
陕西地区烟粉虱生物型的鉴定及其危害分析
2015-06-15 18:56赵瑞华邓振山贺晓龙
延安大学学报(自然科学版) 2015年4期
赵瑞华,邓振山,贺晓龙
(延安大学生命科学学院,陕西延安716000)
陕西地区烟粉虱生物型的鉴定及其危害分析
赵瑞华,邓振山,贺晓龙
(延安大学生命科学学院,陕西延安716000)
为探明陕西地区烟粉虱的危害及主要生物型,实地调查了陕西地区主要农作物上烟粉虱的危害状况与发生程度,并利用mtDNACOI基因测序方法,对陕西地区烟粉虱的生物型组成进行了分析鉴定。同源性分析结果表明,B型和Q型烟粉虱是陕西地区烟粉虱种群的主要生物型,其中B型占所测总样本的78.85%,表明陕西省烟粉虱主要生物型仍是B型,但Q型种群比例在增长,不排除有取代B型成为陕西地区烟粉虱主要危害类型的趋势。在陕西地区烟粉虱主要危害的植物是茄科的烟草、甜椒、番茄以及葫芦科的黄瓜、西葫芦,而十字花科植物普遍受害较轻,茄科的马铃薯和豆科的豇豆、豌豆受害最轻。
烟粉虱;生物型;危害;寄主植物
烟粉虱Bemisiatabaci(Gennadius)是一种世界性的重要害虫,食性广泛,寄主植物多样。其主要危害蔬菜、棉花、观赏植物等,并且能够传播110多种植物病毒,给许多经济作物造成严重的经济损失[1]。烟粉虱是一个正处于快速进化过程的复合种,由多种不同的生物型构成,到2005年为止被赋予特定生物型的种群有26个[2]。不同生物型的烟粉虱在许多方面(如地理分布、寄主范围、传毒能力及对农药敏感性等)存在显著差异。
由于二十世纪末烟粉虱在我国开始爆发,并且引发的危害越来越严重,我国相关单位加大了对烟粉虱的研究力度。首先对烟粉虱的发生危害及生物学特性进行了研究,在此基础上,也对我国烟粉虱的生物型在分子水平上进行了初步研究鉴定。如利用RAPD-PCR分析研究,将我国10个烟粉虱种群分为3种生物型(包括B型烟粉虱),通过rDNA-ITS1序列分析,证明B型烟粉虱是我国的入侵生物型[3]。利用mtCOI(cytochrome oxidaseI)基因片段作标记,采用序列分析法对近年来在我国爆发危害的5个烟粉虱种群的生物型进行鉴定,结果表明5个种群生物型与Texas-B型和Arizona-B型种群为同一生物型“B”[4]。应用mtCOI分子标记证实我国绝大部分地区烟粉虱为B型,云南、北京地区存在Q型,浙江、广东存在非B/Q型烟粉虱[5]。2005年首次报道了于2003年在云南昆明采集的烟粉虱为Q型烟粉虱[6],随后在浙江局部地区、江苏省的南通和盐城、北京、河南省也发现了Q型烟粉虱的存在[7-9]。2003年7月在浙江大学华家池校区的蔬菜上采集到一个烟粉虱种群,经mtDNACOI基因序列分析为非B型烟粉虱,初步认定该种群为中国本地的一种非B生物型,暂时称为浙江非B型烟粉虱China-ZHJ-1(简称ZHJ-1)种群。mtCOI分子标记鉴定结果显示,我国至少存在6个烟粉虱单倍型(B、Q、ZHJ-1、ZHJ-2、ZHJ-3和FJ-1)[10]。
近年来,烟粉虱在陕西省迅速由次要害虫上升为主要害虫,其危害逐渐引起人们的重视,但还没有关于烟粉虱在陕西地区分布危害及生物型鉴定的比较全面的报道。笔者曾于2009~2011年对全国19个省市5个科的寄主植物共58个烟粉虱地理种群进行过较为系统的研究,发现Q型烟粉虱发生频率逐年升高,但当时陕西地区只检测到B型烟粉虱[11]。因此,为了探讨不同生物型烟粉虱种群在陕西省不同地区的发生现状,预测其发展趋势,明确陕西省三大生态区域主要受害作物上烟粉虱的生物型以及不同生物型烟粉虱的地理分布与主要寄主上发生程度之间的关系。我们于2013年烟粉虱发生危害高峰期在陕西省9个市、20个地点对当地主要受害寄主上烟粉虱的发生危害情况进行了抽样调查,应用危害指数对不同作物上烟粉虱的发生危害程度进行了比较,同时通过mtDNACOI分子标记技术鉴定各地主要受害寄主上烟粉虱生物型,并在此基础上构建系统进化树,以期从宏观上进一步把握不同生物型烟粉虱在陕西省的区域性遗传分布与主要寄主种类,从而为该地区烟粉虱的综合治理指导防治提供合理的建议。
1 材料与方法
1.1 采样及危害程度调查
2013年6月到9月在烟粉虱发生高峰期,分别在陕西省6个地区陕北延安、陕北榆林、关中宝鸡、关中西安、陕南安康和陕南商洛调查烟粉虱寄主植物。调查对象主要是大田作物如蔬菜、烟草等。调查时轻轻翻转叶片,收集成虫,用75%乙醇浸泡于-20℃保存备用。记录寄主植物,并确定该作物所属的科、属、种。
对主要寄主植物上烟粉虱虫口密度与危害情况的调查采用5点取样法。每个点随机选取10个植株,每个植株再分别选上、中、下三个部位的叶片各3片,把叶片轻轻翻转过来,观察叶片的背面,记录烟粉虱成虫数。同时采集叶片带回实验室,在解剖镜下观察统计烟粉虱卵、若虫和伪蛹的数量。有些寄主植物叶片比较密集难以区分上、中、下三部分,则分别选取靠近植株中下部的老叶和上部的嫩叶各3片进行镜检,以叶片为单位统计寄主植物上烟粉虱若虫和伪蛹的数量,求取平均值作为最终结果。目前按照国内外有关烟粉虱危害程度的标准,危害等级划分为4级:1级“+”,代表每个叶片小于10头;2级“++”,代表每个叶片10~30头;3级“+++”,代表每个叶片30~50头;4级“++++”,代表每个叶片大于50头。同时寄主上发生危害程度以发生最严重时的样本记录危害等级。
1.2 烟粉虱生物型的鉴定与分析
1.2.1 单头烟粉虱总DNA的提取
从每个采集地点的烟粉虱样本中随即选取数头个体,参照De Barro和Driver(1997)的方法提取烟粉虱个体总DNA[12]。
1.2.2 PCR反应条件
参照罗晨等(2002)的方法,扩增mtDNA COI基因片段采用的引物为C1-J-2195(5′-TTGATTTTTTGGTCATCCAGAAGT-3′)和L2-N-3014(5′-TCCAATGCACTAATCTGCCATATTA-3′)[13]。PCR扩增产物是mtDNACOI3′末端的部分序列,大小约为840 bp。在0.2 mL PCR管中加入50 μL PCR反应体系,包含1 μL TaqDNA酶,2 μL DNA模板,2 μL引物C1-J-2195((20 μmol/L),2μLL2-N-3014((20 μmol/L),1μL dNTP(10 mmol/L),5 μL 10×PCRbuffer。反应体系于94°C下预变性4 min;然后进行38个如下循环:94 ℃变性1 min;50 ℃退火1 min;72 ℃延伸1 min;循环结束后72 ℃延伸5 min,10 ℃保存。反应产物可置于4 ℃冰箱保存或-20 ℃长期保存。
电泳观察PCR结果:琼脂糖电泳检测PCR产物。然后在紫外光下切下条带较亮的目的片段,置于离心管中,进行回收。
1.2.3 PCR产物的纯化及送样测序
使用OMEGABIO-TEK纯化试剂盒对回收的PCR产物进行DNA纯化。将纯化好的产物连同各5 μL的上下游引物一起交由上海生工进行测序。
1.2.4 序列分析
将上海生工提供的DNA序列在NCBI上与国内外代表地区的烟粉虱COI序列进行比对,如比对结果显示同此序列相似性最高的前几个序列为B型或Q型烟粉虱的话(以相似性在99%-100%为标准),则可以初步认为此序列所代表的该烟粉虱个体为B型或Q型烟粉虱。随机选取西安Q型和延安B型的一条测序结果提交GenBank,序列登录号分别是:KT235801和KT235802。提取GenBank和文献中已经明确为Q型、B型和其它烟粉虱的mtCOI序列,并将非洲小粉虱Bemsiaafer(Prienser et Hosey)作为外群,利用MEGA5.1软件,根据Kimura2-paramter模型,采用邻位法(NJ)进行聚类分析构建系统树,系统树各分支置信度(bootstrap)均进行1000次重复检验。系统发育分析中所用的所有mtCOI序列相关信息见表2。
2 结果
2.1 主要寄主危害程度及不同生物型烟粉虱地理分布
通过对陕西地区主要经济作物种植区的调查,共记录到烟粉虱危害的主要经济作物17种(变种)隶属4个科,分别为葫芦科4种、茄科5种、十字花科4种、豆科4种。调查分析发现,在陕西地区烟粉虱主要危害的植物是茄科的烟草、甜椒、番茄以及葫芦科的黄瓜、西葫芦,在这些寄主植物的叶片上虫口密集,植株受害严重。普遍受害较轻的是十字花科植物,受害最轻的是茄科的马铃薯和豆科的豇豆、豌豆(见表1)。
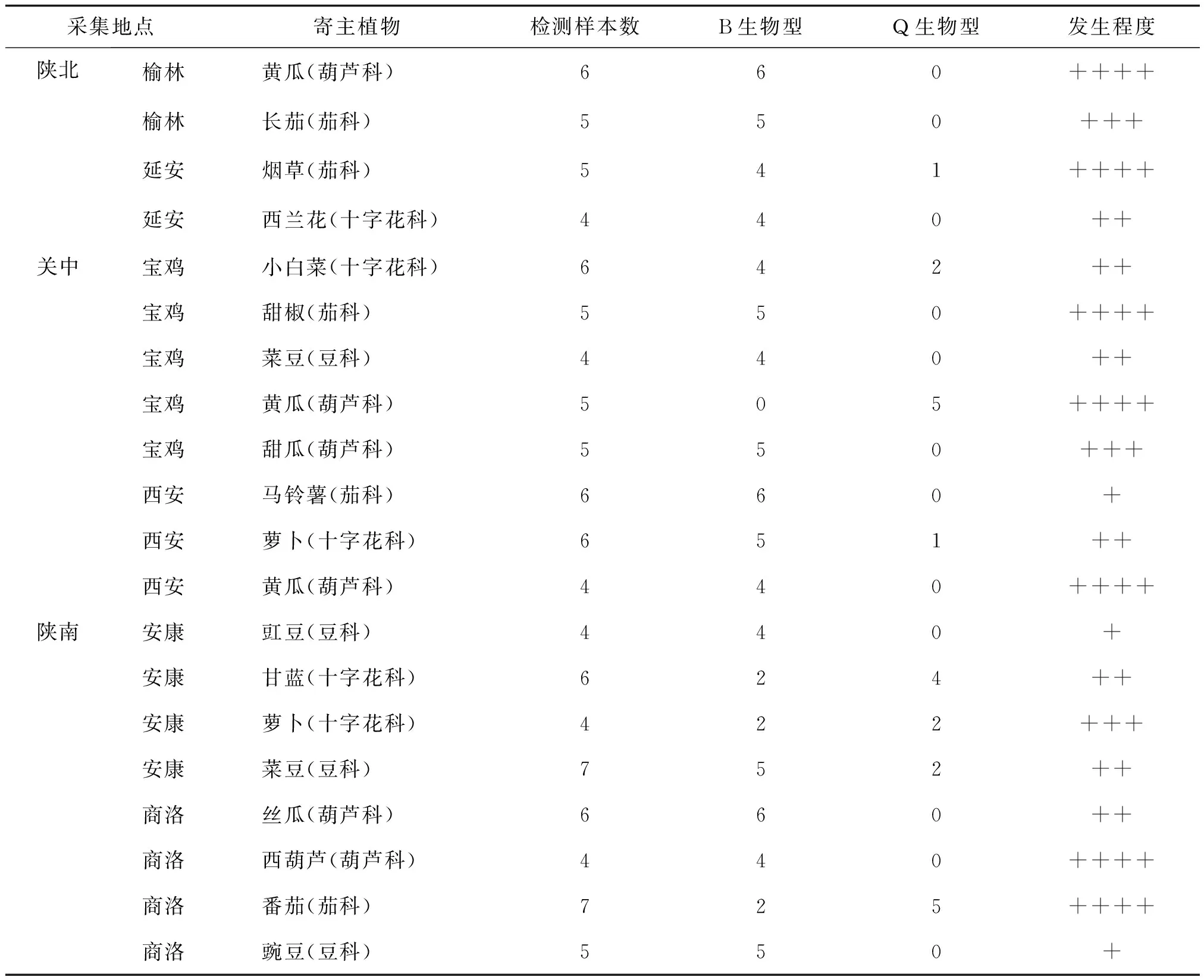
表1 陕西地区烟粉虱生物型分布及主要寄主发生危害程度
通过同源性分析,陕西地区危害的烟粉虱以入侵型B型的比例最大,为78.85%;入侵型Q型的比例为21.15%,没有检测到其它生物型的存在。从不同生态区来说,B型烟粉虱的检出率分别为陕北95.00%、关中80.49%和陕南69.77%,在陕北B型烟粉虱危害更为严重,在陕南Q型烟粉虱危害在加重。从采集寄主上看,B型烟粉虱在所有寄主上都发生危害,而仅在8个采集点7种寄主上检测到Q型烟粉虱,其中在葫芦科和茄科等经济作物上发生危害比较严重,主要对黄瓜和番茄等寄主植物的危害较大。
2.2 生物型检测及其系统发育分析
在陕西三个不同生态区域的20个采集样点,共采集了104个样本,所有样品测序后均得到840bp左右的片段(见图1),分别从每个所测序列中截取720bp的序列进行分析,通过DNAMAN分子生物学分析软件结合基因库序列比对分析发现,104个烟粉虱样本的COI基因序列中有82个相同(B型),其余22个COI基因序列一致(Q型)。
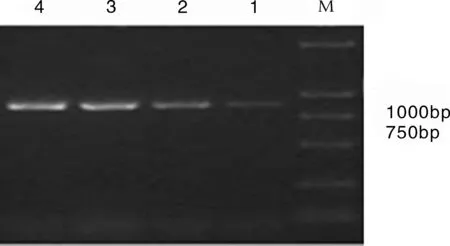
1-4:烟粉虱样本;M:100 bp DNAMarker
图1 使用引物C1-J-2195和L2-N-3014对烟粉虱个体进行线粒体COI基因扩增后得到的电泳图
与已知序列比较(见表2),构建系统树,进行聚类分析,建树时以非洲小粉虱(AF418673)为外群,使用邻位法(NJ)法和非加权配对算数平均法(UPGMA)。UPGMA法和NJ法的系统发育构建结果相似,所以在此只给出采用NJ法构建的系统发育树(见图2)。由构建的系统进化树可以发现,非洲小粉虱、A型、B型、Q型不同生物型的烟粉虱遗传距离较大,均可以明显的分开。Q-KT235801与已知Q型烟粉虱序列Q-FJ375346(湖北)、Q-EU881713(危地马拉)、Q-DQ365876(葡萄牙)等的同源性高达100%,显示Q-KT235801与已知Q型烟粉虱种群聚为同一进化枝,可以确定西安的mtCOI序列所代表的烟粉虱为Q型烟粉虱;B-KT235802与已知B型烟粉虱序列B-AB204577(日本)、B-AJ550170(法国)、B-FJ647211(扬州)等的同源性高达100%,显示B-KT235802与已知B型烟粉虱种群聚为同一进化枝,可以确定延安的mtCOI序列所代表的烟粉虱为B型烟粉虱。上述序列比对与同源性分析结果显示,陕西不同采集样点的烟粉虱种群同时存在B型和Q型。
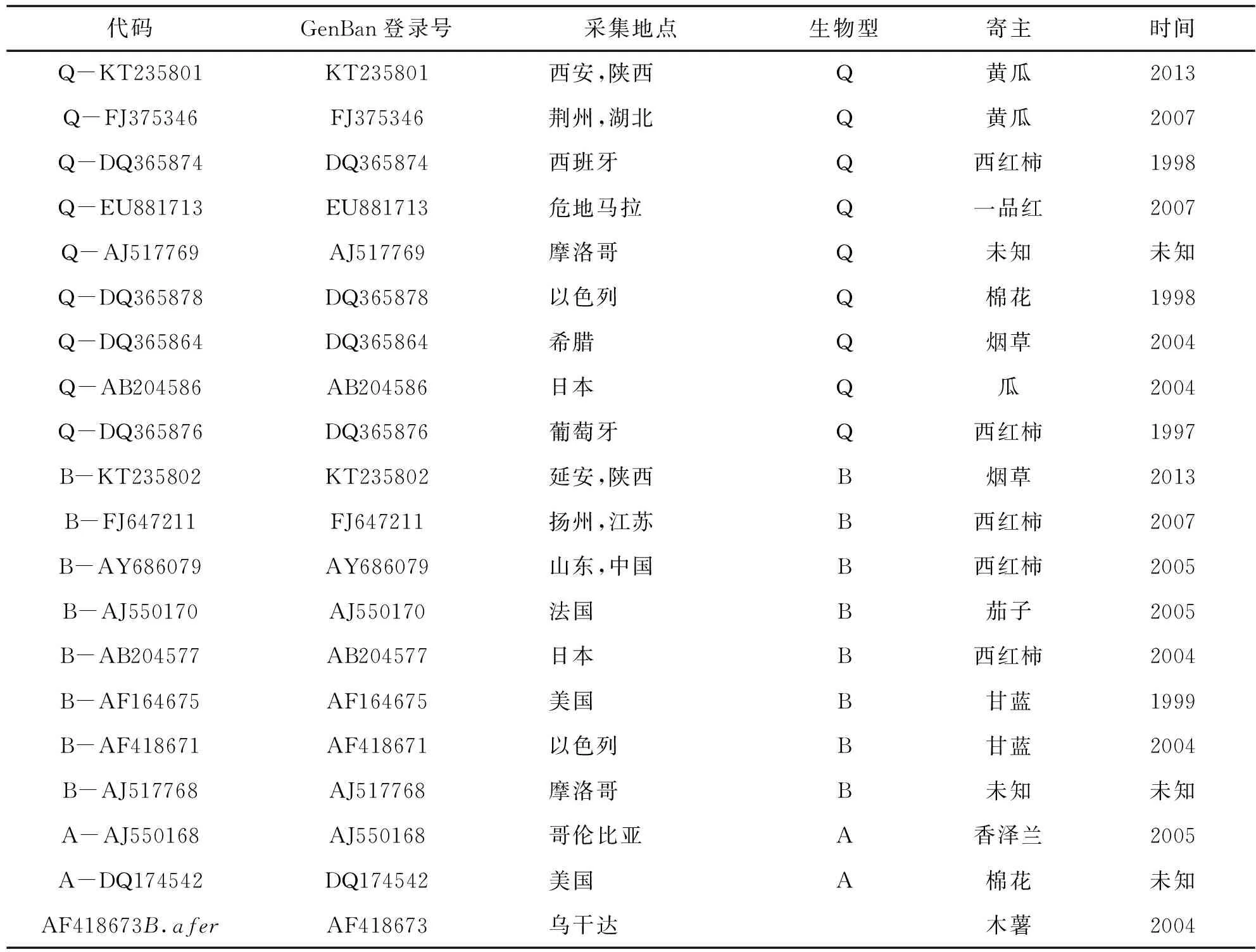
表2 系统发育分析中所涉及的相关mtCOI序列信息

图2 基于mtCOI序列的不同生物型烟粉虱的系统发育分析,外群为非洲小粉虱(AF418673)
3 讨论
烟粉虱是由在形态学上难以区分并且存在明显遗传分化的多个遗传群组组成的复合种,根据其在寄主范围、传毒能力和遗传特性等方面的不同,烟粉虱又被分为至少24种生物型。因此,对烟粉虱生物型的监测是烟粉虱综合治理的基础,也是优化烟粉虱防控策略的一项重要工作。目前在我国至少存在6个烟粉虱单倍型。本世纪初烟粉虱在我国的传播以B型为主并且逐渐取代白粉虱和当地烟粉虱[14]。此后随着Q型烟粉虱的入侵,有逐渐取代B型成为优势生物型的趋势[15]。
本文作者曾于2009~2011年对全国多个烟粉虱地理种群进行过较为系统的研究,发现在绝大部分地区Q型烟粉虱发生频率逐年升高,但当时陕西只检测到B型烟粉虱[11]。2013年我们采集了陕西省多个地区的烟粉虱种群,涉及4科17种寄主植物,可进一步监测不同生物型烟粉虱在本地区的遗传分布特性和危害程度。从本文的调查结果发现,烟粉虱在陕西地区主要经济作物上的寄主数量及分布范围都呈上升趋势,危害越来越严重,已成为该地区农业生产中的严重威胁之一。根据对陕西地区烟粉虱mtDNACOI基因序列分析结果看,陕西地区发生的烟粉虱主要为B型,占所测总样本的78.85%,其分布面积大,危害程度高;Q型烟粉虱在部分寄主上发生,危害程度也高,说明在陕西地区Q型烟粉虱发生频率加快。同时,试验结果还表明,Q型烟粉虱在陕北、关中和陕南的发生比例相差较大,陕北最低5%,陕南最高30.23%,说明Q型烟粉虱在陕西地区的扩散频率从北到南逐渐加强,并有取代B型烟粉虱的趋势。这可能与陕西不同地区的气候有关,从陕北到陕南年均温度逐渐升高,年均温度影响了Q型烟粉虱对B型烟粉虱的种群替代[11]。在年均温度比较高的陕南Q型烟粉虱更容易发生。因此,应加强对陕西地区不同生物型烟粉虱传播的监控。
通过系统聚类树分析发现,陕西地区的B型和Q型烟粉虱分别与国内外其他地区的B型和Q型烟粉虱聚为一支。从对陕西地区烟粉虱的寄主危害程度的调查发现,在葫芦科和茄科等经济作物上烟粉虱发生危害比较严重,其中B型烟粉虱主要对黄瓜、烟草、甜椒和西葫芦等寄主植物的危害较大,Q型烟粉虱主要对黄瓜和番茄等寄主植物的危害较大。本文初步明确了陕西地区烟粉虱的寄主范围、危害程度及主要生物型,对于合理、准确地进行烟粉虱的危害预测和综合治理具有十分重要的意义。
[1]Jones D R.Plant viruses transmitted by whiteflies[J].European Journal of Plant Pathology,2003(109):195-219.
[2]Delatte H,Reynaud B,Granier M,et al.A new siliverleaf-inducing biotype Ms ofBemisiatabaci(Hemiptera:Aleyrodidae)indigenous to the islands of the south-west Indian Ocean [J].Bulletin of Entomological Research,2005(95):29-35.
[3]吴杏霞.我国B型烟粉虱的发生、分布以及分子鉴定[C].中国农业大学,2002.
[4]罗晨,张君明,石宝才,等.北京地区烟粉虱Bemisiatabaci(Gennadius)调查初报[J].北京农业科学,2000,18(增刊):42-47.
[5]褚栋,张友军,毕玉平,等.警惕Q型烟粉虱在我国进一步扩散[J].植物检疫,2005b,3(19):171-174.
[6]褚栋,张友军,丛斌,等.云南Q型烟粉虱种群的鉴定[J].昆虫知识,2005d,1(42):54-56.
[7]徐婧,王文丽,刘树生.Q型烟粉虱在浙江局部地区大量发生危害[J].植物保护,2006,32(4):121-124.
[8]徐文华,左文惠,王瑞明,等.烟粉虱种群在江苏沿海城市市区的寄主分布与虫源性质[J].华东昆虫学报,2007,(16)3:187-196.
[9]谭永安,肖留斌,孙洋,等.江苏烟粉虱生物型鉴定与危害程度分析[J].环境昆虫学报,2012,34(3):277-282.
[10]万方浩,郭建英.农林危险生物入侵机理及控制基础研究[J].中国基础科学,2007,9(5):8-14.
[11]赵瑞华.内共生菌对烟粉虱种群形成与发展的影响及其作用机制[C].中国农业科学院博士后论文,2013.
[12]De Barro P J,Driver F.Use of RAPD-PCR to distinguish the B biotype from other biotypes ofBemisiatabaci(Gennadius) (Hemiptera:Aleyrodidae)[J].Australian Journal of Entomology,1997,2:149-152.
[13]罗晨,姚远,王戎疆,等.利用mtDNA COI基因序列鉴定我国烟粉虱的生物型[ J ].昆虫学报,2002,45(6):759-763.
[14] 臧连生,刘树生,刘银泉,等.浙江B 型与一非B 型(China-ZHJ-1)烟粉虱形态学和生物学特性的比较研究[J].昆虫学报,2005,48(5):742-748.
[15]滕希.Q型烟粉虱在中国的发生现状及基于mtCOI序列的系统发育分析[C].中国农业科学院硕士学位论文,2009.
[责任编辑 李晓霞]
Biotypes Identification and Damage Analysis ofBemisiatabaci(Gennadius) in Shaanxi Area
ZHAO Rui-hua,DENG Zhen-shan,HE Xiao-long
(College of Life Sciences,Yan′an University,Yan′an 716000,China)
To evaluate main biotypes and damage ofBemisiatabaci(Gennadius)in Shaanxi area,we analyzed the extent of damage through investigation and identified the variety of biotypes in Shaanxi main host plants by mtDNA COI sequences ofBemisiatabaci.The sequence analysis results indicated that the B and Q biotypesB.tabacico-existed in Shaanxi area,and B biotypeB.tabaciwas 78.85%.The results showed that B biotype still was the main biotype,but Q biotype spreads widely in Shaanxi area and may replace B biotype as a major crop pest in Shaanxi area.B.tabacidamaged crops seriously such as tobacco,sweet pepper,tomato of solanaceae and cucumber,pumpkin of cucurbitaceae.But the plants of cruciferae were damaged lightly,and damagous to potato of solanaceae and cowpea, peas of leguminosae were lightest.
Bemisiatabaci; biotype; damage; host plant
2015-09-25
陕西省教育厅专项科研计划项目(14JK1822);延安大学博士科研计划项目(YDBK2013-3);延安市科学技术研究发展计划项目(2014CGZH-09);陕西省烟草公司科技专项项目(YAYC-2014-FW-01)
赵瑞华(1978—),女,山东泰安人,延安大学讲师,博士。
S435
A
1004-602X(2015)04-0082-06
猜你喜欢
今日农业(2022年15期)2022-09-20
中国农业科学(2022年13期)2022-07-26
作文·初中版(2019年8期)2019-09-09
植物保护(2018年5期)2018-12-05
生物学教学(2018年3期)2018-08-08
中学生物学(2018年8期)2018-03-01
湖北农业科学(2017年20期)2017-11-17
中国社区医师(2016年8期)2016-12-20
全国新书目(2009年9期)2009-06-10
现代农业科技(2009年19期)2009-03-20
