
ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株上烟粉虱寄主适应性及寄主植物营养成分含量和防御反应变化
2021-05-11 08:33丁天波
昆虫学报 2021年3期
丁天波, 周 雪, 杨 楠, 杨 炀, 唐 瑶, 褚 栋
(青岛农业大学植物医学学院, 山东省植物病虫害综合防控重点实验室, 山东青岛 266109)
烟粉虱Bemisiatabaci是一种至少包含44个隐种的世界性农业害虫(Kanakala and Ghanim, 2019)。除了通过直接刺吸植物汁液和分泌蜜露导致煤污病等方式危害植物生长以外,该虫作为媒介昆虫还可以传播至少111种植物病毒(Jones, 2003),给多种蔬菜和经济作物生产造成严重的经济损失。例如,烟粉虱传播的番茄黄化曲叶病毒(tomato yellow leaf curl virus, TYLCV),自2006年在我国上海首次被报道后,短时间内蔓延至多地,损失惨重(Wuetal., 2006)。近年来,烟粉虱等粉虱传播的另一种植物病毒——番茄褪绿病毒(tomato chlorosis virus, ToCV),自2012年在我国内陆地区首次被发现(Zhaoetal., 2013)后,迅速蔓延至山东、河北、河南、海南等16个省份和地区(Tangetal., 2017; 魏可可等, 2018; 刘勇等, 2019)。研究发现ToCV在我国的快速流行同烟粉虱种群的扩散有着密不可分的联系(刘永光等, 2014; 代惠洁等, 2016)。目前,山东、江苏等多个地区均报道TYLCV和ToCV复合侵染(TYLCV&ToCV复合侵染)发生,给当地番茄种植带来毁灭性伤害(赵黎明等, 2014; 吴淑华等, 2016; 陈利达等, 2020)。ToCV单独侵染以及TYLCV&ToCV复合侵染与烟粉虱的互作研究,对于深入理解植物病毒的扩散流行机制具有重要指导价值(Dingetal., 2019)。目前,对于烟粉虱与这些病毒的互作机制知之甚少。
植物病毒可以对媒介昆虫的寄主适应性产生不同的影响,对其自身的传播具有重要的意义。Belliure等(2005)发现番茄斑萎病毒(tomato spotted wilt virus, TSWV)有利于其媒介昆虫西花蓟马Frankliniellaoccidentalis的存活和生长发育;同健康烟草相比,烟草花叶病毒(tobacco mosaic virus, TMV)侵染的烟草对烟蚜Myzuspersicae的种群增长则具有明显的抑制作用(王佳等, 2017)。诸多研究表明,植物病毒能够改变寄主植物的营养条件和防御反应,并且是媒介昆虫寄主适应性变化的重要原因。木薯花叶病毒(cassava mosaic virus, CMV)侵染能够显著提高木薯植株韧皮部汁液中氨基酸的含量,其媒介昆虫烟粉虱的种群增长速率亦明显加快(Colvinetal., 2006);而西葫芦感染黄瓜花叶病毒(cucumber mosaic virus, CMV)后,叶片和韧皮部的糖分和氨基酸比例均显著下降,从而导致蚜虫的种群适合度降低(Maucketal., 2010, 2014);中国番茄黄化曲叶病毒(tomato yellow leaf curl China virus, TYLCCNV)和其卫星的复合侵染能够显著抑制寄主植物的茉莉酸(jasmonic acid, JA)防御途径,从而对烟粉虱的寄主适应性产生有利影响(Zhangetal., 2012)。
媒介昆虫是虫媒植物病毒流行扩散的重要“桥梁”,开展植物病毒对媒介昆虫的影响及机制研究显得尤为必要。本研究从田间番茄病毒病发生现状出发,重点围绕ToCV单独侵染和TYLCV&ToCV复合侵染对烟粉虱MED隐种的寄主适应性影响及生理机制展开研究,结合存活率和产卵量两个参数解析不同番茄病毒侵染对烟粉虱MED隐种的寄主适应性影响;通过测定番茄植株氨基酸和总糖含量比较分析了ToCV单独侵染和TYLCV&ToCV复合侵染对寄主植物营养状况的影响;最后,综合茉莉酸和水杨酸(salicylic acid, SA)途径关键基因表达模式解析不同番茄病毒侵染对寄主植物防御的影响。研究结果将从植物病毒单独侵染和复合侵染两个角度进一步理解烟粉虱与病毒的互作机制。
1 材料与方法
1.1 供试昆虫
供试烟粉虱MED隐种于2012年采自山东省济南市,并长期以棉花Gossypiumhirsutum(品种:鲁棉28)为寄主在人工气候室中连续饲养。其中饲养温度为27±1℃,相对湿度为60%±5%,光周期为16L∶8D。
1.2 供试病毒来源
供试ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株采自于山东省青岛市,通过室内饲养烟粉虱对番茄Solanumlycopersicum(品种:中杂9号)植株进行病毒接种,成功后置于人工气候室中培养,作为供试病毒来源长期保存。ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株通过以下步骤获得:将25头烟粉虱MED隐种雄成虫置于ToCV单独侵染或TYLCV&ToCV复合侵染番茄植株上取食获毒48 h,将其取出,放入微虫笼中并置于番茄苗(4-5真叶期)从下往上数第2片真叶上,待传毒48 h后,移出所有烟粉虱,将已接种番茄苗置于网笼中隔离培养,1个月后,根据发病症状并结合RT-PCR病毒检测结果判断接种番茄的携毒情况,接种成功的番茄植株作为后续试验用病毒植株;对照健康番茄植株获取方式同上,即用烟粉虱MED隐种雄成虫取食健康番茄48 h后,再置于健康番茄苗相同叶片上48 h。
1.3 烟粉虱MED隐种成虫在不同番茄植株上的存活率和产卵量比较
参照Su等(2015)的方法,分别取20头健康初羽化烟粉虱MED隐种雌成虫放入微虫笼中,并置于1.2节制备的ToCV单独侵染、TYLCV&ToCV复合侵染和健康番茄植株从下往上数第4片真叶上,待7 d后,统计在不同番茄植株叶片上烟粉虱的死亡数,计算其存活率,并统计烟粉虱7 d的产卵量。每种处理保证至少3个生物学重复。
1.4 番茄植株中氨基酸含量测定
分别剪取1.2节ToCV单独侵染、TYLCV&ToCV复合侵染和健康番茄植株新鲜叶片,用液氮进行研磨,称取0.5 g研磨组织,置于水解管中,并加入16 mL HCl溶液(6 mol/L),真空脱气30 min后,充氮封管,110℃下水解24 h后,取出冷却,使用ddH2O转移并定容至50 mL。准确吸取1 mL水解液,真空脱酸抽干,加入1 mL ddH2O抽干(重复2次),并加入1 mL HCl溶液(0.02 mol/L)充分溶解后,用0.22 μm滤膜进行过滤,将滤液置于高速氨基酸分析仪(日立8900)上进行水解氨基酸和总氨基酸含量测定。每种处理至少保证3个生物学重复。
1.5 番茄植株中总糖含量测定
不同番茄植株中可溶性总糖含量测定参照蒽酮比色法(陈毓荃, 2002; 丁剑等, 2017),并稍作改进。 分别称取1.2节ToCV单独侵染、TYLCV&ToCV复合侵染和健康番茄植株新鲜叶片1.0 g,放入三角瓶中,加入25 mL ddH2O后,在沸水浴中煮沸20 min,待自然冷却后,使用滤膜过滤并用ddH2O定容至100 mL即得样品提取液。将500 μL样品提取液、500 μL ddH2O和4 mL蒽酮试剂混匀,在沸水浴中煮沸10 min,待自然冷却后,取250 μL样品混合液在多功能酶标仪(SpectraMax® Plus, Molecular Devices)上620 nm处记录吸光值,对照孔以ddH2O代替样品提取液。最终结合葡萄糖标准曲线,计算不同番茄样品的可溶性总糖含量,每种处理至少保证3个生物学重复。
1.6 番茄植株JA和SA信号通路关键基因表达模式分析
分别剪取1.2节ToCV单独侵染、TYLCV&ToCV复合侵染和健康番茄植株从下往上数第4片真叶,经过液氮研磨后,使用TRIzol(Thermo Fisher Scientific)完成总RNA的提取,并对所提取总RNA的质量和浓度进行测定。不同样品均选取1 μg总RNA,利用PrimeScript RT Reagent Kit (Perfect Real-time)(TaKaRa)并根据说明书完成cDNA第1链的合成。每种处理至少保证3个生物学重复。
番茄JA信号通路关键基因(FAD7和PIII)及SA信号通路关键基因(NPR1和PR1)的实时荧光定量引物参照Li等(2017),根据前人研究选取ubiquitin3(UBI)基因作为内参(Shietal., 2013),具体引物信息见表1。 RT-qPCR反应体系: SYBR Premix ExTaqTMII(TaKaRa)10 μL, cDNA 1 μL, 上下游引物(10 μmol/L)各1 μL, ddH2O 7 μL。反应条件: 95℃ 30 s; 95℃ 5 s, 60℃ 30 s, 40个循环。结合目的基因和内参基因的Ct值,每个基因的相对表达量通过2-△△Ct法计算(Livak and Schmittgen, 2001),每个处理至少保证3个生物学重复和2个技术重复。

表1 实时荧光定量PCR所用引物信息Table 1 Primer sequences used for RT-qPCR
1.7 数据分析
所有实验数据以平均值±标准误表示,运用SPSS 16.0软件对数据进行单因素方差分析(one-way ANOVA),采用Tukey氏检验对不同处理数据之间的差异显著性进行分析(P<0.05)。
2 结果
2.1 烟粉虱MED隐种成虫在病毒侵染和健康番茄植株上的存活率和产卵量比较
烟粉虱MED隐种以病毒侵染和健康番茄作为寄主时,其存活率之间存在显著性差异(P<0.05)(图1: A)。当烟粉虱MED隐种成虫以健康番茄植株为寄主时,7 d后其存活率为62%,分别是以ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株为寄主时的1.82和1.88倍(P<0.05)(图1: A)。当烟粉虱MED隐种成虫以TYLCV&ToCV复合侵染番茄植株为寄主时,其存活率略低于以ToCV单独侵染番茄植株为寄主时,但差异不显著(P>0.05)(图1: A)。

图1 不同番茄病毒侵染7 d的番茄植株上的烟粉虱MED隐种成虫存活率(A)和产卵量(B)Fig. 1 Survival rate (A) and number of eggs laid (B) of Bemisia tabaci MED adultson tomato plants infected with different tomato viruses for 7 dHealthy: 健康番茄植株Healthy tomato plants; ToCV: ToCV单独侵染番茄植株ToCV-singly infected tomato plants; TYLCV&ToCV: TYLCV&ToCV复合侵染的番茄植株TYLCV&ToCV co-infected tomato plants. 图中数据为平均值±标准误;柱上不同字母表示经Tukey氏检验后差异显著(P<0.05)。Data in the figure are means±SE, and different letters above the bars indicate significant differences by Tukey’s test (P<0.05). 下图同The same for the following figures.
烟粉虱MED隐种成虫在不同病毒侵染和健康番茄上的产卵量如图1(B)所示,在ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株上7 d的产卵量分别为261和226粒,均显著低于在健康番茄上的产卵量(389.3粒)(P<0.05)。烟粉虱MED隐种成虫在ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株上的产卵量差异并不显著(P>0.05)。
2.2 病毒侵染对番茄植株氨基酸含量的影响
不同病毒侵染和健康番茄植株体内总氨基酸含量检测结果表明(图2: A),经TYLCV&ToCV复合侵染后,番茄植株中总氨基酸含量明显低于健康番茄植株中总氨基酸含量,并且显著低于ToCV单独侵染后番茄植株中的总氨基酸含量(P<0.05)。ToCV单独侵染和健康番茄植株中的总氨基酸含量则无显著性差异(P>0.05)。
进一步对不同病毒侵染和健康番茄植株中17种水解氨基酸含量进行测定和比较分析,结果发现(图2: B),在ToCV单独侵染、TYLCV&ToCV复合侵染和健康番茄植株中,谷氨酸(Glu)、天冬氨酸(Asp)和亮氨酸(Leu)含量均较高,半胱氨酸(Cys)、蛋氨酸(Met)和组氨酸(His)含量均较低。TYLCV&ToCV复合侵染番茄植株中丝氨酸(Ser)、甘氨酸(Gly)、丙氨酸(Ala)、缬氨酸(Val)、亮氨酸(Leu)和苯丙氨酸(Phe)的比例显著降低(P<0.05),14种氨基酸[天冬氨酸(Asp)、苏氨酸(Thr)、丝氨酸(Ser)、甘氨酸(Gly)、丙氨酸(Ala)、缬氨酸(Val)、异亮氨酸(Ile)、亮氨酸(Leu)、酪氨酸(Tyr)、苯丙氨酸(Phe)、赖氨酸(Lys)、组氨酸(His)、精氨酸(Arg)、脯氨酸(Pro)]含量均显著低于ToCV单独侵染番茄植株(P<0.05);其他3种氨基酸[谷氨酸(Glu)、半胱氨酸(Cys)、蛋氨酸(Met)]含量在ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株中则没有显著性差异(P>0.05)。

图2 不同番茄病毒侵染对番茄植株中总氨基酸(A)和水解氨基酸(B)含量的影响Fig. 2 Effects of the infection of different tomato viruses on the contents of total amino acids (A)and hydrolyzed amino acids (B) in tomato plants
2.3 病毒侵染对番茄植株总糖含量的影响
通过对不同病毒侵染和健康番茄植株内总糖含量进行测定和比较,结果表明(图3),TYLCV&ToCV复合侵染番茄植株中总糖含量低于健康番茄植株和ToCV单独侵染番茄植株总糖含量,并且同ToCV单独侵染番茄植株之间差异显著(P<0.05)。
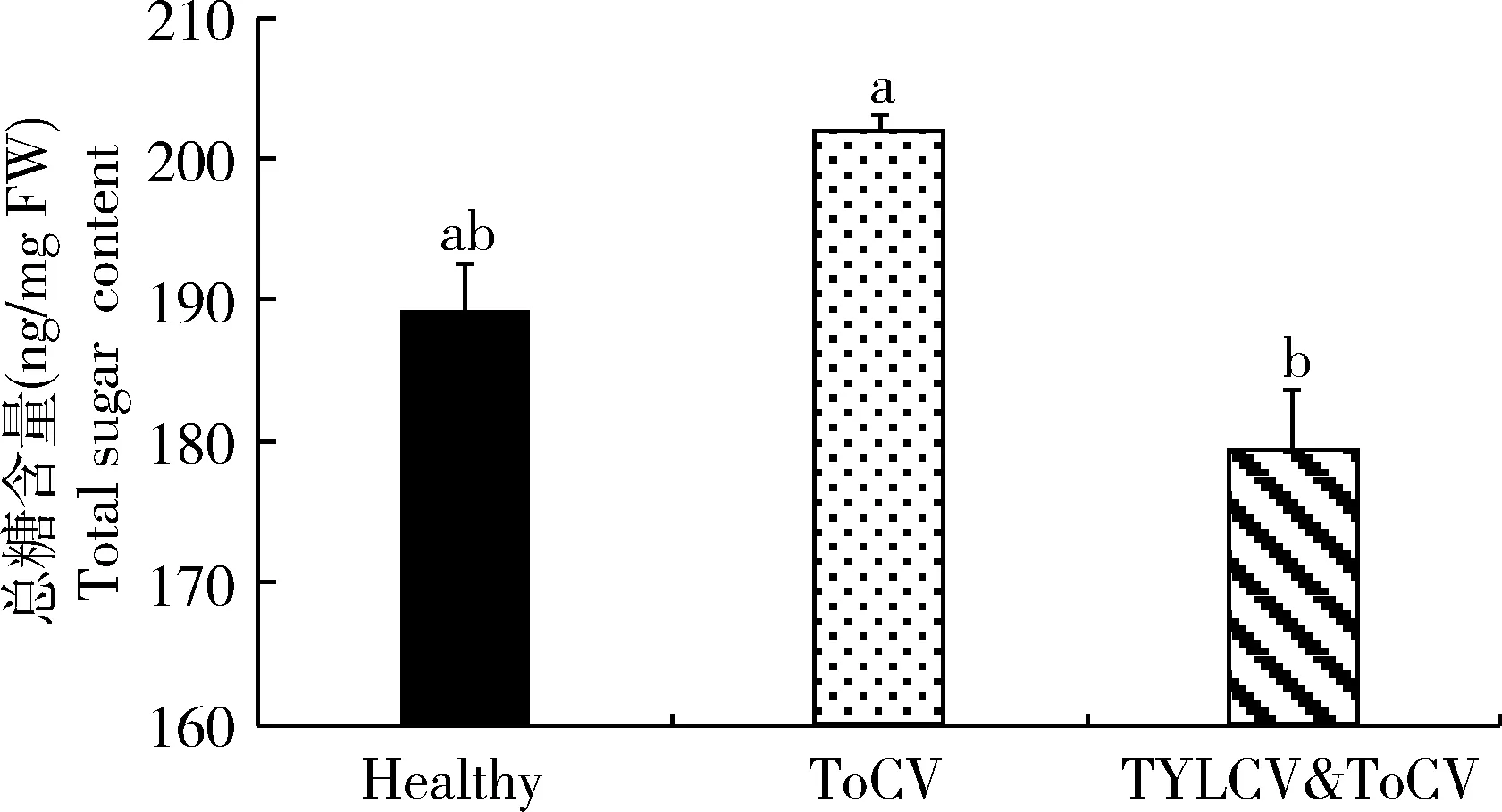
图3 不同番茄病毒侵染对番茄植株中总糖含量的影响Fig. 3 Effects of the infection of different tomato viruses onthe total sugar content in tomato plants
2.4 病毒侵染条件下番茄植株JA信号通路关键基因响应模式比较
本研究选取番茄JA信号通路两个关键基因FAD7和PIII,对二者在不同病毒侵染和健康番茄中的表达模式进行比较研究。相对于健康植株,ToCV单独侵染和TYLCV&ToCV复合侵番茄植株中FAD7基因的mRNA表达水平均显著降低(P<0.05),分别是健康番茄植株中的0.86和0.46倍(图4: A)。TYLCV&ToCV复合侵染番茄植株中FAD7基因的表达量显著低于ToCV单独侵染番茄植株中该基因的表达量(P<0.05),前者是后者的0.53倍。同样,在ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株中PIII基因的表达水平显著低于健康番茄植株中的(P<0.05),分别是健康番茄植株中表达量的0.31和0.05倍(图4: B)。并且TYLCV&ToCV复合侵染番茄植株中PIII基因的表达量最低,是ToCV单独侵染番茄植株中的0.16倍,差异显著(P<0.05)。

图4 不同番茄病毒侵染对番茄植株中JA信号通路关键基因FAD7(A)和PI II(B)表达模式的影响Fig. 4 Effects of the infection of different tomato viruses on the expression profiles of the key genes FAD7 (A)and PI II (B) involved in JA signal pathway in tomato plants
2.5 病毒侵染条件下番茄植株SA信号通路关键基因响应模式比较
本研究选取番茄SA信号通路两个关键基因NPR1和PR1,对二者在不同病毒侵染和健康番茄中的表达模式进行比较研究。结果发现,ToCV单独侵染和TYLCV&ToCV复合侵染后番茄植株中NPR1基因的mRNA表达水平均有上升趋势,其中TYLCV&ToCV复合侵染番茄植株中NPR1基因表达量显著高于健康和ToCV单独侵染番茄植株中的(P<0.05)(图5: A)。ToCV单独侵染和TYLCV&ToCV复合侵染番茄植株中PR1基因表达水平均高于健康番茄植株中的,但被ToCV单独侵染后,番茄植株中PR1基因表达水平显著高于TYLCV&ToCV复合侵染番茄植株中的(P<0.05),且前者是后者的2.78倍(图5: B)。

图5 不同番茄病毒侵染对番茄植株SA途径关键基因NPR1 (A)和PR1 (B)表达模式的影响Fig. 5 Effects of the infection of different viruses on the expression profiles of the keygenes NPR1 (A) and PR1 (B) involved in SA signal pathway in tomato plants
3 讨论
本研究发现,番茄植株经ToCV单独侵染后,烟粉虱MED隐种成虫在其上的存活率和产卵量均显著低于在健康番茄植株上的(图1),与Li等(2018)结合两性生命表研究的研究结果是一致的,该研究以ToCV单独侵染番茄植株作为寄主的烟粉虱MED隐种成虫的繁殖力、寿命等均显著降低。本研究还发现取食TYLCV&ToCV复合侵染番茄植株的烟粉虱MED隐种成虫的存活率和产卵量也明显降低,说明不论是ToCV单独侵染,还是TYLCV&ToCV复合侵染,均能够对其媒介昆虫烟粉虱的寄主适应性带来不利的影响。前人研究发现相对于ToCV侵染番茄植株,携带ToCV烟粉虱MED隐种则更倾向于选择健康番茄植株(Shietal., 2018; Weietal., 2019);本研究发现ToCV侵染植株不利于烟粉虱生存,可能与携毒烟粉虱寄主选择性相关。
植物病毒能够引起植物营养状况(氨基酸、糖分含量等)的改变,从而影响媒介昆虫的寄主适应性(Bluaetal., 1994; Colvinetal., 2006)。例如取食TYLCV侵染植株的烟粉虱体重、产卵量和存活率均显著升高,可能与TYLCV侵染番茄韧皮部汁液中氨基酸和糖分含量升高相关(Suetal., 2015)。二斑叶螨Tetranychusurticae对TSWV侵染番茄植株具有明显的寄主偏好性,并且其产卵量显著升高,可能与TSWV侵染番茄植株中氨基酸含量升高相关(Nachappaetal., 2013)。而本研究结果显示,TYLCV&ToCV复合侵染番茄植株总氨基酸含量明显低于健康番茄植株,其中3种昆虫必需氨基酸[缬氨酸(Val)、亮氨酸(Leu)和苯丙氨酸(Phe)]的比例显著降低(图2: B);TYLCV&ToCV复合侵染番茄植株总氨基酸含量和14种水解氨基酸的比例均显著低于ToCV单独侵染植株(图2)。此外,TYLCV&ToCV复合侵染番茄植株中总糖含量低于ToCV单独侵染和健康植株(图3)。这些结果也表明TYLCV&ToCV复合侵染比ToCV单独侵染对番茄植株营养状况更具不利影响。
植物病毒亦能够影响寄主植物体内的防御响应,导致昆虫寄主适应性的改变(Zhangetal., 2012)。为进一步探究媒介昆虫烟粉虱在ToCV单独侵染或TYLCV&ToCV复合侵染番茄植株上生物学特性的差异,本研究中运用RT-qPCR技术对不同病毒侵染条件下番茄JA和SA信号通路关键基因的响应模式进行了分析。结果显示,ToCV单独侵染和TYLCV&ToCV复合侵染后,番茄JA信号通路两个关键基因FAD7和PIII的表达量均显著下调,并且其在复合侵染番茄植株体内表达水平最低(图4)。前人在病毒侵染植株上也发现了JA途径被抑制的现象(Lewseyetal., 2010; Shietal., 2014),本研究同时也发现番茄SA信号通路两个关键基因NPR1和PR1则在一定程度上被不同番茄病毒诱导表达,NPR1基因在TYLCV&ToCV复合侵染条件下上调幅度最大,而PR1基因的表达水平则在ToCV单独侵染番茄中最高(图5)。同样,前人在烟草花叶病毒(tobacco mosaic virus, TMV)或TYLCV侵染番茄中也发现SA含量上升的现象(Thaleretal., 2010; Shietal., 2013),烟粉虱MEAM1隐种取食SA处理番茄植株后,其存活率和产卵量均显著降低。因此,可以推断本研究中ToCV单独侵染和TYLCV&ToCV复合侵染诱导的SA信号通路正响应在一定程度上降低了烟粉虱MED隐种的寄主适应性。
前人研究发现,TYLCV侵染有利于烟粉虱MED隐种种群上升,可能由于TYLCV侵染可使植物营养条件朝有利于烟粉虱生存的方向发展,并且TYLCV侵染显著抑制了植物防御反应,诸如植物过氧化物酶和超氧化物歧化酶等的合成(Suetal., 2015)。而在本研究中,TYLCV&ToCV的复合侵染植株则不利于烟粉虱MED隐种种群上升,植株营养明显下降,这可能是由于TYLCV&ToCV复合侵染与植物的互作关系不同于病毒单独侵染与植物的互作关系。该方面的机制尚需进一步的研究。
猜你喜欢
当代水产(2022年1期)2022-04-26
中国农业科学(2021年23期)2022-01-14
建材发展导向(2021年11期)2021-07-28
中国瓜菜(2020年8期)2020-09-26
祝您健康(2020年5期)2020-05-14
当代水产(2020年10期)2020-03-17
中国瓜菜(2019年8期)2019-09-19
植物保护(2018年5期)2018-12-05
湖北农业科学(2017年20期)2017-11-17
延安大学学报(自然科学版)(2015年4期)2015-06-15
