O2含量;参照贾新平等[18]的方法測定质膜透性;参照朱为民等[19]的方法测定AsA和GSH含量。
1.2.2 测定指标及方法 参照王学奎等[15]的方法测定MDA、可溶性蛋白质、脯氨酸、可溶性糖含量,根系活力,APX、SOD、POD、CAT活性;参照Nakano等[16]的方法测定APX活性;参照Gay等[17]的方法测定H2O2含量;参照贾新平等[18]的方法测定质膜透性;参照朱为民等[19]的方法测定AsA和GSH含量。
1.3 数据处理
用Excel 2010软件处理数据,Duncans法分析数据差异性。
2 结果与分析
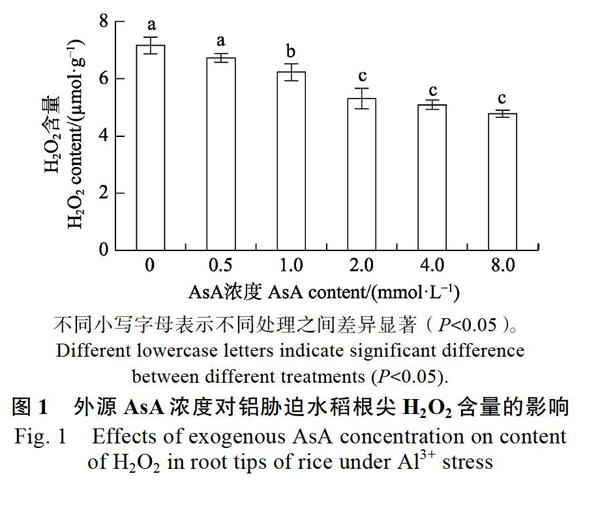
2.1 不同AsA浓度对铝胁迫水稻根尖H2O2含量的影响
不同AsA浓度处理铝胁迫水稻24 h,与铝胁迫组相比,根尖H2O2含量均有不同程度的减少(图1);≥1.0 mmol/L AsA处理组的H2O2含量显著低于无AsA铝胁迫组,2.0 mmol/L AsA处理组的H2O2含量显著低于1.0 mmol/L AsA处理组;而大于2.0 mmol/L AsA处理组的H2O2含量与2.0 mmol/L AsA处理组无显著差异,表明2.0 mmol/L AsA能较好地降低根尖H2O2含量。因此,均采用2.0 mmol/L AsA进行下一步实验。
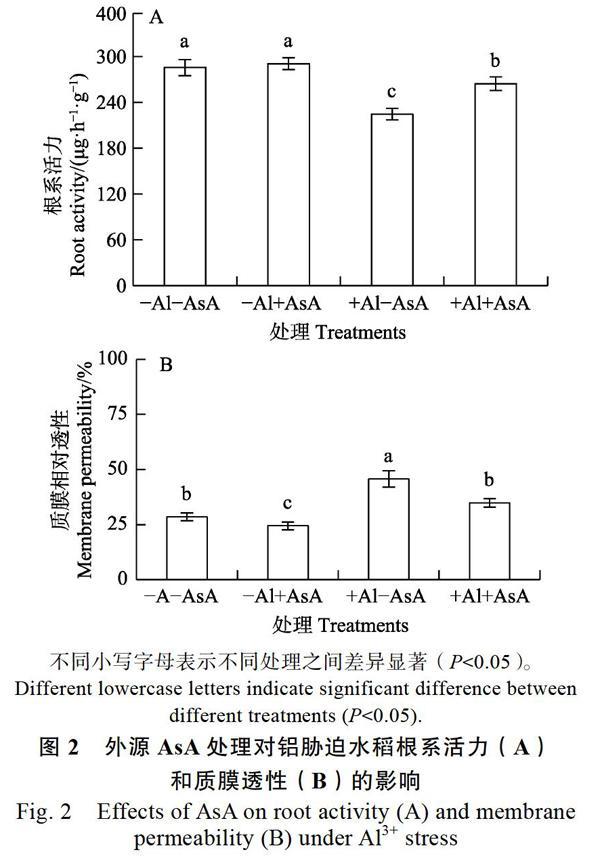
2.2 AsA对铝胁迫水稻根系活力及质膜相对透性的影响
根系活力反映植物根系的代谢水平和生长情况,直接影响植物营养物质的吸收与生长。有铝无AsA(+Al–AsA)处理时根尖根系活力(图2A)和质膜相对透性(图2B)含量分别为对照(–Al–AsA)的0.79倍和1.60倍,无铝有AsA(–Al+AsA)处理组根系活力及质膜透性与对照差异不大;有铝有AsA(+Al+AsA)处理组分别为对照的0.93倍和1.22倍,为铝胁迫(+Al–AsA)处理组的1.18倍和0.76倍,根系活力恢复到接近对照水平。表明外源AsA可以显著提高铝胁迫水稻根系活力并降低根尖质膜透性,有助于维持细胞质膜结构的完整。
ability (B) under Al3+ stress
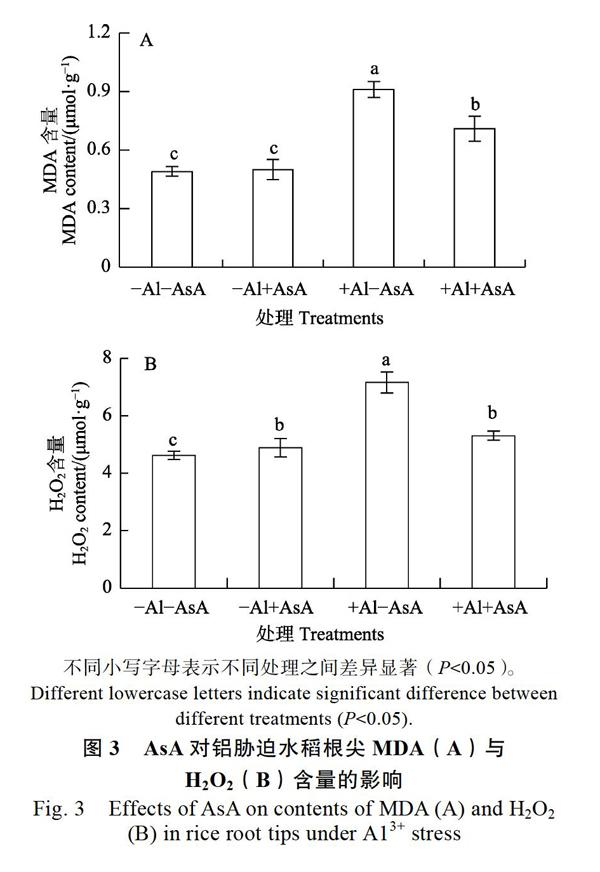
2.3 AsA对铝胁迫水稻根尖MDA及H2O2含量的影响
铝胁迫(+Al–AsA)处理组根尖MDA(图3A)和H2O2(图3B)含量分别为对照(–Al–AsA)的1.86倍和1.55倍,无铝有AsA(–Al+AsA)处理组根尖MDA和H2O2含量为对照的1.02倍和1.05倍;有铝有AsA(+Al+AsA)处理组根尖MDA和H2O2含量降低到对照(–Al–AsA)的1.50倍和1.15倍,为铝胁迫(+Al–AsA)处理组的0.78倍和0.74倍。表明铝胁迫使植株体内MDA和H2O2含量升高,外源AsA可以有效地降低H2O2在水稻根部的积累,膜脂过氧化程度明显下降。
2.4 AsA对铝胁迫水稻根尖可溶性蛋白、可溶性糖及脯氨酸含量的影响
铝胁迫(+Al–AsA)处理组水稻根尖可溶性蛋白质含量(图4A)显著降低,为对照(–Al–AsA)的0.65倍,可溶性糖含量(图4B)和游离脯氨酸含量(图4C)显著增加,分别为对照(–Al–AsA)的1.61倍和1.65倍;无铝有AsA(–Al+AsA)处理组的可溶性糖、游离脯氨酸和可溶性蛋白质含量与对照(–Al–AsA)无显著差异;有铝有AsA(+Al+AsA)处理组根尖可溶性蛋白质、可溶性糖及脯氨酸含量分别为对照(–Al–AsA)的0.75倍、1.87倍和2.29倍,为铝胁迫(+Al–AsA)组的1.15倍、1.16倍和1.39倍。表明外源AsA可提高根尖可溶性蛋白质、可溶性糖及脯氨酸等渗透物质的含量,有利于维持铝胁迫水稻的质膜渗透平衡。
2.5 AsA对铝胁迫水稻根尖SOD、POD、CAT和APX活性的影响
铝胁迫(+Al–AsA)处理组水稻根尖SOD(图5A)、POD(图5B)、CAT(图5C)及APX(图5D)活性分别是对照(–Al–AsA)的1.77倍、1.80倍、1.76倍和1.73倍;无铝有AsA(–Al+AsA)处理组与对照(–Al–AsA)均无显著差异;有铝有AsA(+Al+AsA)处理组根尖的SOD、POD、CAT及APX活性分别为对照(–Al–AsA)的2.34倍、2.20倍、2.28倍和2.13倍,为铝胁迫(+Al–AsA)处理组的1.26倍、1.22倍、1.29倍和1.23倍。表明铝胁迫能增加根尖抗氧化酶的活性,外源AsA能更加突出这种增强效果,增强水稻清除ROS的能力。
2.6 AsA对铝胁迫水稻根尖内源AsA与GSH含量的影响
AsA与GSH是植物体内非常重要的抗氧化物质,植物抵抗非生物胁迫的能力与其含量呈正相关关系,在植物增强抗环境胁迫过程中发挥重要作用[20]。铝胁迫(+Al–AsA)处理组水稻根尖内源AsA含量(图6A)及GSH含量(图6B)分别是对照(–Al–AsA)的0.93倍和1.36倍;无铝有AsA(–Al+AsA)处理组是对照(–Al–AsA)的1.43倍和1.19倍;有铝有AsA(+Al+AsA)处理组的AsA及GSH含量分别为对照(–Al–AsA)的1.32倍和1.73倍,为铝胁迫(+Al–AsA)处理组的1.42倍和1.27倍。表明铝胁迫降低了内源AsA含量,但增加了GSH含量,外源AsA可增加铝胁迫水稻根尖的内源AsA和GSH含量。
2.7 不同生理生化指标之间的相关性分析
采用SPSS21.0软件对外源AsA处理铝胁迫水稻根尖的H2O2含量(A)、MDA含量(B)、质膜透性相对值(C)、可溶性蛋白含量(D)、可溶性总糖含量(E)、脯氨酸含量(F)、内源AsA含量(G)、GSH含量(H)、根系活力(I)、SOD活性(J)、POD活性(K)、CAT活性(L)和APX活性(M)进行双变量相关分析。结果表明,铝胁迫下水稻根尖MDA含量变化与CAT活性变化、内源AsA含量变化与APX活性变化间呈显著的正相关,Pearson相关系数r分别为0.987和0.964(表1),表明外源AsA处理铝胁迫水稻根尖MDA含量主要由CAT与APX活性来调控。
3 讨论
外源AsA能增加铝胁迫水稻渗透调节物质的分泌量。脯氨酸和可溶性蛋白等能维持逆境胁迫的细胞渗透平衡和激发抗氧化酶活性,植物积累越多渗透调节物质,就有较强的逆境适应能力[21]。本研究中,铝胁迫导致根尖脯氨酸含量增加、可溶性蛋白含量下降,铝胁迫水稻根尖正常的蛋白质合成受到抑制,蛋白质分解速度加快,导致其含量降低,通过增加脯氨酸含量保护质膜结构完整性和提高酶的活性,提高水稻的抗铝性能,这与通过提高脯氨酸和蛋白质含量来增强油菜幼苗抗铝性的研究结果相似[22]。外源AsA可使铝胁迫水稻可溶性蛋白含量增加,其促进了蛋白质的水合作用,分解速度下降所致,水稻脯氨酸含量增加更有利于保持质膜的渗透平衡,这与华智锐等[23]的AsA可通过调节渗透调节物质分泌量来增强小麦抗盐性的结论一致。说明外源AsA进一步增强了脯氨酸和可溶性蛋白质分泌量,缓解质膜氧化損伤,有效地维持铝胁迫水稻的细胞渗透平衡,增强水稻的抗铝性能。
外源AsA可降低铝胁迫水稻ROS的积累。MDA、H2O2常用于反映细胞质膜氧化程度的生理指标[24]。已有研究表明,铝胁迫能加剧植物质膜过氧化程度,使细胞膜结构受损,从而导致植物体内积累大量的H2O2等ROS[25-26]。本研究表明,铝胁迫导致水稻根尖积累较多的MDA和H2O2,质膜过氧化程度加剧,这与铝胁迫诱导水稻、玉米ROS的大量积累的研究结果相似[27-29]。外源AsA可减少铝胁迫水稻根尖MDA及H2O2的积累,AsA直接或间接参与了清除ROS的过程,在一定程度上降低ROS的积累,缓解铝毒对质膜的过氧化伤害,从而有利于维持水稻幼苗正常的生理生化反应,这与外源AsA减少盐胁迫小麦体内的ROS含量的结果一致[30]。表明外源AsA可清除H2O2等ROS,降低铝对水稻根系的过氧化水平,提高水稻的耐铝能力。
外源AsA可提高铝胁迫水稻的根系活力和质膜结构的完整性。在逆境条件下,细胞质膜系统结构因质膜过氧化而受损,导致质膜透性增大及根系活力下降[31-32],本研究中,铝胁迫导致水稻根尖质膜相对透性增加,根系活力下降,铝胁迫导致水稻根尖H2O2大量积累,加剧了细胞膜脂过氧化程度,破坏了质膜结构的完整性,从而造成细胞膜透性增大和根尖活力下降,这与罗文姬等[30]和刘拥海等[33]研究的铝胁迫尾桉、水稻根系质膜相对透性增加,与黄玉婷等[34]研究的铝胁迫草本植物根系活力降低的变化趋势相似。外源AsA可降低铝胁迫水稻质膜相对透性并提高根系活力,AsA参与消除了H2O2等ROS过程,降低了水稻根系积累的H2O2量,缓解了铝胁迫诱导的质膜过氧化程度,维持了质膜结构的完整性,从而降低质膜相对透性并提高了根系活力。
外源AsA可提高铝胁迫水稻的抗氧化物质含量和抗氧化酶活性。在逆境条件下,植物为了提高逆境胁迫的适应能力,会启动体内的抗氧化酶系统或提高抗氧化物质含量以消除ROS,从而增强抵御逆境胁迫诱导的ROS对植物的伤害程度[35-36],本研究结果显示,铝胁迫导致水稻根尖SOD、POD、CAT及APX活性增加,表明铝胁迫在一定程度上诱导了水稻根尖抗氧化酶基因的表达,提高了抗氧化酶活性,从而增加消除ROS的能力,研究结果与郑阳霞等[37]报道铝胁迫豆瓣菜抗氧化酶活性增加的趋势一致,铝胁迫导致水稻内源AsA含量减少,GSH含量增加,说明在铝胁迫初期AsA直接参与清除ROS或者其作为APX的电子供体而降低了其含量,在胁迫后期主要通过抗氧化酶和GSH来清除ROS,这与Sharma等[38]研究铝胁迫降低水稻AsA含量的结果一致;外源AsA进一步提高水稻根尖抗氧化酶活性,其通过减少降解生物大分子的速率或上调抗氧化酶相关基因的表达来提高抗氧化酶活性,研究结果与外源AsA提高玉米的抗氧化酶活性以增强玉米的抗旱能力的结果相似[39];并可上调内源AsA和GSH的生物合成,弥补铝胁迫导致水稻内源AsA的不足,维持较稳定的还原环境和还原库力水平,增强清除H2O2等ROS的能力,减少根尖ROS的积累,维持质膜的结构完整性和化学成分的稳定性,进而提高水稻的抗氧化能力,表明外源AsA通过调节抗氧化酶活性和抗氧化物质含量来提高水稻抗铝能力。
MDA是逆境胁迫植物质膜过氧化产物之一,其含量可表征ROS的积累程度,引发细胞质膜严重损伤,H2O2可在CAT作用下分解成无毒的H2O和O2,减少氧化损伤对植物的伤害;外源AsA通过上调铝胁迫水稻内源AsA的生物合成,补充铝胁迫导致内源AsA的不足,持续提高APX的电子供给能力,从而增强APX的活性,增强抗氧化损伤能力[38]。双重相关性分析结果表明,MDA含量变化与CAT活性变化、内源AsA含量变化与APX活性变化呈显著的正相关,表明外源AsA主要通过诱导水稻体内的CAT、APX活性调控水稻铝胁迫的应答机制,以缓解铝胁迫诱导的ROS对水稻的伤害程度。
综上所述,外源AsA显著提高了抗氧化酶活性及内源AsA与GSH含量,减少ROS的积累,降低铝诱发的膜脂过氧化水平,并提高了可溶性蛋白含量、可溶性糖含量及脯氨酸含量,将细胞渗透平衡和细胞膜结构维持在较稳定水平,从而增强了水稻的抗氧化能力。外源AsA主要通過提高CAT、APX活性及内源AsA含量提高铝胁迫水稻清除ROS的能力,从而有效地缓解了铝胁迫对水稻根系造成的氧化损伤,增强水稻的抗铝性能。
参考文献
[1] 任继鹏, 张 逸, 钱 诚, 等. 南方酸性森林土壤中铝的形态分布与活化机理[J]. 环境化学, 2011, 30(6): 1131-1135.
[2] 黄春琼, 刘国道, 陈 振. 耐铝性不同的狗牙根在酸性土壤上的生长差异[J]. 热带作物学报, 2019, 40(11): 2112-2118.
[3] Fan Y, OuyangY Y, Pana Y L, et al. Effect of aluminum stress on the absorption and transportation of aluminum and macronutrients in roots and leaves of Aleurites montana[J]. Forest Ecol Manage, 2020, 458: 117813.
[4] Welcker C, Thé C, Andréau B, et al. Heterosis and com-bining ability for maize adaptation to tropical acid soils[J]. Crop Sci, 2005, 45(6): 2405-2413.
[5] Kang D J, Seo Y J, Futakuchi K, et al. Effect of aluminum toxicity on flowering time and grain yield on rice genotypes differing in Al-tolerance[J]. Journal of Crop Science and Biotechnology, 2011, 14(4): 305-309.
[6] 李清华, 刘 庆, 李 欢, 等. 施钙对甘薯铝胁迫的缓解效应[J]. 中国土壤与肥料, 2016(4): 124-127, 132.
[7] 李 婧, 刘星星, 吴玉环, 等. 外源脱落酸对铝胁迫下栝楼叶绿素荧光及生理活性的影响[J]. 水土保持学报, 2017, 31(2): 293-300.
[8] 吴 坤, 邢承华, 饶玉春, 等. 外源NO对铝毒下水稻根系生长和抗氧化系统的影响[J]. 西北植物学报, 2014, 34(3): 536-542.
[9] Venkatesh J, Park S W. Role of L-ascorbate in alleviating abiotic stresses in crop plants[J]. Bot Stud, 2014, 55(1): 38.
[10] 董秋丽, 夏方山, 丁荷星, 等. 外源抗坏血酸引发促进Na2SO4胁迫下燕麦幼苗的生长[J]. 草业科学, 2018, 35(3): 558-565.
[11] Shah F, Huang J, Cui K, et al. Impact of high-temperature stress on rice plant and its traits related to tolerance[J]. J Agric Sci, 2011, 149(5): 545-556.
[12] Hussain I, Siddique A, Ashraf M A, et al. Does exogenous application of ascorbic acid modulate growthphotosynthetic pigments and oxidative defense in okra (Abelmoschus escu-lentus L.) under lead stress[J]. Acta Physiol Plant, 2017, 39(6): 144-156.
[13] Dolatabadian A, Modarres S M, Sharif M. Alleviation of water deficit stress effects by foliar application of ascorbic acid on zea mays[J]. Agron Crop Sci, 2009, 195(5): 347-355.
[14] 周小华, 徐慧妮, 谷照虎, 等. 铝胁迫下质膜H+-ATPase对水稻硝态氮吸收的影响[J]. 植物生理学报, 2016, 52(7): 1011-1018.
[15] 王学奎, 黄见良. 植物生理生化实验原理与技术[M]. 3版. 北京: 高等教育出版社, 2015.
[16] Nakano Y, Asada K. Hydrogen peroxide is scavenged by ascobate-specific peroxidase in spinach chloroplasts[J]. Plant Cell Physiol, 1981, 22(5): 867-880.
[17] Gay C A, Gebicki J M. Measurement of protein and lipid hydro peroxides in biological systems by the ferric-xylenol orange method[J]. Anal Bioch, 2003, 315(1): 29-35.
[18] 贾新平, 邓衍明, 孙晓波, 等. 盐胁迫对海滨雀稗生长和生理特性的影响[J]. 草业学报, 2015, 24(12): 204-212.
[19] 朱为民, 丁海东, 齐乃敏, 等. Cd2+胁迫对番茄幼苗抗坏血酸-谷胱甘肽循环代谢的影响[J]. 华北农学报, 2005, 20(3): 50-53.
[20] Cuypers A, Vangronsveld J, Clijsters H. The redox status of plant cells (AsA and GSH) is sensitive to zinc imposed oxidative stress in roots and primary leaves of phaseolus vulgaris[J]. Plant Physiol Bioch, 2001, 39(7-8): 657-666.
[21] 吴雪霞, 张圣美, 张爱冬, 等. 外源褪黑素对高温胁迫下茄子幼苗光合和生理特性的影响[J]. 植物生理学报, 2019, 55(1): 49-60.
[22] 韩德鹏, 刘星月, 王馨悦, 等. 铝胁迫对油菜根系形态和生理指标的影响[J]. 核农学报, 2019, 33(9): 1824-1832.
[23] 华智锐, 李小玲. 外源抗坏血酸对盐胁迫下商麦1619生长生理的影响[J]. 江西农業学报, 2019, 31(9): 1-8.
[24] Ding F, Liu B, Zhang S X. Exogenous melatonin ameliorates cold-induced damage in tomato plants[J]. Sci Hortic, 2017, 219(17): 264-271.
[25] Fan W, Xu J M, Wu P, et al. Alleviation by abscisic acid of Al toxicity in rice bean is not associated with citrate efflux but depends on ABI5-mediated signal transduction pathways[J]. J Integr Plant Biol, 2019, 61(2): 140-154.
[26] Muhammad N, Zvobgo G, Fu L, et al. Physiological mecha-nisms for antagonistic interaction of manganese and alumi-num in barley[J]. J Plant Nutr, 2019, 42(5): 466-476.
[27] Cardoso F B, Cambraia J, Oliveira J A de, et al. Alumi-num-induced citric acid secretion is not the sole mechanism of Al-resistance in maize[J]. Acta Physiol Plant, 2016, 38(12): 279-287.
[28] Zhou X H, Gu Z H, Xu H N, et al. The effects of exogenous ascorbic acid on the mechanism of physiological and biochemical responses to nitrate uptake in two rice cultivars (Oryzasativa L) under aluminum stress[J]. J Plant Growth Regul, 2016, 35(4): 1013-1024.
[29] 吳丽芳, 魏晓梅, 张丽芳, 等. 云南2种野生苜蓿资源对酸铝胁迫的响应分析[J]. 热带作物学报, 2020, 41(3): 482-488.
[30] 罗文姬, 罗必敬, 蒋 霞, 等. 铝胁迫下一氧化氮对巨尾桉生长及生理的影响[J]. 热带作物学报, 2020, 41(1): 77-82.
[31] 温泽林, 刘慧英, 周 艳, 等. 一氧化氮参与外源谷胱甘肽对盐胁迫下番茄幼苗抗氧化损伤的调控[J]. 植物生理学报, 2018, 54 (4): 607-617.
[32] 王旭明, 赵夏夏, 周鸿凯, 等. NaCl胁迫对不同耐盐性水稻某些生理特性和光合特性的影响[J]. 热带作物学报, 2019, 40(5): 882-890.
[33] 刘拥海, 俞 乐, 陈秀云, 等. 外源草酸对水稻幼苗铝毒害的缓解作用[J]. 湖南农业大学学报(自然科学版), 2008, 34(3): 281-283, 340.
[34] 黄玉婷, 吴 亚, 刘大林, 等. 铝胁迫对草本植物生理的影响机制[J]. 草业科学, 2018, 35(6): 1517-1527.
[35] Apel K, Hirt H. Reactive oxygen spedes: Metabolism, oxidative stress, and sigllal transduction[J]. Ann Review Plant Biol, 2004, 55: 373-399.
[36] Christelle M A, Yvan L, Danièle E. Dietary antioxidants and oxidative stress from a human and plant perspective: a review[J]. Curr Nutr Food Sci, 2010, 6(1): 2-12.
[37] 郑阳霞, 赵善梅, 向 前, 等. 铝胁迫对豆瓣菜生理特性及营养元素吸收的影响[J]. 甘肃农业大学学报, 2019, 54(4): 83-91.
[38] Sharma P, Dubey R. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum[J]. Plant Cell Reports, 2007, 26(11): 2027-2038.
[39] Dolatabadian A, Modarres Sanavy M, Sharif M. Alleviation of water deficit stress effects by foliar application of ascorbic acid on zea mays[J]. Agron Crop Sci, 2009, 195(5): 347-355.
责任编辑:沈德发
收稿日期 2020-05-05;修回日期 2020-05-22
基金项目 云南省高校联合基金项目(No. 2018FH001-027);昆明学院引进人才科研项目(No. YJL18009)。
作者简介 周小华(1973—),男,博士,副教授,研究方向:逆境植物营养与生理,E-mail:zxh200008@126.com。
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年23期)2019-08-16
农产品市场周刊(2017年35期)2017-10-25
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
河北渔业(2015年10期)2015-10-15
