
鱼游泳能力对体长的响应及其在鱼道设计中的应用
2021-05-12 07:19:36侯轶群胡望斌陈小娟陈继华黄应平韩德举
农业工程学报 2021年5期
蔡 露,侯轶群,金 瑶,杨 志,胡望斌,陈小娟,陈继华,黄应平,韩德举
鱼游泳能力对体长的响应及其在鱼道设计中的应用
蔡 露1,2,侯轶群1,金 瑶1,杨 志1,胡望斌1,陈小娟1,陈继华2,黄应平2,韩德举1※
(1. 水利部中国科学院水工程生态研究所,水利部水工程生态效应与生态修复重点实验室,武汉 430079; 2. 三峡大学水利与环境学院,三峡库区生态环境教育部工程研究中心,宜昌 443002)
为了探讨鱼类体长对游泳能力的影响并为鱼道水流的设计提供参考,该研究在封闭水槽中使用“递增流速法”测试了海南省某水利枢纽鱼道目标对象的游泳能力,并用Origin软件进行了数据统计分析,得到了试验鱼感应流速、临界游泳速度和爆发游泳速度的直线回归方程和Kaplan-Meier曲线。结果表明:1)随着试验鱼体长增大,相对感应流速、临界游泳速度和爆发游泳速度(体长/s)均减小,体长和鱼类速度的相关关系可用直线方程表示,且数据经过对数变换后的直线方程拟合效果比未经过对数变换的拟合效果更好,其中2由0.664~0.725提高至0.907~0.933。2)根据鱼道设计规范、导则及文献,结合本工程目标过鱼对象的感应流速、临界游泳速度和爆发游泳速度(m/s),建议本工程鱼道进口诱鱼流速控制在0.35~0.47 m/s,池室流速控制在0.21~0.59 m/s,竖缝流速控制在0.57~0.74 m/s,出口断面至下一个池室之间的流速控制在0.21~0.50 m/s。鱼类体长对相对游泳速度(体长/s)产生了负面影响,鱼类游泳速度及其变化规律可对鱼道水流设计值提供参考。
流速;体长;游泳能力;感应流速;临界游泳速度;爆发游泳速度;竖缝式鱼道
0 引 言
水利水电工程具有防洪、发电、航运等重要功能,在带来巨大的经济和社会效益的同时,也破坏了河流连通性[1],改变了营养物的转移规律[2]和鱼类栖息环境[3],阻断了鱼类洄游及其种群之间的基因交流[4-6]。修建鱼道可以缓解工程阻隔的负面作用[7-8]。鱼道的目标是要帮助鱼类越过大坝的阻隔,但是其后续的最终目标是要让鱼类能够成功的繁衍后代[9]。虽然鱼道理应帮助所有游过此处的鱼顺利通行,但由于不同鱼类有着不同游泳能力,因此鱼道很难同时满足所有鱼类的过坝需求。为此,应优先考虑帮助洄游型鱼种、国家或地方性保护鱼种、地方特有鱼种等鱼类进行过坝,鱼道设计应优先满足这部分鱼种的需求。鱼道进口流速研究、鱼类通过鱼道能力的定量研究、鱼类自主运动水槽的改进优化等方面是目前鱼类运动行为和鱼道研究的重点[10-11]。鱼类体长是影响游泳能力的重要因素[9],游泳能力是鱼道设计的关键基础数据[12-13],其中鱼类爆发游泳速度、临界游泳速度和感应流速是重要的参考指标[12-16]。目前很多鱼道在设计过程中仍然缺乏针对性的鱼类游泳能力数据[7,13]。因此,本研究旨在探讨鱼类体长对游泳能力的影响,并依据鱼类游泳能力研究结果来设计目标鱼道的水流。
本研究以海南省某水利枢纽工程为目标工程。《海南省某水利枢纽环境影响报告书》建议该工程过鱼设施的类型为竖缝式鱼道,过鱼目的主要是为了帮助鱼类产卵洄游和促进基因交流,主要过鱼对象选取为花鳗鲡(国家保护种)、台细鳊(中国物种红色名录)、锯齿海南䱗(中国物种红色名录)、海南石鲋(省特有鱼类)、条纹刺鲃(省特有鱼类),兼顾过鱼对象选取为3种经济型鱼类胡子鲇、斑鳢、三角鲤。本研究测试之前,依据文献[16]建议的过鱼设施过鱼对象筛选方法,根据鱼类洄游习性、生境适宜性、保护价值、经济价值和资源量状况等方面对目标过鱼对象进行了复核,认为上述环境影响评价报告书中确定的过鱼设施过鱼对象是适宜的。由于花鳗鲡成鱼属于下行洄游,鱼道的水流速度并不对其构成障碍,当其抵达鱼道时可以顺水而下,因此本研究鱼道流速设计需要的花鳗鲡数据为上行洄游的幼鳗的游泳能力数据。其他需要的鱼类数据为常见规格的洄游个体游泳能力数据。
1 材料与方法
1.1 试验鱼
本试验测试对象本应为花鳗鲡(幼)、台细鳊、锯齿海南䱗、海南石鲋、条纹刺鲃、胡子鲇、斑鳢、三角鲤。由于近年来部分鱼类野生资源量急剧下降而未能捕获所需规格的花鳗鲡和台细鳊,因此本研究采用在分类学上和栖息环境均相近且理应具有相似游泳能力[9,17]的日本鳗鲡和海南华鳊进行代替测试。获取目标鱼后,先将鱼暂养于静水水箱24~48 h,减轻鱼类的应激反应,然后再进行游泳能力测试。试验鱼规格见表1,每种鱼均进行30尾试验。
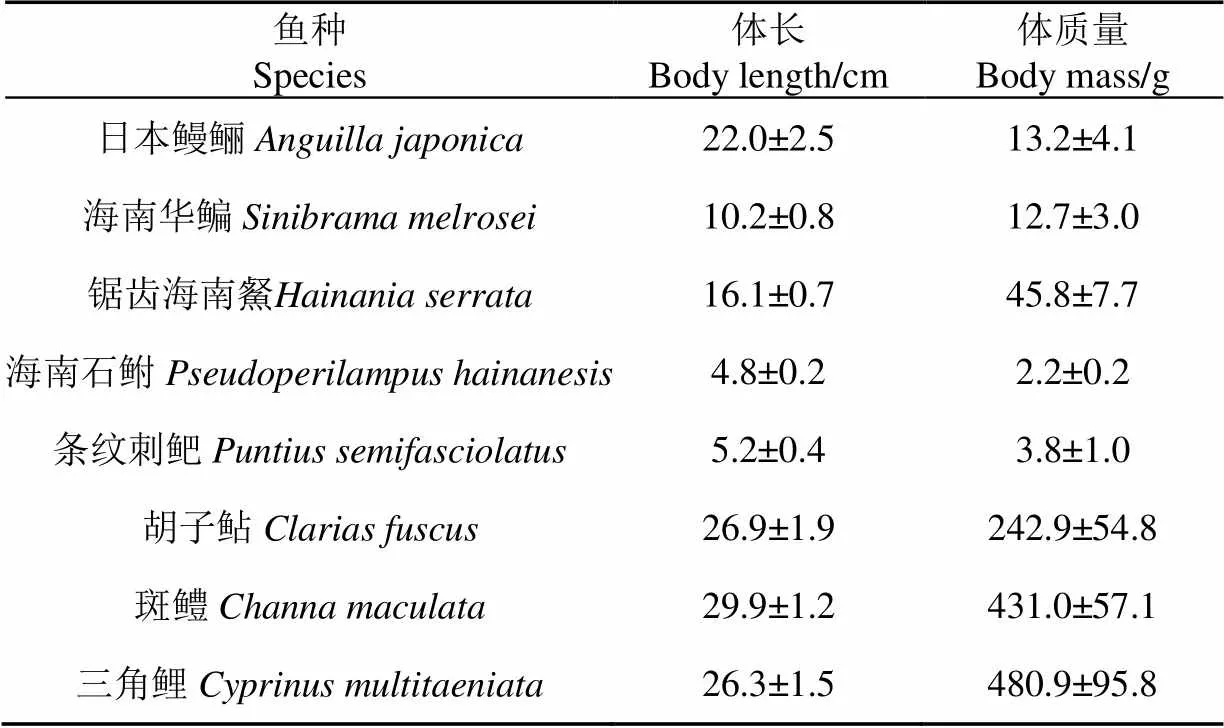
表1 试验鱼规格(平均值±标准差)
1.2 试验设备
文献[18]利用封闭式循环水槽测试了几种鱼类的感应流速并应用于过鱼设施最低流速的设计,本研究利用该装置测了鱼类的感应流速、临界游泳速度和爆发游泳速度并应用于过鱼设施的进口、池室、竖缝和出口流速的设计。鱼类在该装置游泳区内运动,因而可以认为水流速度等于鱼类游泳速度,通过测试水流速度即可推算鱼类游泳速度。用于测试的鱼类为现场捕获,测试在室内进行,室内光照为昼夜自然光周期,使用空气泵向试验水体中泵气以提高溶解氧含量,试验水温为当地自然水温。利用溶氧仪(YSI DO200A,Yellow Springs,USA)监测水中的温度和溶解氧含量,试验水体溶解氧含量范围6.05~6.80 mg/L,水温范围29.1~31.8 ℃。
1.3 试验方法
使用国际通行方法“递增流速法”[18-20]测试鱼类感应流速(试验样本重复数=10)、临界游泳速度(=10)和爆发游泳速度(=10)。在测试前,对所有试验鱼均进行1 h的适应,从而减缓试验鱼对试验设备的应激性。通常递增流速法中的递增速度增量可以以绝对速度m/s或相对速度bl/s(bl为鱼类体长)为单位基准。由于各种鱼类感应流速的绝对值通常较小且差异不大,使用绝对速度m/s为单位基准可以使所有测试过程均在相同的绝对初始速度和绝对递增速度的条件下进行。绝对临界游泳速度和绝对爆发游泳速度通常较大且差异较大,如果使用绝对速度m/s为单位基准则会使得部分鱼类测试总时间过短和部分鱼类测试总时间过长,从而影响试验结果,因此为了能尽量避免上述影响,本研究使用相对速度bl/s为单位基准进行测试。试验中鱼类感应流速、临界游泳速度和爆发游泳速度3种速度测试的递增流速法试验参数可见表2。
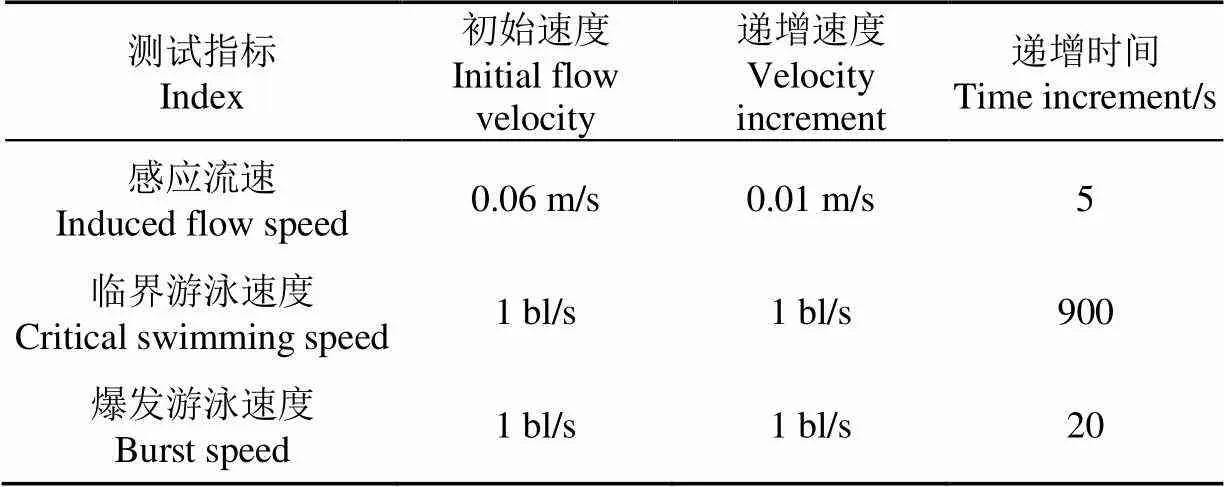
表2 递增流速法试验参数
注:bl为鱼类体长。
Note: The bl is fish body length.
1.4 数据分析
本试验数据分析使用Origin 9.0软件进行。鉴于相同规格大小的相似鱼种有着相似的游泳能力[9,17],因此为了分析本研究中鲤科鱼类游泳能力状况,可利用直线回归方程直接对本研究中鲤科鱼类游泳能力数据(感应流速、临界游泳速度和爆发游泳速度)和体长的相关关系进行拟合[18],得到拟合参数2、、。同时,对游泳能力数据和体长数据进行ln变换,然后利用直线回归方程进行拟合[21],得到拟合参数2、、。对比数据转换前后所得的拟合参数,分析数据转换前后直线方程的拟合效果。
使用Kaplan-Meier曲线(或称生存曲线Survival curves)表示在感应流速测试时的某流速条件下可感应到水流方向并进行逆流运动的鱼类的数量百分比,以及在临界游泳速度或爆发游泳速度测试时的某流速条件下未疲劳的鱼类的数量百分比[22-24]。然后分别计算感应流速、临界游泳速度和爆发游泳速度的EF90值。EF90值,即某流速设计值对90%的鱼有效的参数数值,感应流速EF90值表示90%的鱼可以感应到该流速并逆流运动,临界游泳速度EF90值或爆发游泳速度EF90值表示90%的鱼在该测试条件下未达到运动疲劳[16]。
2 结果与分析
2.1 鱼类游泳能力与体长的相关关系
由于不同鱼种的体长不同,为了从一定程度上排除体长的影响,国内外在研究鱼类游泳能力时通常可用相对速度(bl/s)来表达研究结果和发现[25-28]。本研究结果显示:随着试验鲤科鱼类体长增大,相对感应流速、相对临界游泳速度和相对爆发游泳速度均减小(表3,图1)。由于鱼类的体长会影响其游泳能力,因此为了能良好地进行鱼道流速设计,试验鱼规格应尽量包含所有目标过鱼规格的范围。表3展示了试验鱼的3种相对速度与体长的直线函数拟合关系以及经过ln数据变换后的直线函数拟合关系。通过对比函数拟合参数(2和)说明,经过ln数据变换后的直线函数拟合关系的拟合效果比未经过ln变换的拟合效果更好,其中2由0.664~0.725提高至0.907~0.933,即2提高了0.182~0.266。虽然经过ln数据变换后的鱼类相对速度(bl/s)的拟合效果能够得到明显的提升,但该数据处理方法对鱼类绝对速度(m/s)未能起到明显的效果。
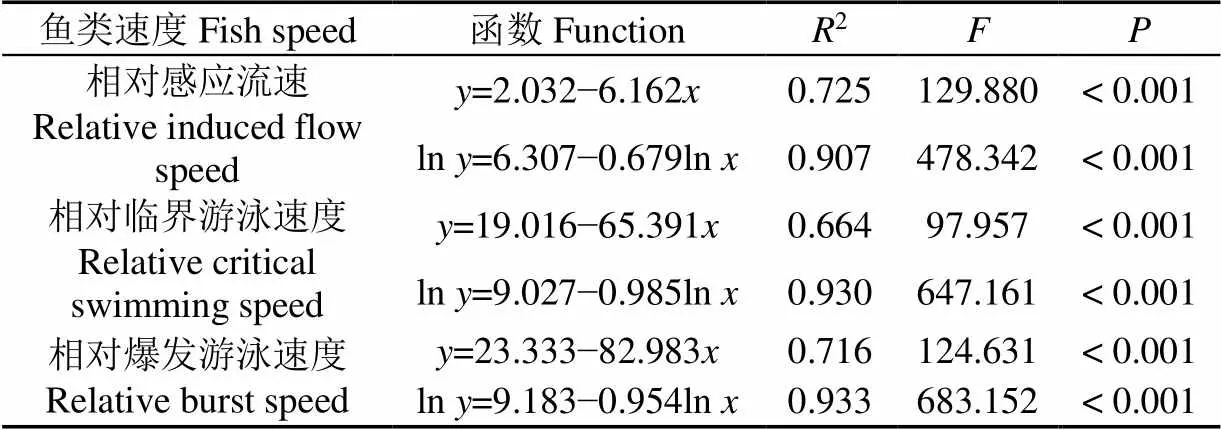
表3 试验鲤科鱼类的游泳能力指标与体长的直线函数关系
注:为鱼类速度(bl/s),为鱼类体长,cm。
Note: Theis fish speed (bl/s) and theis fish body length, cm.
2.2 基于鱼类游泳能力的竖缝式鱼道流速设计
将所有试验鱼测试结果的相对速度(bl/s)进行单位换算,可得到所有试验鱼的绝对感应流速、绝对临界游泳速度和绝对爆发游泳速度(m/s)的平均值(表4)和箱线图(图2)。
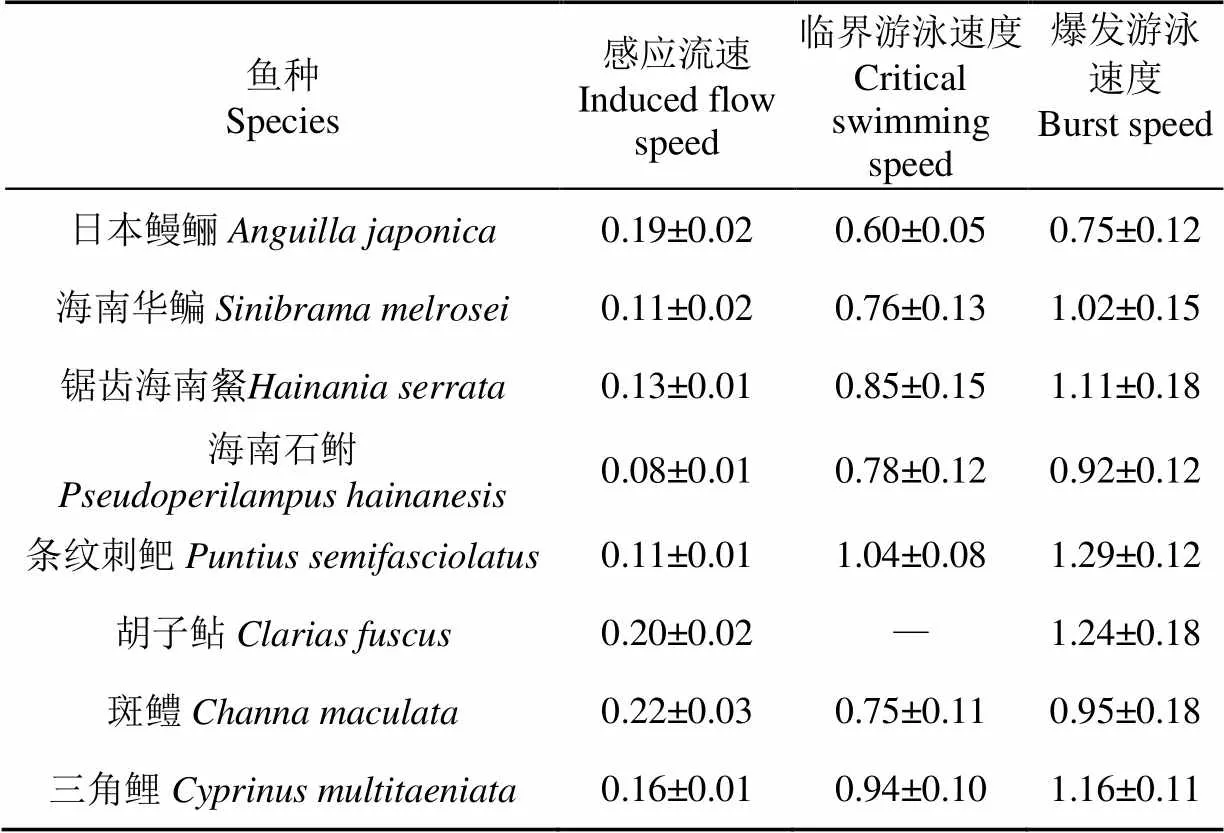
表4 试验鱼游泳能力(平均值±标准差)
图3展示了在感应流速测试的某流速条件下,可感应到水流方向并进行逆流运动的鱼类的数量百分比;和在临界游泳速度或爆发游泳速度测试的某流速条件下未疲劳的鱼类的数量百分比。依据图3的Kaplan-Meier曲线,当百分比为10%时,感应流速为0.21 m/s;当百分比为90%时,临界游泳速度为0.59 m/s;当百分比为90%时,爆发游泳速度为0.74 m/s(即该3项速度值分别为鱼类速度的3种EF90值)。上述3项值也在图2中以虚线形式描绘用以展示分布情况。感应流速EF90值接近最大感应流速,临界游泳速度EF90值接近最小临界游泳速度,爆发游泳速度EF90值接近最小爆发游泳速度,结合前文EF90值的定义可以认为,利用本研究3项EF90值设计的过鱼设施流速可从理论上满足90%的试验鱼需求。
本次鱼类游泳能力测试的主要结果: 感应流速EF90值(Uind-EF90)为0.21 m/s,临界游泳速度EF90值(Ucrit-EF90)为0.59 m/s,爆发游泳速度EF90值(Uburst-EF90)为0.74 m/s,均可用于设计竖缝式鱼道进口(下游)、池室、竖缝和出口(上游)的流速。基于鱼类游泳能力测试结果,针对目标工程竖缝式鱼道流速设计提供如下建议:
1)进口流速。根据中国《水利水电工程鱼道设计导则》和《水电工程过鱼设施设计规范》[29-30]的建议:鱼道进口区域水流速度小于鱼类感应流速时,应采取补水等措施。但该导则和规范并未对补水量及流速进行量化分析。有文献[31]调研并总结了一些鱼类的趋流行为,并认为鱼道进口吸引流速宜为0.6~0.8倍临界游泳速度左右。根据本研究结果,过鱼对象临界游泳速度EF90值为0.59 m/s,因此换算到本工程鱼道,建议进口诱鱼流速控制在0.35~0.47 m/s(0.6~0.8 Ucrit-EF90)。
2)池室流速。《水利水电工程鱼道设计导则》和《水电工程过鱼设施设计规范》建议鱼道池室流速不应大于鱼类爆发游泳速度。鱼道的流速应能够保证鱼类避免迷失方向[9]。鱼类临界游泳速度反映了其最大可持续游泳速度[19],考虑到鱼类通常需要至少游过几百米长的鱼道(约100~500个池室),所以鱼道流速设计必须考虑鱼类的运动耐力。因此,本研究建议鱼道池室流速控制在0.21~0.59 m/s(Uind-EF90~Ucrit-EF90)。
3)竖缝流速。《水利水电工程鱼道设计导则》和《水电工程过鱼设施设计规范》建议鱼道流速不应超过鱼类最大游泳速度。鱼类通常使用爆发游泳速度通过鱼道的竖缝[32]。因此,建议鱼道竖缝流速控制在0.57~0.74 m/s (最小的Uburst~Uburst-EF90)。
4)出口流速。《水利水电工程鱼道设计导则》和《水电工程过鱼设施设计规范》建议鱼道出口水流应平稳并能引导鱼逆流而上,流速不宜大于0.50 m/s。因此建议鱼道出口断面至下一个池室之间的流速控制在0.21~0.50 m/s(Uind-EF90~0.50 m/s);选择有明显水流方向且环境流速在0.21~0.50 m/s的区域作为鱼道出口的选址。
3 讨 论
3.1 鱼类游泳能力与体长的相关关系
虽然一些文献利用直线函数拟合了鱼类游泳速度(临界游泳速度和爆发游泳速度)与体长的相关关系[25-26],但是对于大多数动物(包括鱼类)生理学方面相关关系的变化通常是异速的而不是等速的。随着鱼的表面积或体长平方的增大,鱼在水流中所受到的拖曳力成一定比例增大。运动肌肉的力量与肌肉体积成正比例变化,也与鱼体长的立方成正比例变化。肌肉质量与拖拽力之间的具体关系是复杂的,并随着鱼的形态和大小而变化。随着鱼尺寸的增大,鱼体表面积和肌肉质量增大,使得鱼类绝对游泳速度(m/s)增大,相对游泳速度(bl/s)减小,与本文图1临界游泳速度和爆发游泳速度结果相符。指数方程和基于数据对数变换的直线方程可被用于描述鱼类游泳速度和体长的相关关系[9,21,23,27-28]。根据上述结果与分析,数据经过对数变换的直线方程比未经过变换处理的直线方程的拟合效果更好,因此本研究结果可支撑说明鱼类游泳速度与体长的相关关系是异速变化的,而不是等速变化的。
鱼和许多水生动物一样,本能地逆着水流游动。鱼通过侧线感受器感知水流,并游动以对抗水流和保持稳定的位置[33-34]。鱼类聚集成群可以减少作用在各鱼体上的平均拖拽力[35]。因此,在给定的流速下,鱼群中的鱼比单独游动的鱼更容易抵抗水流拖拽力并保持稳定的位置。以上文作为间接的分析依据,对于特定的鱼种形态来说,随鱼体型的增大,鱼质量的增长速度比鱼表面积的增长速度更快,因此体型较大的鱼比体型较小的鱼在流水中更容易保持稳定的位置,即体型较大的鱼的绝对感应流速(m/s)随体长增大而增大,体型较大的鱼的相对感应流速(bl/s)随体长的增大而减小,与本文图1感应流速结果相符。
3.2 试验鱼游泳能力与其他文献的比较
本研究中5种鲤科鱼类(体长0.046~0.275 m)的感应流速为0.07~0.17 m/s(0.52~2.38 bl/s),与其他文献报道的鲤科鱼类(体长0.06~0.342 m)的感应流速大小类似(0.06~0.33 m/s,0.40~3.00 bl/s)[18,36-40]。由上述数据可知,已公开报道的鲤科鱼类(体长0.46~0.34 m)感应流速范围为0.06~0.33 m/s(0.40~3.00 bl/s),且大部分数据主要分布在0.1~0.2 m/s(1.0~2.0 bl/s)范围内。此次5种鲤科鱼类(体长0.046~0.275 m)的临界游泳速度为0.56~1.17 m/s(2.73~23.84 bl/s),爆发游泳速度为0.57~1.56 m/s(3.60~27.05 bl/s)。通过与其他17种鲤科鱼类(体长0.06~0.44 m)的临界游泳速度0.45~1.40 m/s(2.07~8.39 bl/s)和爆发游泳速度0.80~1.90 m/s (2.95~13.2 bl/s)[16,26]对比,本次测试表明这5种鲤科鱼类的绝对临界游泳速度(bl/s)和绝对爆发游泳速度(bl/s)小于其他17种鲤科鱼类,但绝对游泳速度(m/s)相反。究其原因,主要是由于本次测试研究的鱼类体长相对较小造成的可预期结果,即随着鱼类体长变小,其相对临界游泳速度(bl/s)和相对爆发游泳速度(bl/s)增大,绝对临界游泳速度(m/s)和绝对爆发游泳速度(m/s)减小[9,25-26]。
本研究中的日本鳗鲡(体长0.185~0.268 m)的爆发游泳速度0.57~0.95 m/s(2.63~3.88 bl/s),小于欧洲鳗鲡(体长0.18~0.47 m)的爆发游泳速度1.12~1.28 m/s(2.72~7.19 bl/s)[41]。本研究中的胡子鲇(体长0.239~0.303 m)的爆发游泳速度0.93~1.56 m/s (3.63~5.21 bl/s),大于澳洲鳗鲇和南方鲇(体长0.06~0.12 m)的爆发游泳速度0.41~0.74 m/s(3.57~7.96 bl/s)[42-43]。上述日本鳗鲡和胡子鲇及其近似鱼种的游泳能力大小的比较情况,与上文5种鲤科鱼类游泳能力比较情况类似,也均符合上文阐述的鱼类游泳能力与体长大小的相关关系。本研究中的斑鳢为鲈形目月鳢科鱼类,目前国内外暂时未见到其他月鳢科鱼类游泳能力的报道,因此本文将鲈形目其他科的鱼类与本研究斑鳢进行比较。本研究中的斑鳢(体长0.274~0.323 m)的爆发游泳速度0.75~1.31 m/s(2.58~4.05 bl/s)和临界游泳速度0.62~0.92 m/s (2.04~3.10 bl/s),均分别与其他4种鲈形目鱼类(体长0.18~0.39 m)的爆发游泳速度0.71~1.45 m/s(2.63~3.86 bl/s)和临界游泳速度0.45~0.94 m/s(2.45~4.41 bl/s)类似[44-46]。
本研究通过在封闭水槽中使用“递增流速法”测试了鱼类游泳能力,试验水槽的水流为简单流场,与天然河流中的复杂流场条件有一定差异。目前较多学者认为,由于鱼类可以在复杂流场中利用涡流减少耗能,因此使得封闭水槽中的测试会低估鱼类游泳能力[47-49];但也有学者报道,相对于封闭水槽来说,在相同流量条件下,由于水槽边壁效应和开放水槽水体表面波浪的影响,鱼类在开放水槽中游泳时会受到更大的水流阻力,因此鱼类在封闭水槽中的游泳能力比在开放水槽中的游泳能力更高[50]。因此,在封闭水槽和开放水槽中进行鱼类游泳能力测试所得到的结果都可能会和鱼类在野外以及真实鱼道中的游泳能力有一定差异。但就目前来说,水槽测试法仍然是鱼类游泳能力测试的国内外通行方法[18-23,51]。
虽然本文基于封闭水槽测试得到了鱼类游泳能力数据,并依据中国鱼道设计相关导则和规范以及国内外文献,对本研究目标工程的鱼道提供了流速设计建议,但为了能进一步提高鱼道流速设计水平,下一步应在更丰富的水力条件下进行鱼类研究。
4 结 论
随着试验鱼体长增大,相对感应流速、相对临界游泳速度和相对爆发游泳速度(体长/s)均减小,鱼类体长和游泳速度的相关关系可用直线方程表示,经过对数数据变换后的直线方程拟合关系的拟合效果比未经过对数变换的拟合效果更好。根据本研究鱼类测试结果,建议本工程鱼道进口诱鱼流速控制为0.35~0.47 m/s,池室流速控制为0.21~0.59 m/s,竖缝流速控制为0.57~0.74 m/s,出口断面至下一个池室之间的流速控制在0.21~0.50 m/s,选择有明显水流方向且环境流速在0.21~0.50 m/s的区域作为鱼道出口的选址。
致谢:感谢三峡大学金蕾同学对本文文字校对上提供的帮助。
[1] Barbarossa V, Schmitt R J P, Huijbregts M A J, et al. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide[J]. Proceedings of the National Academy of Sciences of the United States of America, 2020, 117(7): 3648-3655.
[2] Humborg C, Ittekkot V, Cociasu A, et al. Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure[J]. Nature, 1997, 386(6623): 385-388.
[3] Zhang P, Qiao Y, Schineider M, et al. Using a hierarchical model framework to assess climate change and hydropower operation impacts on the habitat of an imperiled fish in the Jinsha River, China[J]. Science of the Total Environment, 2019, 646: 1624-1638.
[4] Chen K Q, Tao J, Chang Z N, et al. Difficulties and prospects of fishways in China. An overview of the construction status and operation practice since 2000[J]. Ecological Engineering, 2014, 70: 82-91.
[5] Shi X, Kynard B, Liu D, et al. Development of fish passage in China[J]. Fisheries, 2015, 40(4): 161-169.
[6] Gardner C J, Rees-Jones J, Morris G, et al. The influence of sluice gate operation on the migratory behaviour of Atlantic salmon Salmo salar (L.) smolts[J]. Journal of Ecohydraulics, 2016, 1(1/2): 90-101.
[7] Katopodis C, Williams J G. The development of fish passage research in a historical context[J]. Ecological Engineering, 2012, 48: 8-18.
[8] Castro-Santos T, Shi X, Haro A. Migratory behavior of adult sea lamprey and cumulative passage performance through four fishways[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2016, 74(5): 790-800.
[9] Katopodis C, Cai L, Johnson D. Sturgeon survival: The role of swimming performance and fish passage research[J]. Fisheries Research, 2019, 212: 162-171.
[10] Silva A T, Lucas M C, Castro-Santos T, et al. The future of fish passage science, engineering, and practice[J]. Fish and Fisheries, 2018, 19(2): 340-362.
[11] 李广宁,孙双科,郄志红,等. 电站尾水渠内鱼道进口位置布局[J]. 农业工程学报,2019,35(24):81-89.
Li Guangning, Sun Shuangke, Qie Zhihong, et al. Arrangement of fishway entrance in power station tailrace channel[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2019, 35(24): 81-89. (in Chinese with English abstract)
[12] Peake S, Beamish F W, McKinley R S, et al. Relating swimming performance of lake sturgeon, Acipenser fulvescens, to fishway design[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(6): 1361-1366.
[13] Katopodis C. Developing a toolkit for fish passage, ecological flow management and fish habitat works[J]. Journal of Hydraulic Research, 2005, 43(5): 451-467.
[14] Baki A B M, Zhu D Z, Harwood A, et al. Rock-weir fishway II: design evaluation and considerations[J]. Journal of Ecohydraulics, 2017, 2(2): 142-152.
[15] Rodgers E M, Heaslip B M, Cramp R L, et al. Substrate roughening improves swimming performance in two small-bodies riverine fishes: Implications for culvert remediation and design[J]. Conservation Physiology, 2017, 5(1): cox034.
[16] Cai L, Katopodis C, Johnson D, et al. Case study: Targeting species and applying swimming performance data to fish lift design for the Huangdeng Dam on the upper Mekong River[J]. Ecological Engineering, 2018, 122: 32-38.
[17] Katopodis C, Gervais A R. Fish Swimming Performance Database and Analyses[M]. Ottawa: Fisheries and Oceans Canada. 2016.
[18] 蔡露,王伟营,王海龙,等. 鱼感应流速对体长的响应及在过鱼设施流速设计中的应用[J]. 农业工程学报,2018,34(2):176-181.
Cai Lu, Wang Weiying, Wang Hailong, et al. Response of induced flow speed to fish body length and its application in flow design of fish passage facilities[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2018, 34(2): 176-181. (in Chinese with English abstract)
[19] Brett J R. The respiratory metabolism and swimming performance of young Sockeye salmon[J]. Journal of the Fisheries Board of Canada, 1964, 21(5): 1183-1226.
[20] 付翔,付成,付世建. 五种淡水鱼类幼鱼游泳能力的比较[J]. 生态学杂志,2020,39(5):1629-1635.
Fu Xiang, Fu Cheng, Fu Shijian. Comparison of swimming ability among five freshwater fish species[J]. Chinese Journal of Ecology, 2020, 39(5): 1629-1635. (in Chinese with English abstract)
[21] Santos H A, Pompeu P S, Martinez C B. Swimming performance of the migratory neotropical fish(characiformes: anostomidae)[J]. Neotropical Ichthyology, 2007, 5(2): 139-146.
[22] Brett J R. Swimming performance of sockeye salmon () in relation to fatigue time and temperature[J]. Journal of the Fisheries Board of Canada, 1967, 24(8): 1731-1741.
[23] Santos H A, Viana E M F, Pompeu P S, et al. Optimal swim speeds by respirometer: An analysis of three neotropical species[J]. Neotropical Ichthyology, 2012, 10(4): 805-811.
[24] Castro-Santos T, Shi X, Haro A. Migratory behavior of adult sea lamprey and cumulative passage performance through four fishways[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2017, 74(5): 790-800.
[25] Hammer C. Fatigue and exercise tests with fish[J]. Comparative Biochemistry and Physiology Part A, 1995, 112(1): 1-20.
[26] Hou Y, Cai L,Wang X, et al. Swimming performance of 12 Schizothoracinae species from five rivers[J]. Journal of Fish Biology, 2018, 92(6): 2022-2028.
[27] Brett J R, Glass N R. Metabolic rates and critical swimming speeds of Sockeye salmon () in relation to size and temperature[J]. Journal of the Fisheries Board of Canada, 1973, 30(3): 379-387.
[28] Verhille C E, Poletto J B, Cocherell D E, et al. Larval green and white sturgeon swimming performance in relation to water-diversion flows[J]. Conservation Physiology, 2014, 2(1): cou031.
[29] 中华人民共和国水利部. 水利水电工程鱼道设计导则:SL609-2013[S]. 北京:中国水利水电出版社. 2013.
[30] 中华人民共和国能源局. 水电工程过鱼设施设计规范:NB/T35054-2015[S]. 北京:中国电力出版社. 2015.
[31] Pavlov D S. Structures assisting the migrations of non-salmonid fish: USSR[J]. FAO Fisheries Technical Paper, 1989, 308: 28-29.
[32] Blake R W. Fish Locomotion[M]. Cambridge: Cambridge University Press. 1983.
[33] Arnold G P. Rheotropism in fishes[J]. Biological Reviews, 1974, 49(4): 515-576.
[34] Montgomery J C, Macdonald J A. Sensory tuning of lateral line receptors in antarctic fish to the movements of planktonic prey[J]. Science, 1987, 235(4785): 195-197.
[35] Weihs D. Hydromechanics of fish schooling[J]. Nature, 1973, 241(5387): 290-291.
[36] 白艳勤,路波,石小涛,等. 草鱼、鲢和瓦氏黄颡鱼幼鱼感应流速的比较[J]. 生态学杂志,2013,32(8):2085-2089.
Bai Yanqin, Lu Bo, Shi Xiaotao, et al. Induction velocity of juvenile grass carp, silver carp, and darkbarbel catfish[J]. Chinese Journal of Ecology, 2013, 32(8): 2085-2089. (in Chinese with English abstract)
[37] 王博,石小涛,周琛琳,等. 北盘江两种鱼感应流速[J]. 北华大学学报:自然科学版,2013,14(2):223-226.
Wang Bo, Shi Xiaotao, Zhou Chenlin, et al. Induced velocity of two species of fishes in Beipan River[J]. Journal of Beihua University: Natural Science, 2013, 14(2): 223-226. (in Chinese with English abstract)
[38] 李志敏,陈明曦,金志军,等. 叶尔羌河厚唇裂腹鱼的游泳能力[J]. 生态学杂志,2018,37(6):1897-1902.
Li Zhimin, Chen Mingxi, Jin Zhijun, et al. Swimming ability ofDdy in Yarkand River[J]. Chinese Journal of Ecology, 2018, 37(6): 1897-1902. (in Chinese with English abstract)
[39] 丁少波,施家月,黄滨,等. 大渡河下游典型鱼类的游泳能力测试[J]. 水生态学杂志,2020,41(1):46-52.
Ding Shaobo, Shi Jiayue, Huang Bin, et al. Swimming capability of six typical fish species from the lower Dadu River. Journal of Hydroecology, 2020, 41(1): 46-52. (in Chinese with English abstract)
[40] 雷青松,涂志英,石迅雷,等. 应用于鱼道设计的新疆木扎提河斑重唇鱼的游泳能力测试[J]. 水产学报,2020,44(10):1718-1727.
Lei Qingsong, Tu Zhiying, Shi Xunlei, et al. Swimming ability ofin Muzati River related on fishway design[J]. Journal of Fisheries of China, 2020, 44(10): 1718-1727. (in Chinese with English abstract)
[41] Clough S, Lee-Elliott H, Turnpenny A, et al. Swimming speeds in fish: Phase 2[M]. Swindon: Environment Agency. 2003.
[42] Watson J R, Goodrich H R, Cramp R L, et al. Assessment of the effects of microPIT tags on the swimming performance of small-bodied and juvenile fish[J]. Fisheries Research, 2019, 218: 22-28.
[43] 曾令清,彭韩柳依,王健伟,等. 饥饿对南方鲇幼鱼游泳能力个体变异和重复性的影响[J]. 水生生物学报,2014,38(5):883-890.
Zeng Lingqing, Peng Hanliuyi, Wang Jianwei, et al. The effects of food deprivation on individual variation and repeatability in swimming performance of juvenile southern catfish (Chen)[J]. Acta Hydrobiologica Sinica, 2014, 38(5): 883-890. (in Chinese with English abstract)
[44] Dockery D R, McMahon T E, Kappenman K M, et al. Swimming performance of sauger (Sander canadensis) in relation to fish passage[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2017, 74(12): 2035-2044.
[45] Raffini F, Schneider R F, Franchini P, et al. Diving into divergence: differentiation in swimming performances, physiology and gene expression between locally-adapted sympatric cichlid fishes[J]. Molecular Ecology, 2020, 29(7): 1219-1234.
[46] Starrs T, Starrs D, Lintermans M, et al. Assessing upstream invasion risk in alien freshwater fishes based on intrinsic variations in swimming speed performance[J]. Ecology of Freshwater Fish, 2017, 26(1): 75-86.
[47] Mallen-Cooper M. Swimming ability of juvenile Australian Bass, Macquaria novemaculeata (Steindachner), and juvenile Barramundi, lates calcarifer (Bloch), in an experimental vertical-slot fishway[J]. Marine & Freshwater Research, 1992, 43(4): 823-833.
[48] Liao C J, Beal N D, Lauder V G, et al. Fish exploiting vortices decrease muscle activity[J]. Science, 2003, 302(5650): 1566-1569.
[49] Tudorache C, Viaenen P, Blust R, et al. Longer flumes increase critical swimming speeds by increasing burst-glide swimming duration in carp Cyprinus carpio, L[J]. Journal of Fish Biology, 2007, 71(6): 1630-1638.
[50] Shi X, Chen Q, Liu D, et al. Juvenile silver carp Hypophthalmichthys molitrix swim faster in closed surface flow than open surface flow[J]. Environmental Biology of Fishes, 2014, 97(12): 1411-1416.
[51] Zhao Z, Liang R, Wang Y, et al. Study on the swimming ability of endemic fish in the lower reaches of the Yangtze River: A case study. Global Ecology and Conservation, 2020, 22: e01014.
Response of fish swimming ability to body length and its application in fishway design
Cai Lu1,2, Hou Yiqun1, Jin Yao1, Yang Zhi1, Hu Wangbin1, Chen Xiaojuan1, Chen Jihua2, Huang Yingping2, Han Deju1※
(1.,,430079,;2.443002,)
Water conservancy and hydropower projects have posed a great threat to the fish habitat environment and migration, even the gene exchange between fish populations, where the river connectivity is restructured via manmade canals. Fishways are an increasingly popular solution to facilitate the passage of fish migrating. However, the flow speed of fishway depends mainly on fish swimming ability. Consequently, the body length of fish is a key factor in fish swimming ability. In this study, a series of stepped velocity tests were carried out to evaluate the induced flow speed (Uind), critical swimming speed (Ucrit), and burst speed (Uburst) of target species using the modified Brett-type swimming respirometer. A new fishway was also developed in Hainan Province of China to explore the effect of fish body length on swimming ability and flow speed. An Origin 9.0 software was used to transform three fish speeds and body lengths for linear regressions. Kaplan-Meier curves (Survival curves) were plotted using cumulative percentages and speeds in fish swimming ability tests. The suggestions were also given on flow speeds for fishway design. The results indicated that the relatively induced flow speed, critical swimming speed, and burst speed (bl/s) decreased with body length increasing. Linear relationships were achieved to describe the fish speeds and body lengths. The fitting efficiency of linear equation (2and) after natural logarithmic data transformation was better than that without transformation, and the2values increased from 0.664-0.725 to 0.907-0.954. The effective speed for 90% of tested fish (EF90) was set as the flow speed in the vertical slot fishway. The EF90 value for Uind(90% induction rate, Uind-EF90) was 0.21 m/s, while the values for Ucritand Uburst(90% success rates, Ucrit-EF90and Uburst-EF90) were 0.59 m/s and 0.74 m/s, respectively. According to the standard design of vertical slot fishway realized by the, andin China, the flows of a fishway were recommended as follows: 1) Entrance. The flush water should be provided if the flow speed of the entrance was lower than fish-induced flow speed. The recommended entrance speed was 0.35-0.47 m/s (0.6-0.8 Ucrit-EF90) if the Ucirt-EF90was 0.59 m/s, where the critical swimming speed of 0.6-0.8 m/s was generally set for attracting fish to the entrance. (2) Pool. The recommended pool speed was 0.21-0.59 m/s (Uind-EF90-Ucrit-EF90). The reason was that the pool speed was lower than burst speed, where the fish cannot lose their orientation. Since the fish critical swimming speed was the maximum sustainable swimming speed, the fish can commonly swim several hundred meters of fishway (100-500 pools), where the fishway design considered fish swimming endurance. 3) Vertical slot. The recommended velocity of the vertical slot was 0.57-0.74 m/s (minimum Uburst-Uburst-EF90). The reason was that the flow speed of fishway should lower the maximum swimming speed of fish, where the fish commonly used burst swimming to pass vertical slots. 4) Exit. The recommended exit speed was 0.21-0.50 m/s (Uind-EF90-0.50 m/s). The exit flow should be smooth and guide fish upstream, exceeding the induced flow speed of <0.50 m/s. These demonstrated that the fish speed decreased relatively, as the body lengths increased using natural logarithmic function in flow speed of fishway design.
flow velocity,body length, swimming ability, induced flow speed, critical swimming speed, burst speed, vertical slot fishway
蔡露,侯轶群,金瑶,等. 鱼游泳能力对体长的响应及其在鱼道设计中的应用[J]. 农业工程学报,2021,37(5):209-215.doi:10.11975/j.issn.1002-6819.2021.05.024 http://www.tcsae.org
Cai Lu, Hou Yiqun, Jin Yao, et al. Response of fish swimming ability to body length and its application in fishway design[J]. Transactions of the Chinese Society of Agricultural Engineering (Transactions of the CSAE), 2021, 37(5): 209-215. (in Chinese with English abstract) doi:10.11975/j.issn.1002-6819.2021.05.024 http://www.tcsae.org
2020-07-15
2021-01-21
国家自然科学基金(51609155,51709187);水利部水工程生态效应与生态修复重点实验室开放课题基金(野生及人工养殖鱼类的游泳能力差异研究项目);三峡大学三峡库区生态环境教育部工程研究中心开放基金(KF2019-09)
蔡露,助理研究员,研究方向为鱼类行为和过鱼设施。Email:tyzhcl@hotmail.com
韩德举,研究员,研究方向为鱼类行为和过鱼设施。Email:handeju@163.com
10.11975/j.issn.1002-6819.2021.05.024
Q958; TV61
A
1002-6819(2021)-05-0209-07
猜你喜欢
天天爱科学(2022年9期)2022-09-15 01:12:28
科普童话·百科探秘(2022年8期)2022-05-30 10:48:04
农业工程学报(2022年4期)2022-04-24 03:45:42
昆明医科大学学报(2022年1期)2022-02-28 07:46:32
河北农业大学学报(2020年6期)2021-01-08 03:48:08
中国水能及电气化(2020年8期)2020-09-29 00:46:12
卷宗(2020年16期)2020-08-10 18:21:27
水利水电科技进展(2019年5期)2019-11-01 00:53:12
小哥白尼(野生动物)(2019年6期)2019-10-10 06:27:22
水利科技与经济(2018年5期)2018-03-29 21:14:44
