
亚高山草地土壤原生生物群落结构和多样性海拔分布格局
2021-04-25 06:26:14罗正明刘晋仙周妍英杜京旗柴宝峰
生态学报 2021年7期
罗正明,刘晋仙,周妍英,杜京旗,吴 强,柴宝峰,*
1 忻州师范学院地理系,忻州 034003
2 山西大学黄土高原研究所,黄土高原生态恢复山西省重点实验室,太原 030006
3 忻州师范学院生物系,忻州 034003
4 山西省五台山国有林管理局,繁峙 034300
亚高山生态系统是陆地系统重要的组成部分,具有涵养水源、调节气候、维持生物多样性等重要的生态服务功能[1]。海拔梯度变化综合了多种环境因子的梯度效应。随着亚高山海拔梯度的上升,土壤、温度和湿度等环境因子均发生改变,从而引起土壤微生物群落多样性的变化[2]。例如,土壤pH、碳氮比和植被类型通常被认为是土壤微生物多样性海拔分布模式的关键驱动因素[3-4]。目前,土壤微生物海拔分布模式和群落构建机制并不明确,已有研究都集中在一个完整的或较大范围的海拔梯度上,具有较大的海拔间隔和不同的植被类型[3, 5-6]。生物多样性采样的尺度会强烈地影响所观察到的生物分布模式[7],因此,在小尺度海拔梯度范围内单一的生态系统中,能够更好地反映环境因素和地理距离对微生物群落结构和多样性分布格局的影响。
微生物群落构建的理论基础主要包括生态位理论和中性理论。传统的生态位理论假设环境条件、物种特征、物种间相互作用等因素控制着群落结构,这通常被称为确定性过程[2, 8]。相反,中性理论假设群落结构独立于物种特征,受随机过程(如物种形成、灭绝、定殖和扩散限制)控制[8]。越来越多的研究表明,确定性过程和随机性过程在微生物群落构建过程中不是独立存在的,而是共同起作用的[2, 9-10]。如今更多的研究趋向于生态位理论和中性理论的整合,探究群落构建过程中随机作用和确定性作用的相对贡献[11]。目前关于微生物群落多样性维持机制的研究主要集中于细菌和真菌[4-6],原生生物作为土壤微生物的第三大生物区系,其多样性分布模式与群落构建机制目前还没有系统的研究。
原生生物是土壤微生物群落的重要组成部分,在土壤微生物组中起到枢纽作用[12],直接参与细菌、真菌和其他微小真核生物的群落调节过程,在微生物食物链、土壤营养转化和生态系统稳定性等方面发挥重要作用[13-14]。尽管原生生物对土壤功能具有普遍性和生态重要性,但对原生生物群落的认知远远落后于其他微生物群体(细菌和真菌等)。之前针对异养类群(原生动物),包括土壤纤毛虫(ciliate)和变形虫(amoebae)等原生动物的群落结构、时空分布、食性与功能营养类群,以及人类干预土壤环境(如土地利用方式、放牧和氮沉降)对纤毛虫的影响开展了一些研究[15-17]。但这些研究大多针对单类群原生生物[17],目前在群落水平对原生生物多样性维持机制的研究还不够全面和深入。近年来,原生生物数据库以及基于18S rDNA高通量测序技术方法的进一步优化,为探索原生生物群落多样性和生态重要性提供了新的机会[14]。在有限的研究中,已经证实土壤条件(如pH、营养素和水分),以及农药用量和其他扰动显著影响了整个土壤原生生物群落组成和结构[13, 17-18]。尽管原生生物群落对生物和非生物因素及其复杂的组合做出了不同程度的反应,但它们之间的相互作用机制仍远未被理解。
五台山位于山西省东北部,主峰叶斗峰为华北最高峰,高达3061 m,山顶属高寒气候,土壤和植被垂直带谱明显,各种植被类型保护完整,是中国华北境内生物多样性最丰富的地区之一[19]。五台山海拔2000 m以上广泛分布的亚高山草地是华北地区保存较为完好、相对稳定的典型亚高山草地生态系统,对全球气候变化敏感[2]。这为群落生物多样性海拔分布格局及构建机制的研究提供了一个天然平台。本研究我们选择五台山亚高山草地生态系统中不同海拔梯度土壤原生生物群落为研究对象,旨在探讨:(1)沿海拔梯度,草地土壤原生生物群落结构组成和多样性分布模式;(2)确定土壤原生生物群落构建的主要驱动因子;(3)确定性过程和随机过程在土壤原生生物群落构建过程中的相对作用。
1 材料与方法
1.1 样品采集
2017年8月,在五台山北台(叶斗峰)沿海拔高度 2000—3061 m 范围分布的亚高山草地,设置了 4个不同海拔梯度样带:E1(2000—2100 m)、E2(2300—2400 m)、E3(2600—2700 m)和E4(2900—3000 m),每个海拔梯度包含五个不同的样地。在每个样地,设置3个1 m×1 m的样方,样方之间间隔约20 m。用土钻采集表层土壤样品(0—10 cm),每个样方内采集5个子样(每个角1个,中心1个),在聚乙烯袋中混合成一个样品。经2 mm网筛,将大部分根、动物和石头去除,然后将样本分成两部分。其中一部分风干后进行理化分析,另一部分保存在-80℃冰箱中,用于DNA的提取。在每个样方中调查植被参数,记录每种植物的名称、高度、盖度及多度等指标。植物的丰富度指数(richness)采用单位面积内的物种数目来测度物种的丰富程度。
1.2 土壤理化性质测定
1.3 DNA提取、高通量测序及生物信息学分析
称取0.25 g土壤样品,使用E.Z.N.A.®土壤DNA试剂盒(Omega Bio-tek, USA)按照说明书提取和纯化微生物DNA。将每个采样点的3个土壤样品等体积混合,一共20个DNA样品,送往上海美吉生物医药科技有限公司进行高通量测序。采用TAReuk454FWD1(正向引物5′-CCAGCASCYGCGGTAATTCC-3′)和TAReukREV3(反向引物5′-ACTTTCGTTCTTGATYRA-3′)对原生生物18S rRNA 基因的V4高变区进行PCR扩增,并通过Illumina Miseq测序平台对扩增产物进行测序。
原始序列经过质控后,剩下的序列使用UPARSE以97%的相似性作为阈值划分分类操作单元(operational taxonomic units,OTUs)。然后根据原生生物核糖体参考数据库(Protist Ribosomal Reference database,PR2)对原生生物OTUs进行分类学比对。得到原生生物OTUs分类水平数据表,根据Zhao等人[20]的方法将门水平中的红藻门(Rhodophyta),链形植物(Streptophyta),真菌(Fungi),后鞭毛生物(Opisthokonta),后生动物(Metazoan)和模糊分类群剔除,然后进行后续的分析。
1.4 数据分析
基于R studio(v.3.4.3)vegan包,计算原生生物和植物的α多样性(Shannon指数和Richness指数)和β多样性,采用非度量多维尺度分析(non-metric multidimensional scaling,NMDS)和相似性分析(analysis of similarity,ANOSIM)对不同海拔梯度间原生生物群落结构进行比较。LEfSe分析(linear discriminant analysis effect size)检测组间原生生物群落(从超群到科共5个分类学水平)具有显著丰度差异类群(P<0.05和LDA评分>3.0),采用线性判别分析(linear discriminant analysis,LDA)来估算每个物种丰度对差异效果影响的大小,识别潜在的生物标记物(biomarker)。根据各采样点地理坐标数据,采用邻体矩阵主坐标(principal coordinates of neighbor matrices,PCNM)分析空间变量(R studio(v.3.4.3)vegan包中的PCNM函数)。采用冗余分析(redundancy analysis,RDA)评价原生生物群落结构与环境变量之间的相关性。在RDA之前,使用逐步回归和蒙特卡罗置换测试对所有环境变量进行前选择,选择具有统计学意义(P<0.05)的环境变量进行下一步分析。采用方差分解分析(variance partitioning analysis, VPA)分析环境变量和地理距离对原生生物群落结构变化的相对贡献(CANOCO 5.0)。采用Mantel和偏Mantel检验评价原生生物群落与环境变量和地理距离的相关性。零模型分析方法使用R studio(v.3.4.3)reldist、vegan和bipartite程序包,参照Tucker等[21]和Chase[22]构建的β-零偏差值来分析确定性过程和随机过程在群落构建中的相对重要性,[(βobs-βnull)/βnull]偏离零表示群落的构建是确定性过程决定的(βobs:观测值(observed β-diversity),βnull:期望值(null-model β-diversity)),正值表示种间竞争,负值表示环境选择,接近零表示随机过程在群落构建中起决定作用。利用IBM SPSS statistics 20进行皮尔逊相关性分析(pearson correlation test)以及采用单因素方差分析(one-way analysis of variance)和Duncan多重比较分析进行显著性差异分析。所有统计分析的显著性水平均为P<0.05。
2 结果与分析
2.1 土壤理化性质和植被参数与海拔梯度的关系
不同海拔梯度的土壤理化参数和植物多样性数据可见表1。
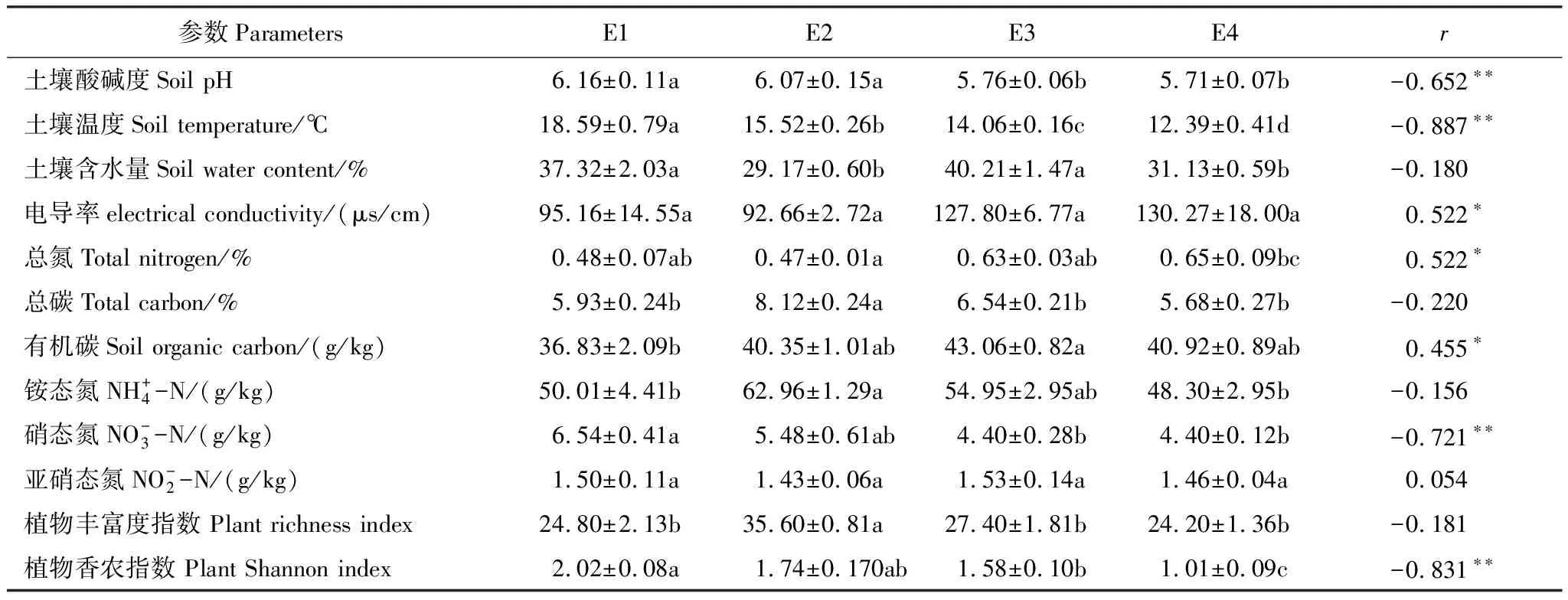
表1 不同海拔梯度下土壤理化性质与植物多样性参数
海拔梯度与土壤总氮含量(r=0.522,P<0.05)、有机碳含量(r=0.455,P<0.05)和电导率(r=0.522,P<0.05)之间具有显著的正相关关系,但与土壤 pH(r=-0.652,P<0.001)、土壤温度(r=-0.887,P<0.001)、硝态氮(r=-0.721,P<0.001)和植物Shannon指数(r=-0.831,P<0.001)之间具有显著的负相关关系。土壤含水量(r=-0.180,P>0.05)、总碳(r=-0.220,P>0.05)、铵态氮(r=-0.156,P>0.05)、亚硝态氮(r=0.054,P>0.05)、植物richness指数(r=-0.181,P>0.05)与海拔梯度没有显著相关关系。以上结果表明,五台山亚高山草地土壤理化因子和植被参数沿海拔梯度发生了明显的变化。
2.2 原生生物群落组成和多样性沿海拔的分布格局
在对非原生真核生物进行筛选和去除后,剩余的520673条优质序列聚类成1980个原生生物OTU,然后按最小样本序列数抽平,共鉴定出1734个OTU。20个样地共鉴定出8个超群,包括有孔虫界(Rhizaria)、变形虫界(Amoebozoa)、后鞭毛生物(Opisthokonta)、不等鞭毛类(Stramepopila)、囊泡虫类(Alveolata)、Hacrobia、泛植物界(Archaeplastida)、古虫界(Excavata)。24个门、65个纲、125个目、222个科、350个属。其中丝足虫门(Cercozoa)、褐藻门(Ochrophyta)、纤毛门(Ciliophora)、顶复门(Apicomplexa)、锥足亚门(Conosa)、绿藻门(Chlorophyta)和叶足亚门(Lobosa)被定义为优势门(相对丰度>1%)(图1)。图1中可知,四种海拔梯度下原生生物群落优势门种类基本一致,但它们相对丰度存在差异。
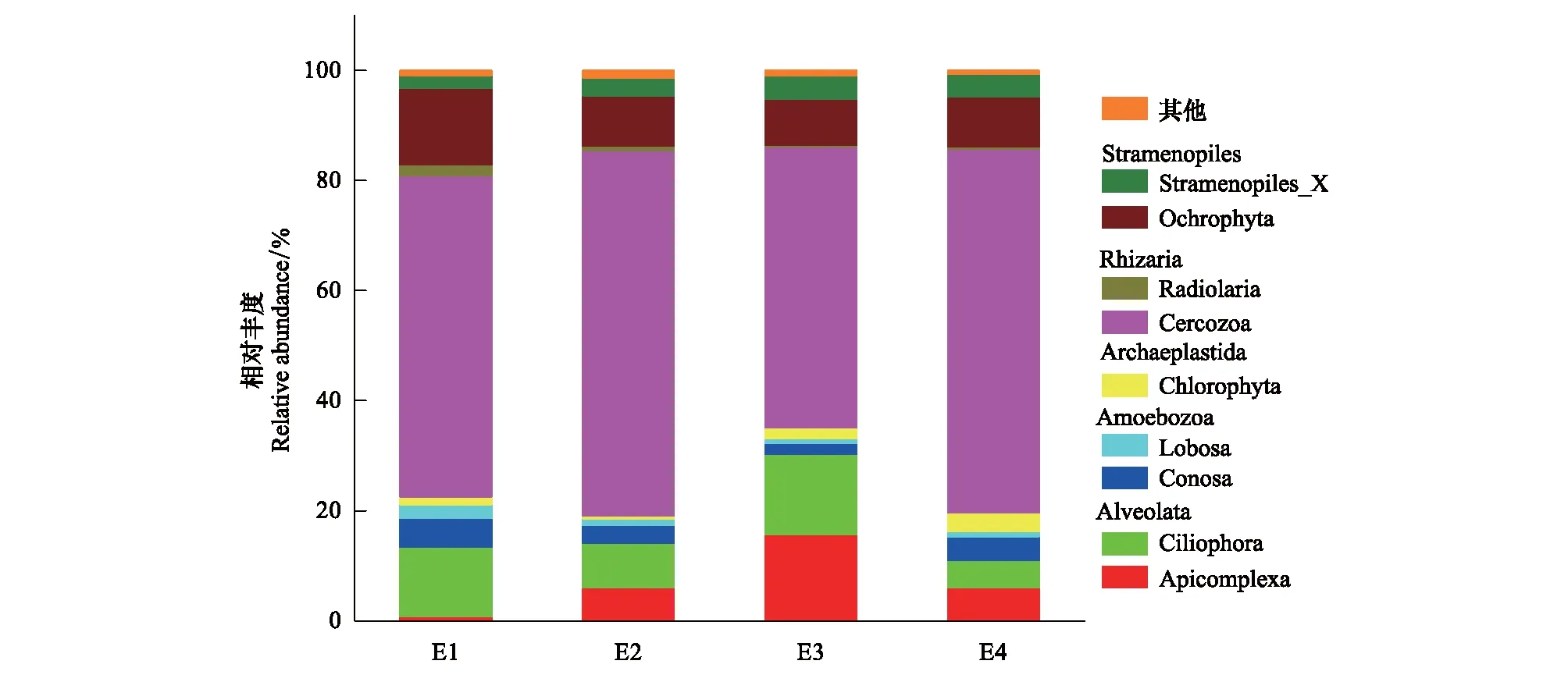
图1 不同海拔梯度土壤原生生物群落优势门(平均相对丰度> 1%)的相对丰度
LEfSe分析显示(图2),17个生物标志物对海拔梯度非常敏感(P<0.05, LDA>3.0)。在海拔梯度E1中富集了4种显著生物标志物,分别为放射虫门(Radiolaria)、等幅骨虫纲(Acantharea)、节棘目-粘合棘目(Arthracanthida-Symphyacanthida)、Arthracanthida-Symphyacanthida_X;E2中显著富集了3种生物标志物,包括有孔虫界(Rhizaria)、丝足虫门(Cercozoa)和Glissomonadida;E3中显著富集了2种生物标志物,篮管纲(Nassophorea)和篮管纲中未定义的目(Nassophorea_X);E4中显著富集了泛植物界(Archaeplastida)、绿藻门(Chlorophyta)、共球藻纲(Trebouxiophyceae)、溪菜目(Prasiolales)、溪菜目中未定义的科(Prasiolales_X、Sandonidae)、Filosa-Sarcomonadea和袋形目中未定义的科(Bursariomorphida_X)共8种生物标志物。这些结果表明,亚高山草地土壤原生生物群落组成和相对丰度具有明显的海拔分异。
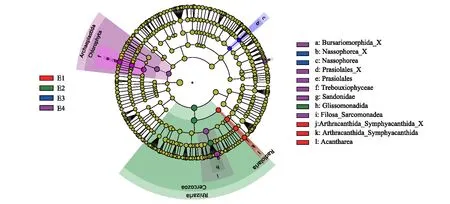
图2 不同海拔梯度土壤原生生物群落组成差异的LEfSe分析结果
四个海拔梯度间的土壤原生生物群落 Shannon 指数和丰富度指数均没有显著差异(图3)。基于Bray-Curtis距离的非度量多维尺度分析(NMDS)和ANOSIM结果表明,沿海拔梯度土壤原生生物群落结构发生了显著的变化,4个海拔梯度间的土壤原生生物群落结构存在显著的差异(r=0.496,P<0.01)(图4)。土壤原生生物群落β多样性(Bray-Curtis距离)与海拔距离(欧式距离)呈显著正相关关系(r=0.375,P<0.001),土壤原生生物群落之间的相异性随着海拔距离的增加而显著升高(P<0.01)(图5),表明选择过程可能是亚高山草地土壤原生生物群落分布格局的主要驱动因素。
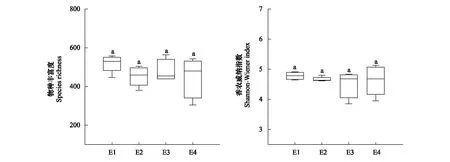
图3 不同海拔梯度土壤原生生物群落α多样性
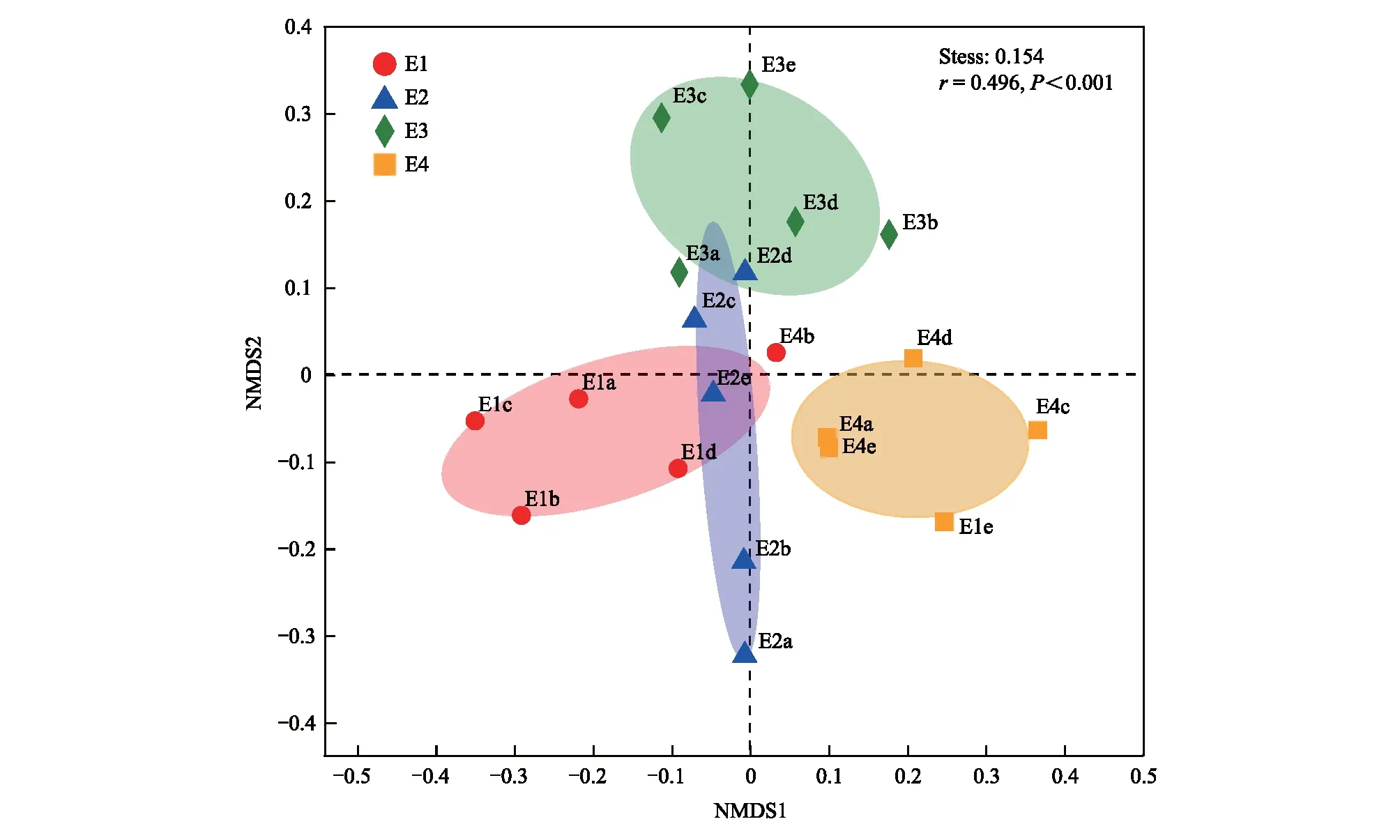
图4 基于Bray-Curtis距离的不同海拔梯度土壤原生生物群落非度量多维尺度分析(NMDS)
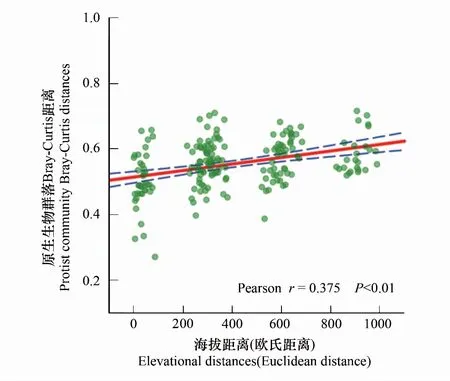
图5 土壤原生生物β多样性与海拔距离(欧式距离)的关系
2.3 环境因子和空间变量对原生生物群落的影响
RDA和蒙特卡罗置换测试(Monte Carlo permutation test)表明,海拔、土壤含水量、总氮和植物丰富度指数对原生生物群落结构有显著的影响(P<0.05),RDA轴1和轴2分别解释变异量的25.93%和10.94%(图6)。偏Mantel检验发现环境因子(P<0.01)和空间变量(P<0.05)与原生生物群落的相异性均显著相关(表2),表明原生生物群落相似性具有显著的距离衰减关系。VPA分析发现环境因子和空间变量共同解释了原生生物群落变异的28.7%,其中环境因子单独解释了7.9%,空间变量单独解释了1.8%,环境因子对群落结构变异的贡献远大于空间变量(图7)。说明土壤原生生物群落构建过程中,现代环境比空间变量(地理距离)起着更重要的作用。根据以上结果推断,在土壤原生生物群落构建过程中,确定性和随机性过程都起作用,而且确定性过程占绝对的优势。
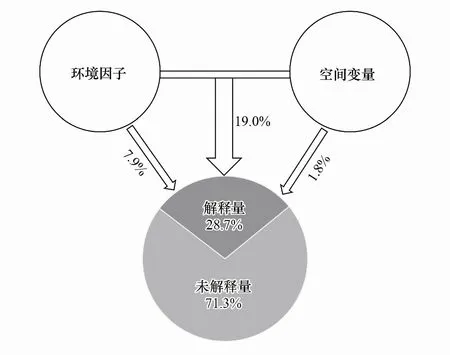
图7 环境因子和空间变量解释原生生物群落变异百分比的方差分解分析(VPA)

表2 通过偏Mantel分析原生生物群落相异性、环境因子和空间变量之间的关系
2.4 零模型推断群落构建过程
通过基于β多样性的零模型方法分析了沿海拔梯度确定性过程和随机性过程在原生生物群落构建过程中的相对重要性(图8)。4个海拔梯度间的零偏差值没有显著的差异,且均小于零(-0.352—-0.404),推断4个海拔梯度下土壤原生生物群落构建过程中环境选择均比随机过程在起的作用要大。说明确定性过程驱动了研究区域土壤原生生物群落的构建过程。
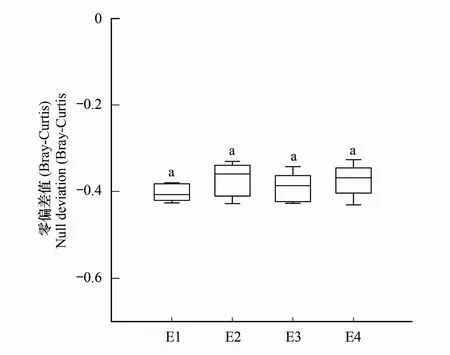
图8 不同海拔梯度土壤原生生物群落的零偏差
3 讨论
原生真核生物形体微小,结构简单,分布广泛,生命周期短,种类多样,对扰动反应迅速,是维持微生态平衡的主要参与者。对原生生物群落结构和多样性维持机制的了解,有助于深入研究全球变化对生态系统结构和功能的影响。本研究结果显示,五台山亚高山草地土壤原生生物群落的优势类群在四个海拔梯度下基本一致(图1),但在相对丰度上存在差异,而且每种海拔梯度都有其独特的微生物种群(图2),说明土壤原生生物群落组成和相对丰度具有明显的海拔分异。本研究区的优势门包括,丝足虫门(Cercozoa)、褐藻门(Ochrophyta)、纤毛门(Ciliophora)和顶复门(Apicomplexa),这与以往对德国温带草原土壤、中国东北黑土的调查一致,但与绿藻门(Chlorophyta)、丝足虫门和褐藻门占主导地位的湖南丘陵红色土壤不同[20,23]。
亚高山草地生态系统中微生物群落结构对海拔梯度的响应敏感。我们的结果表明,海拔对土壤原生生物群落的α多样性没有显著影响(图3),这与之前本课题组在该研究区对土壤细菌和真菌的研究结果不一样,细菌群落α多样性随海拔高度的增加而增加[2],而真菌群落α多样性随海拔高度的增加呈线性减少[24],表明不同类型微生物群落α多样性对海拔梯度的响应模式不同。原生生物α多样性在不同海拔之间并没有显著差异,这可能是由于已知因子(包括大风和动物活动)造成的出人意料的高效扩散。此外,许多原生生物物种可以形成休眠的原生生物包囊(cyst)种子库[13],即使在严酷的环境条件下,也可以形成大量的繁殖体,可能导致各海拔梯度土壤原生生物群落组成发生了明显的变化,但是α多样性变化不显著。亚高山草地土壤原生生物群落结构(β多样性)随海拔梯度变化显著(图4),并且土壤原生生物群落之间的距离(相异性)随海拔距离增加而显著增加(图5),与五台山亚高山生态系统土壤细菌和真菌的β多样性海拔分布模式相同[2,24]。这表明环境选择过程可能是不同海拔梯度土壤原生生物群落分布模式的主要驱动机制。同时,VPA分析和偏Mantel分析结果表明(表2和图7),环境因子和空间变量对原生生物群落海拔分布格局均有显著影响,但是环境因子的相对作用(7.9%)明显大于空间变量(1.8%),这表明在五台山亚高山草地生态系统中土壤原生生物群落沿海拔梯度的构建过程中,确定性过程(中性理论)起主导作用。这与本研究区细菌和真菌的研究结果一致[2,24],说明微生物群落结构主要受到确定性过程(环境选择)的影响,而空间变量对群落构建的相对作用较弱。通过零模型分析进一步证实了确定性过程在原生生物群落构建中的相对作用大于随机过程(图8)。因此,确定性过程主导了五台山亚高山草地海拔尺度下土壤原生生物群落分布模式的很大程度的变异。由于同一草地生态系统不同海拔区域之间的环境条件显著不同,在不同海拔上的原生生物群落在适应环境变化的能力也不同,对土壤微生物产生了环境过滤的作用,从而导致土壤原生生物群落的结构和组成在不同海拔存在明显的差异。
海拔、土壤含水量、总氮和植物丰富度指数是原生生物群落组成和结构变化的主要驱动因子(图6)。海拔是最重要的环境因子,它不仅可以通过调节小气候和养分的有效性,还可以通过地理距离来影响微生物群落[4]。海拔梯度范围内由于具有不同土壤性质的环境梯度,不同的气候带可能在很短的地理距离内出现[25]。海拔气候带可能是导致研究区土壤原生生物群落变化的主要原因。另外,沿海拔梯度的地理距离也会对原生生物的分布格局产生较大的影响(图4、图5和表2)。
土壤含水量是调节原生生物群落结构的另一个重要因素。土壤原生生物需要水来维持生命,它们的所有功能都受到土壤孔隙空间水分有效性的严格限制[18]。从北极到热带地区,土壤水分是调节土壤原生生物多样性、密度和群落组成的关键因素[26-28]。异养原生生物多样性通常在持续湿润的土壤中达到峰值[18],但某些类群(如网柄细胞状黏菌)随着干湿季节交替而变得更加多样化[29]。一项全球尺度范围的研究也证实,与气候条件有关的土壤水分有效性控制着原生生物群落[30]。
总氮含量也与原生生物群落结构显著相关,一些研究也报道了这一点[13,22,31]。氮等营养物质的形态也决定了土壤原生生物的多样性,纤毛虫、有壳变形虫和藻类的多样性和密度沿土壤氮梯度变化很大[31-32]。氮的添加增加了有壳变形虫的多样性和密度,与之相反,碳和磷降低了多样性和密度[33]。Zhao等[20]发现施肥可以塑造土壤微生物食物网的结构,原生生物是对施氮肥最敏感的微生物类群,可作为土壤变化的生物指标。
研究结果证实植物丰富度对原生生物群落结构产生的重要影响。原生生物与植物关系非常密切,植物和植被类型可以通过不同的方式影响土壤原生生物群落[32],包括凋落物或根系分泌物的质量差异、小气候的变化以及对细菌或真菌群落的影响。有研究表明,两种不同热带植物的根际差异显著影响其纤毛虫群落[34]。植物多样性越高,变形虫原生生物的丰度越高[35],在植被演替过程中,随着时间的推移,寄主变形虫物种丰富度同步增加[36]。
本研究中环境因子和空间变量的解释度都相对较低,还有约73.1%的变异无法解释(图7),这些无法解释的变异可能是由其他未测量但重要的环境因子、时间因素[2]或方法的局限[11]所造成的。另一个合理的解释是生物因素,如土壤原生生物与其他分类类群(细菌和真菌)之间的生物相互作用(竞争、捕食和被捕食等)[13,37],会影响群落的构建。在资源可获得性高的条件下,微生物间相互作用比非生物过滤更重要,而在高环境胁迫时期,非生物过滤的作用更显著[38]。因此,未来对原生生物群落的构建机制进行更系统的研究,微生物间相互作用的相对贡献是不可忽视的。
4 结论
亚高山草地生态系统中微生物群落结构对海拔梯度的响应敏感,群落间的相异性随着海拔距离的增加而显著升高。环境因子和空间变量都在原生生物群落构建过程中起作用,环境因子占绝对的优势。海拔、土壤含水量、总氮和植物丰富度指数是原生生物群落组成和结构变化的主要驱动因子。总之,确定性过程(环境选择)是五台山亚高山草地原生生物群落结构和多样性的主要驱动力。
猜你喜欢
散文诗(2021年22期)2022-01-12 06:14:04
数学物理学报(2021年6期)2021-12-21 06:24:38
应用数学(2020年2期)2020-06-24 06:02:50
环球时报(2019-05-23)2019-05-23 06:17:36
数学年刊A辑(中文版)(2018年2期)2019-01-08 01:59:52
少儿美术(快乐历史地理)(2018年7期)2018-11-16 05:31:12
绿色科技(2017年6期)2017-04-20 08:13:04
河北林业科技(2016年5期)2016-11-08 03:13:30
西南农业学报(2016年5期)2016-05-17 05:42:44
西藏大学学报(自然科学版)(2015年1期)2015-12-25 11:43:44
