
耐盐芦苇大片段BIBAC载体构建及水稻遗传转化
2021-05-19 10:09高鸿马启林吴孚桂刘慧芳聂佳俊
热带作物学报 2021年3期
高鸿 马启林 吴孚桂 刘慧芳 聂佳俊

摘 要:利用盐生植物的耐盐基因改良作物耐盐性是保障土壤盐渍化地区粮食生产和改良盐渍化土地的最经济最有效的途径。本研究以生长于西北盐盖上的一种耐盐野生芦苇为材料,制备了Mb级耐盐芦苇基因组DNA,酶切后经脉冲场凝胶电泳将其分离为不同大小的大片段DNA区段,并与酶切回收的BIBAC-S载体进行连接,成功构建了耐盐芦苇不同大小的大片段DNA-BIBAC载体,经根癌农杆菌EHA105介导,成功地转化了粳稻品种‘中花11成熟胚愈伤组织,获得了转基因水稻植株。研究表明,不同大小的大片段DNA-BIBAC载体在转化同一受体材料时,其所获得的愈伤转化率、植株转化率、转基因植株的阳性率等存在差异,说明插入片段的大小与转化效率之间存在关系。本研究为创制耐盐水稻新种质、克隆盐生植物的耐盐基因提供了理论和技术参考。
关键词:水稻;芦苇;耐盐性;BIBAC;大片段DNA转化
中图分类号:S511 文献标识码:A
Construction of Large Fragment DNA-BIBAC Vector of Salt Tolerant Reed and Its Genetic Transformation in Rice
GAO Hong, MA Qilin*, WU Fugui, LIU Huifang, NIE Jiajun
College of Tropical Crops, Hainan University, Haikou, Hainan 570228, China
Abstract: Using the salt-tolerant genes of halophytes to improve crop salt tolerance will be the most economical and effective way to ensure food production in soil salinized areas and to improve salinized land. In this study, a salt-tolerant wild Phragmites australis grown in the saline-alkali soil of Northwestern China was used as the materials to prepare Mb class genomic DNA, which was digested by restriction enzyme, and was separated into different size of DNA segments by pulsed field gel electrophoresis, then connected with the BIBAC-S vector, and successfully constructed the salt tolerant reed large fragment DNA-BIBAC vector. The large fragment DNA-BIBAC vector was successfully transformed into the callus of the mature embryo of ‘Zhonghua 11 mediated by Agrobacterium tumefaciens EHA105, and 23 transgenic positive rice plants were obtained. The results showed that there were differences in callus transformation rate, plant transformation rate and transgenic plant positive rate among different sizes of the DNA-BIBAC vector, indicating that there was a relationship between the size of inserted fragments and transformation efficiency. This study would provide a theoretical and technical reference for creating new germplasm of salt tolerant rice and cloning salt tolerant genes of halophytes.
Keywords: Oryza sativa L.; Phragmites australis; salt tolerance; BIBAC; genetic transformation of large DNA fragments
DOI: 10.3969/j.issn.1000-2561.2021.03.006
隨着全球气候变化和土地的不合理使用,世界上大约三分之一的耕地都不同程度地受到自然或次生盐渍化的影响[1-2],越来越严重的土壤盐渍化现象导致可耕地面积下降。土壤盐渍化是严重制约世界各地植物生长、造成作物减产的主要非生物胁迫因子之一[3-7],阻碍着农业生产的可持续发展,威胁着全球粮食安全[8-9]。为了维持作物正常生长、保障粮食安全,加强盐碱地资源的开发利用,改良和培育耐盐作物新品种已成为重要的课题和研究的热点。其中,选育和种植具有较高耐盐性的作物新品种,已经成为保障土壤盐渍化地区粮食生产和改良盐渍化土地的有效途径之一[10-11]。
水稻(Oryza sativa L.)是甜土植物,对盐胁迫敏感。从现有水稻种质资源中筛选耐盐材料以及通过常规育种手段改良现有水稻品种的耐盐性,虽取得了一定的进展,但这些品种所表现出的耐盐能力不强,满足不了盐碱地生产的要求。随着现代分子生物技术的快速发展,利用发现和克隆到的盐生植物的耐盐基因,培育耐盐水稻新品种是提高水稻耐盐性,维持盐碱地农业生产的重要途径之一[12-13]。尽管利用生物工程技术将特定外源基因导入水稻后,一定程度上提升了水稻的耐盐能力[14-16],但因为耐盐性是受多数数量性状基因控制的复杂性状,转移单个基因对提高水稻耐盐性的作用比较有限,仍然难以达到盐碱地水稻生产的要求。因此,同时转化多个耐盐基因及其调控因子,是大幅提高作物耐盐性的必要途径。
双元细菌人工染色体(bibary bacterial artificial chromosome,BIBAC)載体可以在大肠杆菌和根癌农杆菌中复制,与一般载体相比,除了能构建大片段文库外,还能直接用于大片段DNA、多基因或基因簇的遗传转化,来促进基因发现和功能研究[17-18]。利用BIBAC载体已成功构建了番茄[19]、野生稻[20]、粳稻[21]、盐芥[22]等基因文库,为后续优良基因的发掘与利用奠定了基础。同时,利用BIBAC载体将大片段成功转入植株基因组中,如Hamilton等[23]和Frary等[24]利用BIBAC载体将150 kb外源片段通过农杆菌介导法转入烟草和番茄基因组中,He等[20]将携带褐飞虱抗性基因的120 kb片段-BIBAC载体成功导入水稻基因组中。
一种生长于西北盐盖上的野生芦苇,具有极强的耐盐性,与水稻同属禾本科,对提高水稻的耐盐性具有极高的潜在价值。本研究以此耐盐野生芦苇为材料,构建了耐盐芦苇大片段DNA- BIBAC载体,利用农杆菌介导法转化水稻愈伤组织,并成功获得了转基因植株。耐盐芦苇大片段DNA转化水稻技术为创制耐盐水稻新种质,以及后续克隆耐盐芦苇的耐盐基因奠定了基础,同时对于水稻耐盐性的提高、盐碱地的有效利用等具有潜在的重要价值。
1 材料与方法
1.1 材料
供体材料为一种生长于西北盐盖上的野生芦苇。pHZAUBIBAC1载体由华中农业大学罗美中教授惠赠;大肠杆菌菌株DH10B,购自上海唯地生物科技有限公司;根癌农杆菌EHA105,购自北京华越洋生物科技有限公司;CIP(小牛肠碱性磷酸酶)、T4 DNA连接酶、Lambda PFG ladder购自NEB公司;BamH I、λDNA、I-Sce I购自Thermo Scientific公司;质粒提取试剂盒购自OMEGA公司;用于转化的材料粳稻‘中花11(Oryza satva L. subsp. japonica cv Zhonghua11)以及209转基因‘中花11质粒由海南波莲水稻基因有限公司提供。
1.2 方法
1.2.1 BIBAC-S载体质粒的制备 取?80 ℃冻存的pHZAUBIBAC1菌株于LB固体平板上进行划线培养,再挑选单菌落于含有12.5 ?g/mL Amp和Kan的LB液体培养基中振荡培养,待菌液处于对数生长期,利用OMEGA试剂盒进行载体质粒提取,微量分光光度计及凝胶电泳检测提取的质粒质量;将提取的载体质粒置于37 ℃下酶切2.5 h;酶切完成后,37 ℃下脱磷50 min,随后进行纯化及自连反应;将上述已完成自连的载体DNA进行1%脉冲场凝胶电泳分析,通过凝胶成像平行切割出含有目的载体BIBAC-S的凝胶块,装入无菌并含有100 μL 0.5×TBE透析袋中(8000~ 14000 D)进行电洗脱回收,检测回收液的质量。脉冲场凝胶电泳条件:泵速70,6 v/cm,1~50 s,120°,0.5×TBE,14 ℃,14 h;电洗脱回收条件:4 ℃,125 V,2.5 h,电泳结束后180°颠倒透析袋,继续电泳2 min。
1.2.2 芦苇大片段DNA的制备 参照文献[25]来制备芦苇大片段DNA,并从脉冲场凝胶电泳后的凝胶中切割出含有50~300 kb大小的芦苇DNA片段的凝胶块,电洗脱回收凝胶中的DNA大片段,电洗脱条件同1.2.1。检测回收DNA的浓度,确定目的DNA浓度大于1 ng/μL才能进行后续连接实验。
1.2.3 芦苇大片段DNA-BIBAC载体构建 将回收的BIBAC-S载体和不同大小的芦苇大片段DNA通过T4连接酶16 ℃连接16 h;将连接产物吸入至脱盐胶,静置于冰中1.5 h,进行脱盐处理;脱盐处理后的连接产物通过电击转化至大肠杆菌DH10B中;涂板培养,挑选阳性克隆摇菌,利用碱裂解法提取克隆质粒;利用I-Sce I对BIBAC克隆质粒进行酶切,脉冲场凝胶电泳检测,获取插入不同大小的芦苇大片段DNA阳性克隆。脉冲场电泳条件:泵速70,6 v/cm,5~15 s,120°,0.5× TBE,14 ℃,16 h。
1.2.4 BIBAC载体遗传转化水稻愈伤组织 粳稻‘中花11种子去壳,用25%的NaClO消毒15~20 min,无菌水清洗7~8次后平铺于无菌滤纸上风干30 min,接种到诱导培养基,33~34 ℃恒温暗培养35~40 d,将种子胚周围新生长出的愈伤组织进行继代培养;从继代培养基中挑选出颜色嫩黄、表面干燥、生长旺盛的愈伤组织转移至预/共培养基(含100 μmol/L乙酰丁香酮)上,25 ℃暗培养4 d;用含有不同片段BIBAC载体的EHA105菌液(OD600值为0.4~0.6,含100 μmol/L乙酰丁香酮)侵染25 min,置于无菌滤纸风干,转移至预/共培养基中25 ℃暗培养3 d;用灭菌超纯水将共培养愈伤组织上的农杆菌冲洗干净,再加入500 mg/L的头孢水浸泡25~30 min,风干愈伤后,转移至筛选培养基(含50 mg/L的Hyg和500 mg/L的头孢)中,28 ℃暗培养20~50 d,期间更换一次培养基;将抗性愈伤组织转入分化培养基(含2 mg/L KT和2 mg/L NAA)中,置于25 ℃光照培养20~50 d;将分化出小苗的根清理干净后,转移至1/2 MS生根培养基中,25 ℃光照培养15 d;解开封口膜,加入2~3 cm的自来水后培养2~3 d进行炼苗,移苗完成后移栽至水稻池中。诱导与继代培养基成分:100 mL/L 10×N6大量、10 mL/L 100×N6微量+100× N6有机+100× N6铁盐、2,4-D(1 mg/mL)、0.5 g/L脯氨酸、0.6 g/L水解络蛋白、30 g/L蔗糖、3 g/L植物凝胶,pH 5.9,116 ℃灭菌15 min,其中2,4-D(1 mg/mL)在诱导培养基与继代培养基中的加入量分别为3 mL/L和2.4 mL/L。
1.2.5 PCR筛选遗传转化植株 CTAB法提取转化植株的DNA,用潮霉素和水稻内参基因Actin引物进行PCR扩增鉴定转基因植株。引物序列见表1。PCR反应体系均为:0.4 μL模板DNA、5 μL 2×Taq MasterMix(Dye)、0.25 μL 10 μmol/L Hn-F/R引物、(或0.4 μL 10 μmol/L LH-F/R引物),用灭菌双蒸水补足10 μL。PCR反应条件均为:94 ℃预变性5 min;94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸25 s,共进行30个循环;72 ℃延伸5 min。PCR反应产物进行1%琼脂糖凝胶电泳检测,确定转基因阳性植株。2对引物的扩增长度分别为664 bp和110 bp。
1.3 数据分析
记录农杆菌介导不同片段大小的BIBAC载体在转化水稻过程中抗性愈伤数目、再生植株数目以及阳性植株数目,并计算出各自最终的遗传转化效率。愈伤转化率=抗性愈伤数目/侵染的总愈伤数目×100%;植株转化率=再生植株数目/接种的抗性愈伤总数目×100%;植株阳性率=阳性植株数目/检测植株总数目×100%;终转化效率=愈伤转化率(%)×植株转化率(%)×植株阳性率(%)× 100%。
2 结果与分析
2.1 BIBAC-S载体质粒与芦苇大片段DNA的制备
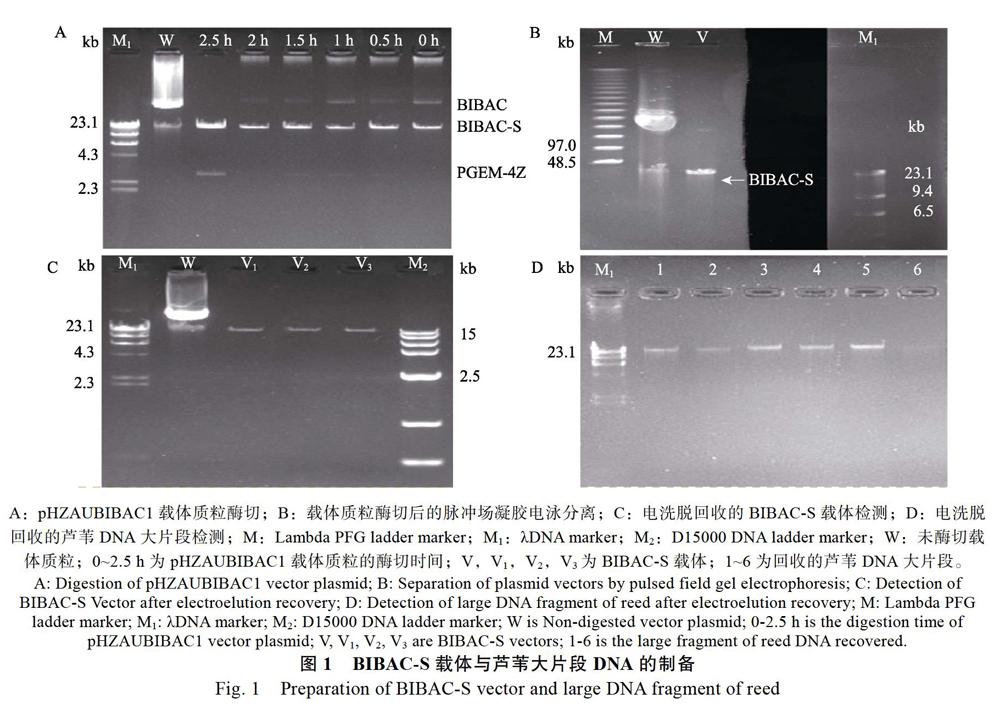
经摇菌培养后提取的pHZAUBIBAC1载体质粒浓度约为400 ng/μL左右,并且DNA条带清晰,纯度较高。不同时间的酶切效果显示,当酶切至2.5 h时,载体质粒酶切完全,其浓度约为25 ng/μL左右(图1A)。依据此条件,对pHZAUBIBAC1载体质粒酶切2.5 h,经脱磷纯化后进行脉冲场电泳分离,从图1B中可知BIBAC-S载体条带亮度明显,说明载体浓度较高,并且BIBAC-S与PGEM-4Z载体已经完全分离;图1B中黑色部分的凝胶在显色前被平行切割出来,用于电洗脱回收BIBAC-S载体,该部分凝胶中含有与“V”列相同的BIBAC-S载体质粒。电洗脱回收的BIBAC-S载体经电泳检测,回收的BIBAC-S载体质粒纯度高,浓度约为20 ng/μL,符合载体连接时的要求(图1C)。同时,对提取的Mb级耐盐芦苇基因组DNA经酶切、脉冲场电泳分离、凝胶切割、电洗脱回收,回收的载体浓度在2~4 ng/μL左右,满足与载体连接时的要求(图1D)。
2.2 芦苇大片段DNA-BIBAC载体的构建
在电洗脱回收的载体DNA与目的大片段DNA连接转化后的筛选平板中,随机挑选12个白斑单菌落摇菌,提取质粒,I-Sce I单酶切,酶切产物经脉冲场凝胶电泳检测。从图2可知,12个单克隆均为阳性克隆,均含有插入片段且不存在空载现象,插入片段大小介于50~150 kb之间,不存在双克隆和多克隆的现象,总体来说芦苇大片段DNA-BIBAC载体构建非常成功。随后,将构建好的载体转化农杆菌EHA105,用于水稻的遗传转化。
2.3 芦苇大片段DNA-BIBAC载体遗传转化水稻
水稻愈伤组织的诱导与继代:水稻种子接种到愈伤组织诱导培养基上,33~34 ℃条件下暗培养40 d长出愈伤组织(图3A);愈伤组织继代培养(图3B)。抗性愈伤组织的筛选:农杆菌与愈伤组织在25 ℃条件下暗培养4 d后,转移至筛选培养基,一部分愈伤组织会褐化死亡,一部分会长出新的米黄色抗性愈伤组织(图3C)。抗性愈伤组织的分化:转移抗性愈伤组织于分化培养基中分化,愈伤会慢慢分化变大,并逐渐转绿,最后分化出幼苗(图3D)。抗性转化苗的生根与移栽:将分化出的幼苗根部清理干净,转移至生根培养基上进行生根培养(图3E);待幼苗复壮后炼苗3 d,移栽至室外水稻池中(图3F)。
2.4 遗传转化苗的PCR鉴定
提取转基因苗叶片DNA,用潮霉素(Hn664)基因和水稻内参(Actin110)基因引物进行PCR扩增,部分扩增结果如图4所示。由图4可知,已转化成功的水稻植株能扩增出与阳性对照相同大小的条带,证明农杆菌介导的BIBAC载体已经成转入粳稻‘中花11中。
2.5 大片段DNA-BIBAC载体转化效率
本试验转化了3组不同大小的大片段DNA- BIBAC载体,其大小分别为50~75 kb、60~90 kb和90~120 kb,遗传转化结果如表2所示。由表2可知,获得阳性植株总数为23株,3组不同大小的大片段DNA-BIBAC载体的最终转化效率分别为7.71%、7.23%和2.99%,初步的转化数据显示,插入DNA片段越大转化效率越低。
3 讨论
利用大片段DNA转化技术或原位的基因组导入技术向栽培作物中导入有益性状进行遗传转化是作物改良中的有益实践。许多研究已经证实外源基因的大片段DNA能稳定地导入植物基因组。通过对许多禾本科植物基因组的比较,已经发现含有功能基因的大片段DNA在远缘物种间是可以横向传递的,并且被受体物种接受使用,扩展了遗传基础,这可能是植物进化的重要驱动力[26]。
盡管存在争议,但农杆菌介导的转化系统由于其T-DNA的精确处理和低拷贝数转基因的简单整合,已成为生产转基因水稻品种的较好选择。大片段DNA转化过程中的许多因素决定了转化效率或者直接导致试验失败。外植体鉴定、基因转移技术、整合构建、无遗传损伤的转基因表达和转化子选择是影响水稻转化的技术难题,获取大片段DNA转化过程中的这些信息对于改良难驯化水稻品种的转化体系是必要的[27]。本研究结果显示,pHZAUBIBAC1的酶切对于后续实验极为重要,若酶切不完全,那么pHZAUBIBAC1载体会夹杂其中,降低线性BIBAC-S载体回收的纯度,从而影响到后续BIBAC-S载体与大片段DNA的连接效率,在进行DH10B转化时,空载体会优先于重组质粒进入大肠杆菌,产生大量蓝斑,影响筛选。酶切之后的脱磷也对后续实验起着关键性作用,脱磷时间过长会降低连接效率,脱磷时间过短会让载体产生自连,2种情况都会产生数目较多的蓝斑,经多次摸索试验,脱磷的最适时间在55~65 min之内。脱磷后利用电洗脱的方法来进行载体与芦苇大片段的回收,与试剂盒回收比较起来,此方法回收的浓度会比试剂盒回收的载体浓度低,但是其回收纯度高,能够最小限度地对目的载体和大片段造成机械损伤,保证其完整性,可有效提高连接效率。
利用农杆菌EHA105介导粳稻中花11的转化过程中,选择生长旺盛的愈伤组织作为转基因的受体能够提高转化效率[28];同时选择合适的菌液浓度与侵染时间也至关重要[29]。有研究认为,在BIBAC系统中,高浓度的农杆菌细胞可提高转化效率,若农杆菌浓度过低且侵染时间过短会出现侵染不充分,导致转化效率变低且筛选时大量愈伤发生褐变死亡的情况[30]。本研究中,侵染粳稻‘中花11的菌液浓度OD600为0.4~0.6之间,侵染时间为25 min。若农杆菌浓度过高且侵染时间过长会导致大量农杆菌增殖,对植物细胞造成伤害且脱菌困难,降低愈伤组织的再生频率且容易导致后续实验出现农杆菌污染的情况。此外,有研究表明水稻中可能缺少对农杆菌Vir基因活化所需的酚类物质,因此表现出对农杆菌并不敏感,所以在悬浮液和预/共培养基中加入少量的乙酰丁香酮(AS)能够有效提升转化效率[31],是转化的必要条件。
本研究通过农杆菌介导法,将耐盐芦苇不同大片段DNA-BIBAC载体对粳稻‘中花11进行遗传转化,经过PCR扩增初步鉴定获得阳性植株23株,实现了农杆菌介导的大片段DNA在水稻中的遗传转化,为创制耐盐水稻新种质,以及后續挖掘和克隆芦苇耐盐相关基因奠定了基础。但从最终的转化效率来看,不同大小的大片段DNA载体在转化同一受体材料时,因其所获得的愈伤转化率、植株转化率、转基因植株的阳性率等存在差异,说明插入DNA片段的大小与转化效率之间有一定关系。今后在创制耐盐水稻新种质的时候,既要考虑一次转入片段的大小,在导入更多基因的同时,也要考虑遗传转化的效率,从而获得更多的突变体。
参考文献
[1] Li B, Takahashi D, Kawamura Y, et al. Comparison of plasma membrane proteomic changes of Arabidopsis suspension-cultured cells (T87 line) after cold and ABA treatment in association with freezing tolerance development[J]. Plant Cell Physiology, 2012, 53(3): 543-554.
[2] Wang X, Wang J G, Liu H L, et al. Influence of natural saline-alkali stress on chlorophyll content and chloroplast ultrastructure of two contrasting rice (Oryza sativa L. Japonica) cultivars[J]. Australian Journal of Crop Science, 2013, 7(2): 289-292.
[3] Zhu J K. Plant salt tolerance[J]. Trends in Plant Science, 2001, 6(2): 66-71.
[4] Tian L, Tan L B, Liu F X, et al. Identification of quantitative trait loci associated with salt tolerance at seedling stage from Oryza rufipogon[J]. Journal of Genetics and Genomics, 2011, 38(12): 593-601.
[5] Wang X C, Chang L L, Wang B C, et al. Comparative proteomics of Thellungiella halophila leaves from plants subjected to salinity reveals the importance of chloroplastic starch and soluble sugars in halophyte salt tolerance[J]. Molecular and Cellular Proteomics, 2013, 12(8): 2174-2195.
[6] Zarei M, Paymaneh Z. Effect of salinity and arbuscular mycorrhizal fungi on growth and some physiological parameters of Citrus jambheri[J]. Archives of Agronomy and Soil Science, 2014, 60(7): 993-1004.
[7] 李 刚, 袁彩勇, 王 健, 等. 水稻新品种淮香粳15号丰产性、稳产性及适应性分析[J]. 福建农业学报, 2016, 31(2): 113-117.
[8] Kronzucker H J, Szcerba M W, Schulze L M, et al. Non-reciprocal interactions between K+ and Na+ ions in barley (Hordeum vulgare L.)[J]. Journal of Experimental Botany, 2008, 59(10): 2793-2801.
[9] Schmidt R, Mieulet D, Hubberten H M, et al. SALT-RESPONSIVE ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice[J]. Plant Cell, 2013, 25(6): 2115-2131.
[10] Yamaguchi T, Blumwald E. Developing salt-tolerant crop plants: challenges and opportunities[J]. Trends in Plant Science, 2005, 10(12): 615-620.
[11] 郑崇珂, 窦玉慧, 解丽霞, 等. 水稻耐盐相关基因的研究进展[J]. 分子植物育种, 2017, 15(11): 4411-4422.
[12] 刘士平, 李 信, 汪朝阳, 等. 基因聚合对水稻稻瘟病的抗性影响[J]. 分子植物育种, 2003, 1(1): 22-26.
[13] 郭龙彪, 程式华, 钱 前. 水稻基因设计育种的研究进展与展望[J]. 中国水稻科学, 2008, 22(6): 650-657.
[14] 王彩芬, 刘冬成, 马晓玲, 等. 水稻耐盐基因SKC1特异性CAPS标记的开发与验证[J]. 分子植物育种, 2015, 13(11): 2437-2440.
[15] 李荣田, 张忠明, 张启发. RHL基因对粳稻的转化及转基因植株的耐盐性[J]. 科学通报, 2002, 47(8): 613-617.
[16] Hou X, Xie K, Yao J, et al. A homolog of human ski-interacting protein in rice positively regulates cell viability and stress tolerance[J]. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(15): 6410-6415.
[17] Shibata D, Liu Y G. Technical Focus-Agrobacterium-me-diated plant transformation with large DNA fragments[J]. Trends in Plant Science, 2000, 5(8): 354-357.
[18] 何瑞锋, 王媛媛, 杜 波, 等. 水稻双元细菌人工染色体载体系统转化体系的建立[J]. 遗传学报, 2006, 33(3): 269-276.
[19] Hamilton C M, Frary A, Xu Y, et al. Construction of tomato genomic DNA libraries in a binary-BAC (BIBAC) vector[J]. Plant Journal, 1999, 18(2): 223-229.
[20] He R F, Wang Y Y, Shi Z Y, et al. Construction of a genomic library of wild rice and Agrobacterium-mediated transformation of large insert DNA linked to BPH resistance locus[J]. Gene, 2003, 321: 113-121.
[21] Tao Q, Wang A, Zhang H B. One large-insert plant-transformation-competent BIBAC library and three BAC libraries of japonica rice for genome research in rice and other grasses[J]. Theoretical and Applied Genetics, 2002, 105(6-7): 1058-1066.
[22] Wang W, Wu Y, Li Y, et al. A large insert Thellungiella halophila BIBAC library for genomics and identifi cation of stress tolerance genes[J]. Plant Molecular Biology, 2009, 72(1/2): 91-99.
[23] Hamilton C M, Frary A, Lewis C, et al. Stable transfer of intact high molecular weight DNA into plant chromosomes[J]. Proceedings of the National Academy of Sciences of the United States of America, 1996, 93(18): 9975-9979.
[24] Frary A, Hamilton C M. Efficiency and stability of high molecular weight DNA transformation: an analysis in tomato[J]. Transgenic Research, 2001, 10(2): 121-132.
[25] 杨 俊, 高 鸿, 周申遒, 等. 基于BIBAC文库的芦苇高分子量核DNA的制备方法[J]. 分子植物育种, 2017, 15(11): 4578-4584.
[26] Dunning L T, Olofsson J K, Parisod C, et al. Lateral transfers of large DNA fragments spread functional genes among grasses[J]. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(10): 4416-4425.
[27] Mohammed S, Samad A A, Rahmat Z. Agrobacterium-me-diated transformation of rice: Constraints and possible solutions[J]. Rice Science, 2019, 26(3): 133-146.
[28] 刘亮亮, 唐 维, 喻 旭, 等. 根癌农杆菌介导的籼稻蜀恢527遗传转化方法的探究[J]. 中国农业科技导报, 2012, 14(4): 135-141.
[29] 李朝炜, 冯 丹, 翟会青, 等. 农杆菌介导的旱稻遗传转化主要影响因素[J]. 湖北农业科学, 2014, 53(15): 3482-3487.
[30] 孙华军, 李国瑞, 陈永胜, 等. 农杆菌介导的植物遗传转化影响因素研究进展[J]. 安徽农业科学, 2015, 43(24): 26-27, 77.
[31] Aldemita R R, Hodges T K. Agrobacterium tumefaciens-mediated transformation of japonica and indica rice varieties[J]. Planta, 1996, 199(4): 612-617.
责任编辑:崔丽虹
收稿日期 2020-06-12;修回日期 2020-07-17
基金项目 国家自然科学基金项目(No. 31660381);国家科技支撑计划项目(No. 2015BAD01B02-1-1)。
作者简介 高 鸿(1994—),男,硕士,研究方向:植物生物技术与种质创新。*通信作者(Corresponding author):馬启林(MA Qilin),E-mail:hbhnqlm@163.com。
猜你喜欢
海外文摘·文学版(2022年4期)2022-04-14
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
现代妇女(2019年2期)2019-02-24
岁月(2018年2期)2018-02-28
农产品市场周刊(2017年35期)2017-10-25
天津农业科学(2016年12期)2017-01-11
山东农业科学(2016年11期)2016-12-17
山东农业科学(2016年11期)2016-12-17
青年文摘·上半月(1996年1期)1996-12-31
