
基于海马体位置细胞的认知地图构建与导航
2021-04-24 12:33阮晓钢张晓平
自动化学报 2021年3期
阮晓钢 柴 洁 武 悦 张晓平 黄 静
环境认知和导航是智能移动机器人必备的技能[1-2].人和动物可以在复杂环境中进行环境认知和导航,如觅食、归巢等[3],而目前移动机器人的认知和导航能力不足以达到动物的水平,因此,模拟动物的神经结构和认知机制,使机器人更加智能地进行环境认知和导航得到了越来越多的关注[4-7].一方面,用数学模型或人工智能算法辅助传统神经科学研究来验证大脑运作的多种假设理论,可以促进我们对动物环境认知心理和导航行为的理解;另一方面,动物的环境认知和导航机制为开发机器人的类脑空间认知模型提供了一个很好的视角,可以帮助我们设计和构造像动物一样进行自主环境认知的智能移动机器人[8-10].
啮齿类动物出色的导航能力,其原因长期以来假设为是内部神经空间的类地图的表达,也就是环境认知地图.美国行为心理学家Tolman[11]发现老鼠能在没有强化因素的情况下自由探索并学会迷宫的布局,初次提出认知地图(Cognitive map) 的概念,认为老鼠能利用对空间环境的内在表达进行环境的认知与记忆.之后,O'keefe 等[12]发现,当老鼠处于某一特定位置时,大脑海马体中有若干个神经细胞会一直处于活跃状态;而当其从这一位置离开,其他神经细胞则会变得活跃,这些细胞称为位置细胞(Place cells).位置细胞实时编码老鼠在环境中的位置信息,从而在老鼠大脑内部产生一个对其所在空间环境的拓扑表达,提供了以非自我为中心的认知地图(Allocentric cognitive map)[13].研究表明,在海马CA3 区域的位置细胞能形成对环境的独立表征[14].此外,海马体能够存储大量的场景,并在相同场景出现的时候进行场景复现,从而指导人或动物的导航行为.
从老鼠环境认知的机制中获得启发,研究人员通过对老鼠大脑认知机理进行建模,进一步探究动物的环境认知机理,并用于指导机器人环境认知与导航[15-16].Tolman 经典迷宫实验,包括T 巷迷宫实验(T-alley maze)、T 高架迷宫实验(T-elevated maze)、放射迷宫实验(Radial maze)、老鼠绕道实验(Tolman detour task)等,是对老鼠环境认知行为的探讨.其中,绕道实验(Detour task)用于研究不同的认知技能,如洞察力(Insight)、抑制控制(Inhibitory control)、路径规划(Route planning)等[17],其特点在于,更近的目标会产生更强的吸引力,使动物在遇到障碍物时很难摆脱认为的最短路径而进行绕道.构建老鼠大脑的数学模型或神经网络模型,用机器人对老鼠绕道实验进行再现得到了各国研究者们的关注[18].Schmajuk 等[19]提出可以存储空间位置和时间事件连接的递归联结网络(Recurrent associative networks)来构建空间和时间认知地图,能在Tolman 绕道实验中通过选择正确的子目标最终到达目标点,但其没有给出认知地图的形成过程与形象展示.Martinet 等[20]建立了一个可以为空间学习和行动规划提供分布式信息的前额皮质网络模型,展示了前额皮质柱能够从冗余的海马输入中学习到稀疏的拓扑-度量表示,再现Tolman 绕道实验并说明老鼠的认知洞察能力得益于老鼠大脑神经机制.Gao 等[21]提出了一种基于基底神经节的运动学习模型,从基底神经节中强化学习的动作-评价机制着手再现了Tolman 迷宫绕道实验,但并未涉及认知地图的形成机制.
2011 年,Alvernhe 等[22]用电生理学方式对Tolman 绕道实验中的局部重映射进行研究,认为位置细胞的局部重映射反映了迷宫局部结构的更新,使得老鼠在遇到熟悉的道路被阻断或被阻断的道路重新连通等情况时可以选择最优路径.此外,自组织图被认为是最接近人脑或生物神经系统的模型,其表现出了神经细胞群的拓扑特征映射关系,这与海马体位置细胞放电的机理非常相似.动态增长的自组织图可以在不需要预先指定其大小的情况下进行拓扑映射,具有较好的环境映射效果[23-24].
基于此,本文结合位置细胞和自组织图的特性,建立动态增减位置细胞认知地图模型 (Dynamic growing and pruning place cells-based cognitive map model,DGP-PCCMM),该模型使机器人通过与环境的交互激活可达点的位置细胞,构建并更新位置细胞认知地图;以构建的认知地图作为输入,运用位置细胞序列规划算法,进行机器人实时导航.本文的认知地图构建方法具有以下优点:1) 具有类脑仿生特性与一定的生物合理性;2) 能够适应动态环境(如门的开闭);3) 能够降低建图的复杂性.实验结果表明,DGP-PCCMM 能动态构建和更新认知地图,能初步完成对Tolman 老鼠绕道实验的再现,体现了其自组织与自学习特性.此外,本文进行了与栅格地图、动态窗口法的对比实验和与其他认知地图模型的讨论分析.结果表明,本文方法在所构建地图的简洁性、完整性和对环境中动态障碍适应性方面更有优势.
1 位置细胞认知地图模型
1.1 DGP-PCCMM 的“感知 — 响应”结构
DGP-PCCMM 具有如图1 所示类似自组织图的“感知—响应”结构,由感知层和响应层组成,或称由感知输入层和海马体位置细胞认知地图形成层组成.感知输入层VI中的点如同机体感受器,从外部环境中获得机器人的位置信息、场景图像、障碍物信息等;海马体位置细胞认知地图形成层VO,如同老鼠大脑皮质中海马记忆区,用来形成位置细胞认知地图.VO层中的每个节点对应不同位置点处激活的位置细胞,节点之间的连线对应于位置细胞之间的连接关系.机器人在环境中的某个特定位置处,都有对应的获胜位置细胞激活,同时,该获胜位置细胞还记忆了场景图像信息,以便在下次经过时进行有效识别.
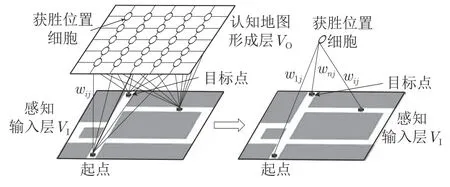
图1 DGP-PCCMM 的“感知—响应”框架Fig.1 The“sense-response”structure of DGP-PCCMM

位置细胞认知地图形成过程中会涉及到一些相关概念,在此提前进行说明如下,并绘制其之间连接关系图如图2 所示.
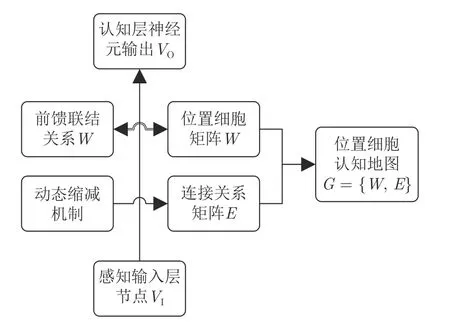
图2 各概念及其之间相互关系Fig.2 Concepts and their associations
1) 认知地图形成层中的神经元不同于位置细胞.机器人每经过一个位置x,都会对应一个能使认知层神经元输出最大的神经元vi(x),vi(x)便是感知层输入x的获胜神经元;获胜神经元会对应一个前馈联结权值wj,wj即为对应输入x的激活的位置细胞.
2) 前馈联结关系矩阵等价于位置细胞矩阵,都用W表示,记录N个激活的位置细胞的位置信息.
3) 连接关系矩阵E记录各位置细胞之间的连通性,在位置细胞增长过程中形成,并能通过动态缩减机制更新.
4) 位置细胞矩阵W和连接关系矩阵E共同组成位置细胞认知地图G.
1.2 构建动态增长的认知地图
为体现机器人与环境交互过程中位置细胞动态增长的特性,基于上述“感知—响应”框架,首先构建以位置细胞为顶点,以位置细胞之间连接关系为边的动态增长认知地图,其构建步骤如下.



动态增长认知地图构建流程如图3 所示.
认知地图与人工神经网络类似,是一个由输入空间到输出空间的映射,即由环境感知到认知地图的映射.模型中,由前馈突触权重向量构成的集合构成映像输出空间的节点,即获得位置细胞矩阵W;并能在此过程中获得位置细胞邻接矩阵E;从而获得认知地图G={W,E}.

图3 动态增长认知地图构建流程图Fig.3 The flow chart of dynamic growing cognitive map
1.3 认知地图的动态缩减机制
为使认知地图模型能够适应动态环境,即在遇到动态障碍物时能动态更新认知地图,我们设计了认知地图的动态缩减机制.此处的动态障碍物是指,在初次访问某位置时不存在,而在第2 次访问相同位置时出现的障碍物.认知地图动态缩减机制的基本原则是:机器人在导航过程中检测到动态障碍物时,将当前位置细胞与即将到达的位置细胞之间的连接关系变为 0,认为两者之间的距离变为无穷大,并对认知地图的连接关系矩阵E进行实时缩减与更新.之后调整机器人位姿至可行方向,应用第2节介绍的位置细胞序列规划算法重新进行序列规划,并指导机器人导航.值得一提的是,由于机器人探测到障碍物时只能观察到距离障碍物表面的距离,并不能得知障碍物的厚度信息,故不能删除即将到达的位置细胞,而只是将其之间的连接关系变为0.
2 基于认知地图的导航
基于位置细胞认知地图的移动机器人路径规划与导航,可分为三大部分内容:定位、构图和决策.当给定一个导航任务后,首先对机器人当前位置进行定位;其次,将感知输入层得到的机器人位置信息输入到认知地图形成层,根据DGP-PCCMM 进行认知地图的构建和更新;接着,利用位置细胞序列规划算法进行行为决策,指导机器人导航.机器人每行进一步,判断是否到达目标场景,如果到达目标,则机器人对认知地图及场景信息进行记忆,导航结束;如果没到达目标,则重新对当前位置和目标位置进行定位,根据认知地图进行行为决策,直至到达目标点,结束导航.机器人导航框图如图4所示,本文主要研究获得精确定位信息后的认知地图构建和行为决策部分的内容,用粗虚线框表示.
为实现基于认知地图的路径规划与导航,提出位置细胞序列规划算法.从动物导航有目标取向性获得启发,定义导航中的负取向函数为当前位置细胞经相邻位置细胞到目标点位置细胞的能量消耗.所经过的路径越长,能量消耗越多,负取向函数值越大.因此在导航时,选择负取向函数值最小的相邻位置细胞作为下一步的运动方向.机器人每到达一个位置,对应一个位置细胞激活,计算一次负取向函数,获得机器人下一步的运动方向,行进一个步长后更新当前位置细胞并将其加入导航路径,如此迭代直到到达目标点位置细胞,最终可以获得从起始点位置细胞到目标点位置细胞的位置细胞序列P,从而实现导航.算法流程图如图5 所示.
3 仿真实验

图4 机器人导航框图Fig.4 The diagram of robot navigation
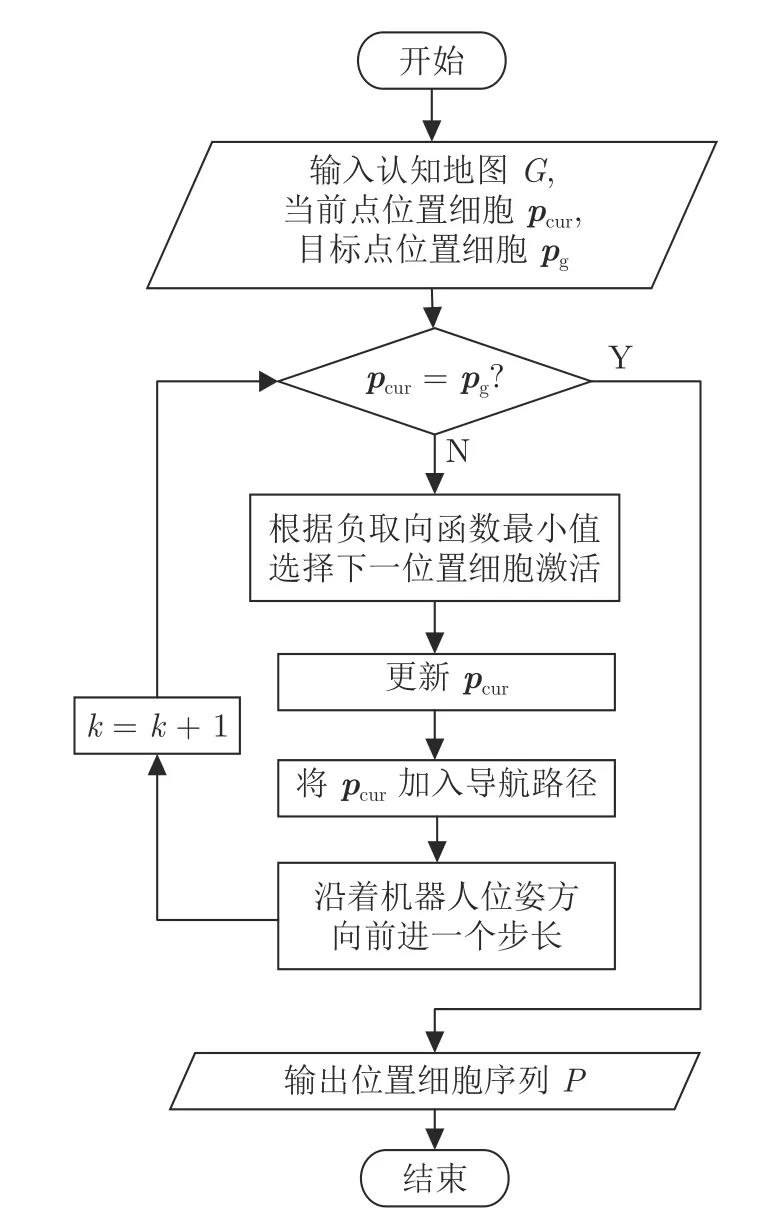
图5 位置细胞序列规划算法Fig.5 The sequence planning algorithm of place cells
检验机器人是否具有智能的一个方法是令其完成与动物相类似的任务,例如用机器人再现老鼠实验.老鼠的绕道取食行为使其能适应环境的变化,提高其存活的几率,且这种绕道取食行为是由后天学习获得.认知模型结果与动物实验结果的契合度对比是评价、判断和比较认知模型的最广泛的方法[25].本文研究了能动态增减的位置细胞认知地图模型 DGP-PCCMM.为验证该学习模型的正确性,本文对Tolman 老鼠迷宫中的绕道实验进行再现,首先对绕道实验进行介绍,之后介绍机器人再现绕道实验仿真环境的设计,最后给出实验结果.
3.1 Tolman 老鼠绕道实验
1930 年,Tolman 设计了三迷宫实验用于研究老鼠的绕道行为智能;1964 年,Maier 团队将Tolman 实验进行简化,本文采用简化后的Tolman 迷宫环境[26],如图6 所示.Tolman 老鼠绕道实验分为两个阶段,环境探索阶段和觅食导航阶段.在环境探索阶段,通过分别在离三叉路口不远的地方设置障碍来迫使老鼠从三条不同路径中选择一条来达到目标,最短路径和中间路径共享一个在最长路径中没有的公共路径.在觅食导航阶段,老鼠会首先选择通往目标点的最短路径Route 1;当发现A处障碍后,老鼠会选择中间路径Route 2;当发现在最短和中间路径的公共部分有障碍即障碍B时,它们会选择最长的路径Route 3,而不是选择中间路径,从而证明它们知道被阻塞的部分同时属于最短路径和中间路径.
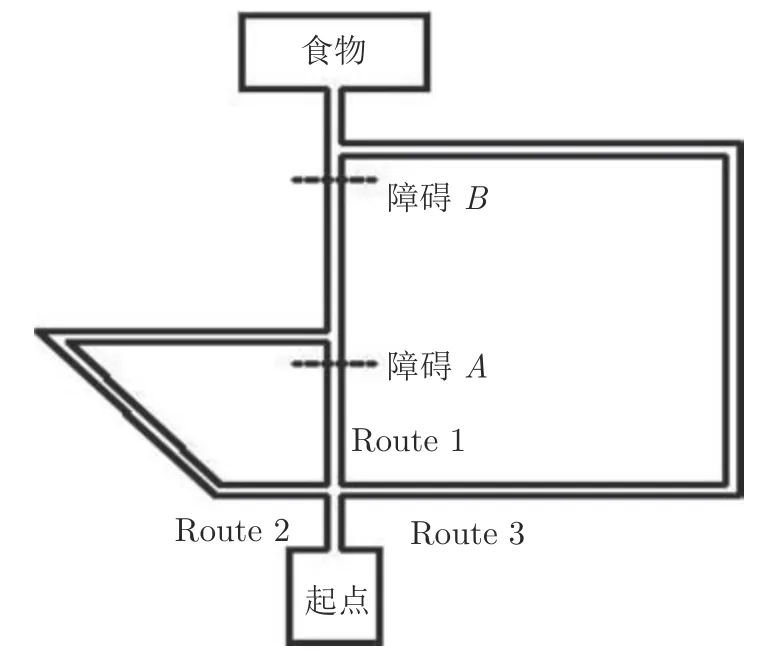
图6 Tolman 老鼠绕道实验迷宫环境Fig.6 Maze environment of Tolman detour task
Tolman 认为在老鼠大脑中存在认知地图,当在迷宫中寻找目标时,即使在迷宫中的任意点看不到全局环境,但能根据认知地图进行行为决策,选择最优路径进行导航.
3.2 实验设计与结果
本文实验对象是轮式圆形机器人,半径为2.5 cm,运动线速度v=2.5 cm/s,机器人通过左右两轮WL和WR的差动驱动行走,通过改变方向轮WD朝向可以实现任意角度的转向,其俯视示意图如图7 所示.从老鼠感知系统获得启发,本文假设可以获得机器人精确的位置信息,作为认知地图形成层VO的输入;配备8 个测距传感器均匀分布在圆形机身用于探测与障碍物之间的距离,测距范围为 1~ 10 cm.本文仿真环境尺寸参考文献[22]中的Tolman 迷宫环境,尺寸为 130 cm×130 cm,其中灰色代表障碍物,白色代表通路,通路宽度为 10 cm,如图8 所示.

图7 轮式圆形机器人俯视图Fig.7 Top view of wheeled circular robot
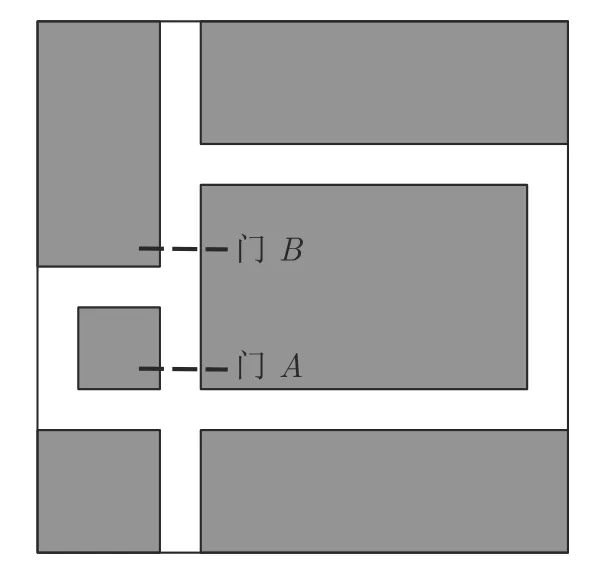
图8 Tolman 迷宫仿真环境Fig.8 The simulation environment of Tolman maze
实验 1.认知地图构建过程
首先让机器人漫游环境,以(35,5)为起点,每经过一个步长都采集位置信息、场景信息和障碍信息,边漫游边构建认知地图.考虑到机器人自身尺寸,在仿真实验采集样本点的过程中,首先将墙边界进行膨胀化处理,机器人所处的位置中心离边界的距离一定大于机器人的半径,此处设置为3 cm.设置机器人随机搜索的行进步长dstep为 5 cm.初始实验参数设置见表1.
分别采集1 000 和2 000 个能遍历整个迷宫环境的位置点及相应的场景记忆信息,每有25 个位置细胞激活就记录一次认知地图,可以得到以下的认知地图的构建过程,分别展示于图9 和图10.图中圆点为机器人所经过的位置记录,星号表示生成的位置细胞.机器人刚开始只有一个位置细胞激活,随着机器人探索环境获得可达位置点的增多,所激活的位置细胞也逐渐增多,最终只用了少量的位置细胞就可以映射整个迷宫环境.如图9和图10 所示,当有1 000 个可达位置点时,共有90 个位置细胞激活过;当有2 000 个可达位置点时,共有93 个位置细胞激活过.机器人在随机探索环境的过程中,所激活的位置细胞个数不断增多,表明其对环境的认知程度逐渐增强,最终能自组织地形成整体环境的认知地图.
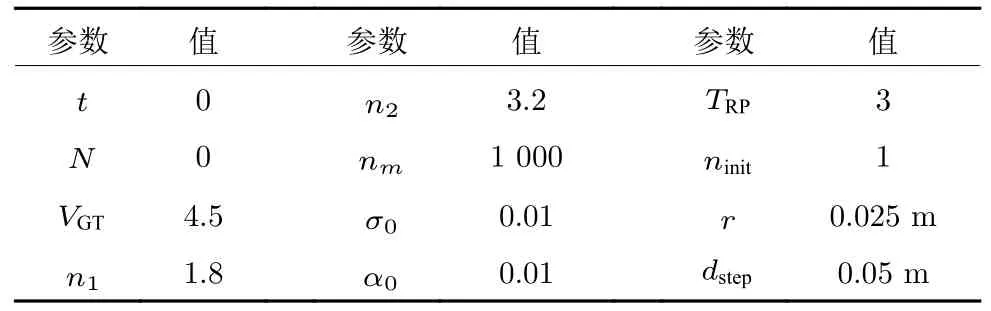
表1 DGP-PCCMM 初始参数设置Table 1 Initial simulation parameters for DGP-PCCMM
图11 展示了位置细胞个数N随学习次数n的变化情况,图例中nm表示最大学习次数.实线和点划线表示学习次数分别为1 000 和2 000 时,位置细胞个数的变化.由图11 可知,在初始阶段只有一个位置细胞激活;随着与环境交互次数的增多,激活过的位置细胞个数不断增加;当机器人基本遍历环境后,认知地图中的激活过的位置细胞个数大体一致;之后继续遍历环境,位置细胞的个数变化不大.值得一提的是,由于老鼠在迷宫中属于探索式运动,经常出现折返现象,故对环境的认知也是渐进的.
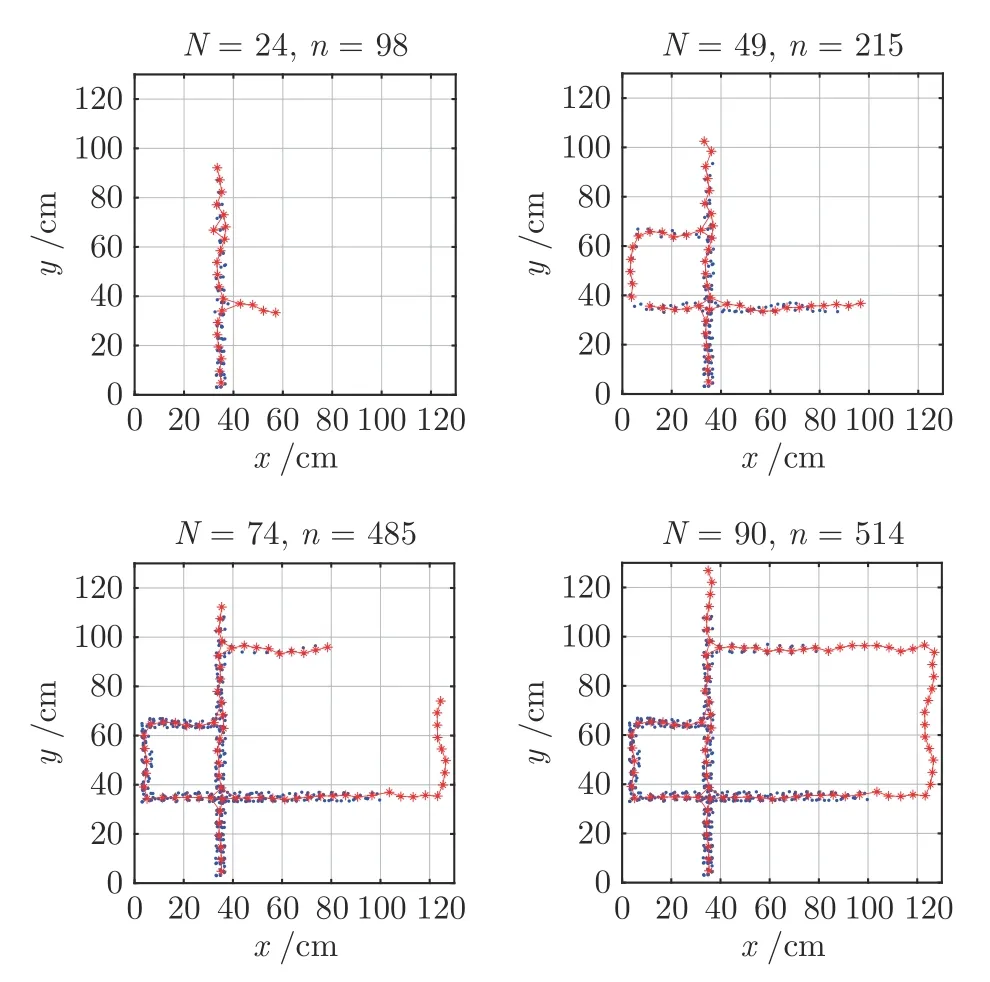
图9 认知地图构建过程(nm=1 000)Fig.9 The formation process of cognitive map(nm=1 000)

图10 认知地图构建过程(nm=2 000)Fig.10 The formation process of cognitive map(nm=2 000)
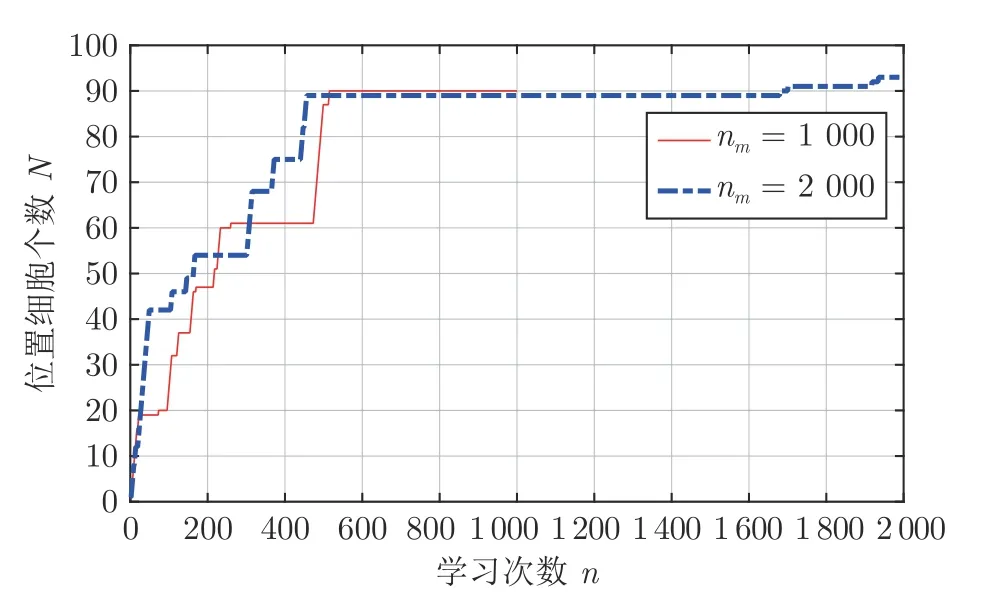
图11 位置细胞个数随学习次数变化情况Fig.11 The number of place cells changing with the number of learning times
实验 2.再现Tolman 老鼠绕道实验
上述实验阐明基于位置细胞的认知地图是如何自组织构建出来的.在这个实验中,我们使用已构建好的认知地图进行Tolman 绕道实验的再现,所用迷宫环境如图8.起始点设置在(35,5),目标点设置在(35,125).实验分为三部分,首先进行门A和B都打开情况下的导航,其次是门A关闭门B打开情况下的导航,最后是门A打开而门B关闭情况下的绕道行为再现.
首先在起始点根据位置细胞序列规划算法得到一条初始全局最优位置细胞序列,机器人根据所规划的位置细胞序列向目标点位置细胞行进;在行进过程中,当环境感知模块检测到当前场景与记忆中的场景信息不匹配时,启动认知地图动态缩减机制,更新位置细胞之间的连接关系,得到更新后的认知地图;调整机器人位姿,重新进行位置细胞序列规划选择行进方向,最终到达目标点.
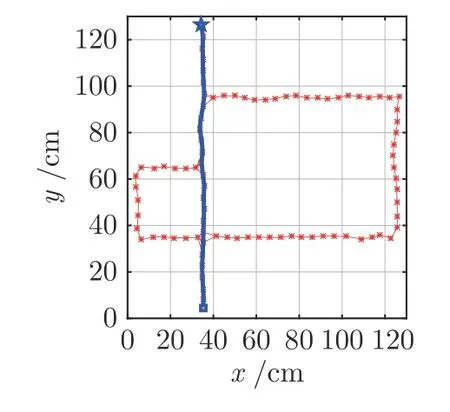
图12 门A 和门B 都打开情况下的导航Fig.12 Navigation with door A and door B open
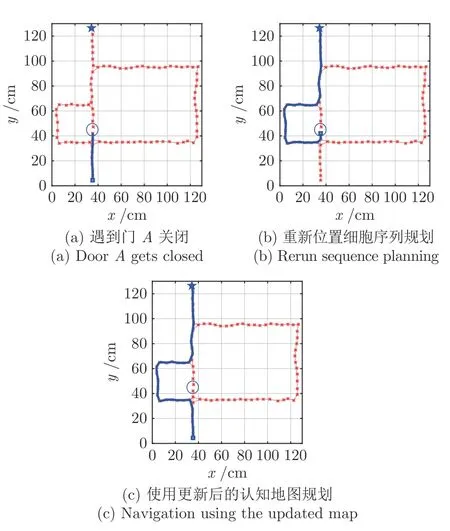
图13 门A 关闭门B 打开情况下的导航Fig.13 Navigation with door A closed and door B open
图12~ 14 为再现Tolman 绕道实验结果图,其中,星号表示激活的位置细胞,星号及其之间的细线表示得到的认知地图,方块表示起点,五角星表示目标点,粗线表示根据位置细胞规划算法得到的导航路径.图12 为门A和B都打开的情况,此时机器人选取最短路径Route1 作为导航路径.图13为门A关闭而门B打开的情况,开始时机器人在起始点继续沿着Route 1 前进,当检测到A处障碍物,启动认知地图动态缩减机制,将当前位置细胞与下一位置细胞之间的连接关系设为0;机器人位姿调转180°方向,重新进行位置细胞序列规划,可以看到机器人选择中间路径到达目标点;在下次导航时,由于认知地图已经更新,机器人直接选择Route 2到达目标点.同理,当门A打开而门B关闭时,最终机器人将学习到选择Route 3 进行导航,如图14 所示.
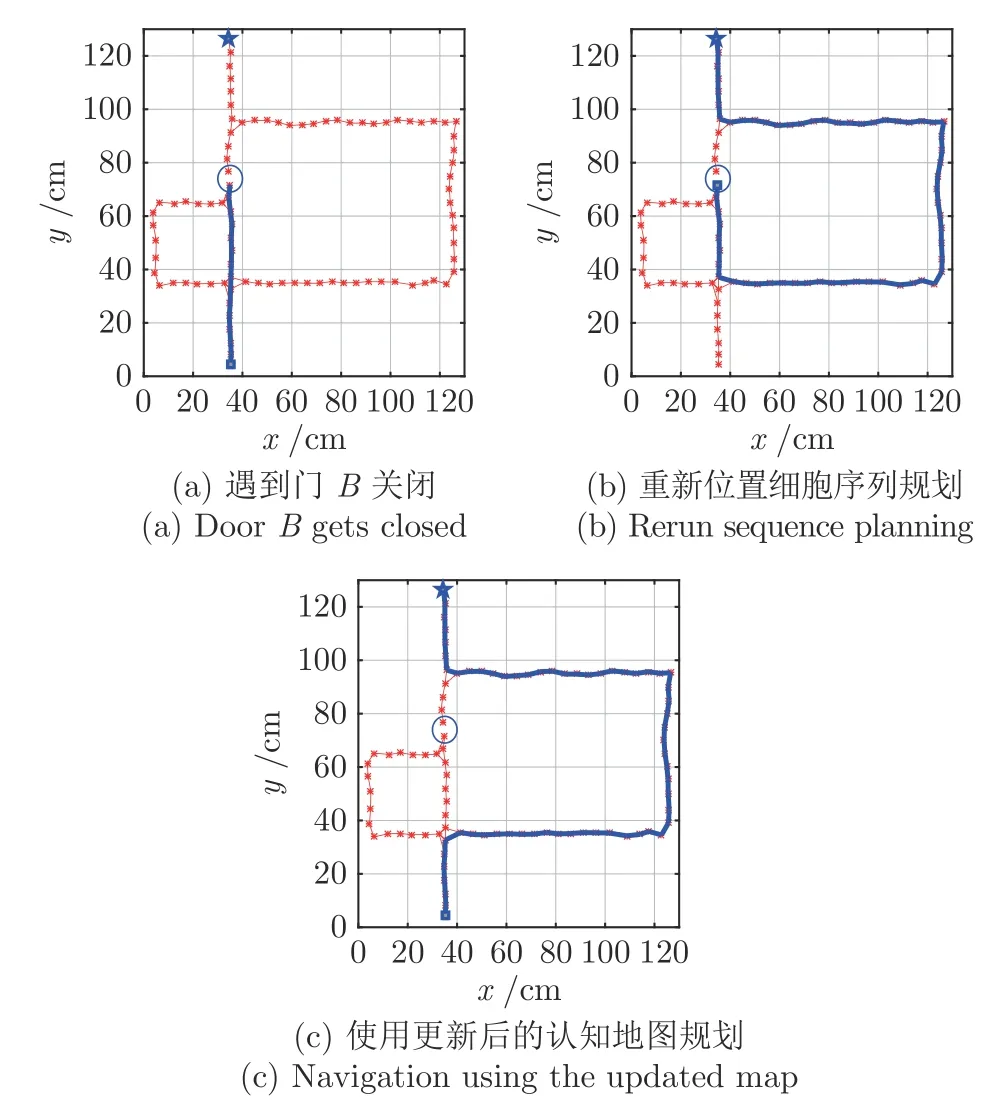
图14 门A 打开门B 关闭情况下的导航Fig.14 Navigation with door A open and door B closed
仿真结果表明,移动机器人能够在与环境交互的过程中感知环境并在遇到动态障碍物时动态更新认知地图;同时基于所构建的位置细胞认知地图,利用位置细胞序列规划可以获得最优导航路径,从而实现对Tolman 绕道实验的再现.
4 讨论
机器人建图一直是机器人和人工智能领域的一个非常活跃的研究领域,主要解决通过移动机器人获取物理环境空间模型的问题.在20 世纪80 年代到90 年代早期,机器人建图主要分为栅格地图和拓扑地图两种方法.栅格地图采集环境的几何特性,而拓扑地图侧重描述不同位置之间的连接[27].同步定位与地图构建(Simultaneous localization and mapping,SLAM)问题从1986 年提出后逐渐兴起[28],侧重于工程实践研究.之后,随着位置细胞、网格细胞、头朝向细胞等与环境认知和导航相关细胞的发现,仿生建图研究逐渐成为新的研究潮流,主要以探索动物环境认知机理和构建更智能的移动机器人为目的[15,29],侧重于仿生理论研究.本文属于仿生建图研究的范畴.下面将本文的认知地图与几种经典的建图方法进行对比讨论分析.
4.1 与栅格地图对比讨论
栅格地图是常用的地图表示方法,其优点在于建模简单、易于更新和处理,但随着地图范围扩大,所需要的存储空间将会大大提升.近年来,自适应栅格地图表示方法被提出,能根据环境自适应调整栅格大小,从而有效节省存储空间,如基于四叉树的自适应栅格地图创建方法.然而,自适应栅格地图仅适用于静态环境,在障碍物密度差异大的环境中效果明显;在障碍密度差异小且障碍物较多的环境中,其优势不突出[30].本文模型不仅具有类脑仿生特性和生物合理性,而且经实验验证,在所构建地图的简洁性和对环境中动态障碍的适应性方面更有优势.
1) 在所构建地图简洁性方面的对比
为进行对比分析,本文分别采用四叉树自适应栅格建图法和本文方法对Hairpin 迷宫进行建图.Hairpin 迷宫为如图15 所示的 70 cm×70 cm 的静态室内环境.在标准的四叉树栅格建图中,环境地图分割方法是将整个未知环境切分成4 个节点树,确定子栅格中哪些状态是空置或被占据的,哪些状态是未知的,然后将未知栅格进一步划分为4 个小节点树,指导整个地图搜索完毕并满足精度要求,则建图完成.基于四叉树的栅格地图在一定程度上解决了传统栅格占用海量存储空间的问题,但该方法对应用环境有要求,在障碍密度差异小且障碍物较多的环境中,优势不突出[30].本文认为,Tolman迷宫属于障碍密度差异大且障碍物较多的环境,Hair pin 迷宫属于障碍密度差异小且障碍物较多的环境.

图15 Hairpin 迷宫Fig.15 Hairpin maze

图16 四叉树栅格地图Fig.16 Occupancy grids based on quadtree
图16 是构建的四叉树栅格地图,图16(a)和图16(b)分别为采用四叉树栅格建图法对Tolman迷宫和Hairpin 迷宫进行建图的结果.图17 是本文方法构建的认知地图,图17(a)和图17(b)分别为采用本文方法对Tolman 迷宫和Hairpin 迷宫进行建图的结果.从仿真结果图可以定性地看出,本文认知地图构建整个环境所需要的位置细胞数比四叉树栅格地图所需要的栅格数要少.对实验结果做定量分析,以四叉树栅格建图法形成的大栅格计数,构建Tolman迷宫地图需要的栅格数大于196,构建Hairpin 迷宫地图需要的栅格数大于256.而以本文认知地图建图法构建Tolman 迷宫仅需要90个位置细胞,构建Hairpin 迷宫地图仅需要65 个位置细胞,比四叉树栅格地图更简洁.四叉树栅格地图和本文认知地图的性能指标对比如表2 所示.此处需要说明的是,Tolman 迷宫地图的尺寸为130 cm×130 cm,而Hairpin 迷宫地图的尺寸是70 cm×70 cm,所以构建Tolman 迷宫所形成的位置细胞个数比构建Hairpin 迷宫多.此外,本文建图过程与海马体的一次学习或快速学习机制[31]相吻合,本文地图生成过程更符合动物对环境的认知过程.
2) 在对动态障碍适应性方面的对比
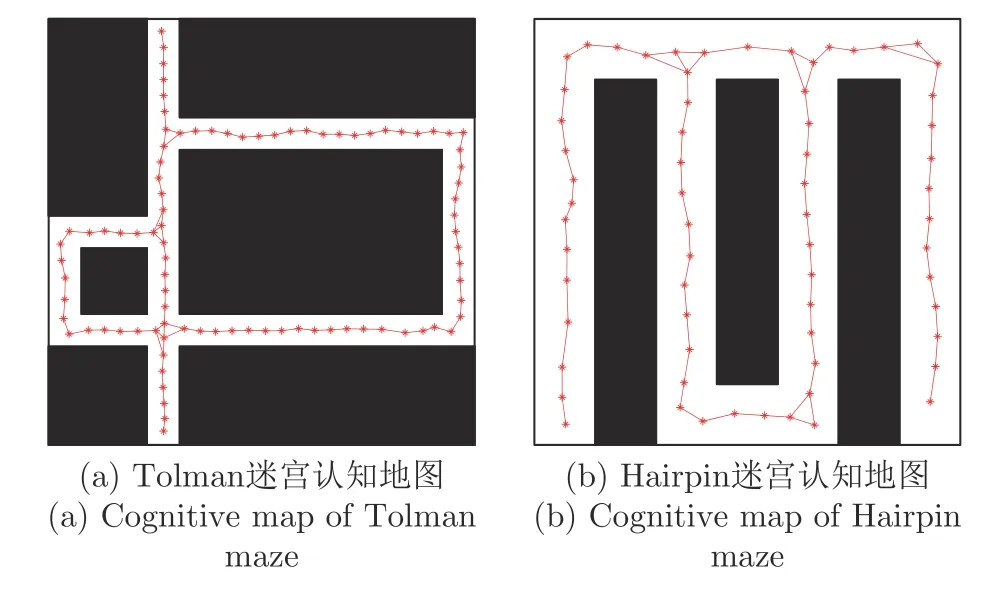
图17 本文方法构建的认知地图Fig.17 Cognitive maps based on methods of this paper

表2 本文与四叉树栅格对比Table 2 Comparison between occupancy grids based on quadtree and DGP-PCCMM
对动态障碍的适应性是机器人环境认知的重要内容.文献[30]表明,四叉树栅格地图法较适合于对静态环境的建图,如果环境发生变化,则需要重新进行四叉树栅格建图,对动态变化的适应性较弱.人工势场法、遗传算法、蚁群算法、动态窗口法等都有一定的局部避障能力[32-33].其中,动态窗口法是近年来较常用的一种局部路径规划方法,对动态变化有良好的适应性,能够实时避障,但其不能有效利用学习到的先验信息,导致耗时较长,且其无法考虑路径的全局最优性,时常会陷入局部最优[33-34].本文方法能在与环境的交互中学习到环境的地图信息,并在之后的导航中能利用学习到的认知地图指导导航,耗时短且能实现全局最优.为对比动态窗口法和本文方法对动态障碍的适应性,将这两种方法分别用于在Tolman 迷宫的导航实验,并在图18中展示两者的导航结果,其中实线和虚线分别表示动态窗口法和本文方法得到的路线.在18(c)门A打开而门B关闭的情况下,动态窗口法会由于避障而出现绕远路或陷入局部最优的现象,并且在多次导航后并不会学习到环境的地图信息,导致每次导航都会绕远路.而在本文方法中,机器人在第一次发现临时障碍后,会及时更新认知地图,从而在之后的导航中能采用规划出的全局最优路径.两种方法的性能指标对比表如表3 所示,其中,T1、T2 和T3 分别表示图18(a)、18(b)和18(c)的情况.由表3 可知,在T1 和T2 情况下,两种方法都能近似最优地进行导航,本文由于根据认知地图进行导航,运行时间相对较短.在T3 情况下,采用动态窗口法由于绕远路和陷入局部最优,运行时间相对很长,T3 的运行时间比T1 增长了约5 倍,而采用本文方法运行时间只比T1 时增长了约2.5 倍.本文认知地图构建方法的一大优点在于在遇到动态障碍时可以利用已有的认知地图,从而在下次导航时直接使用学习后的认知地图进行导航,避免了重新学习环境信息的时间浪费.
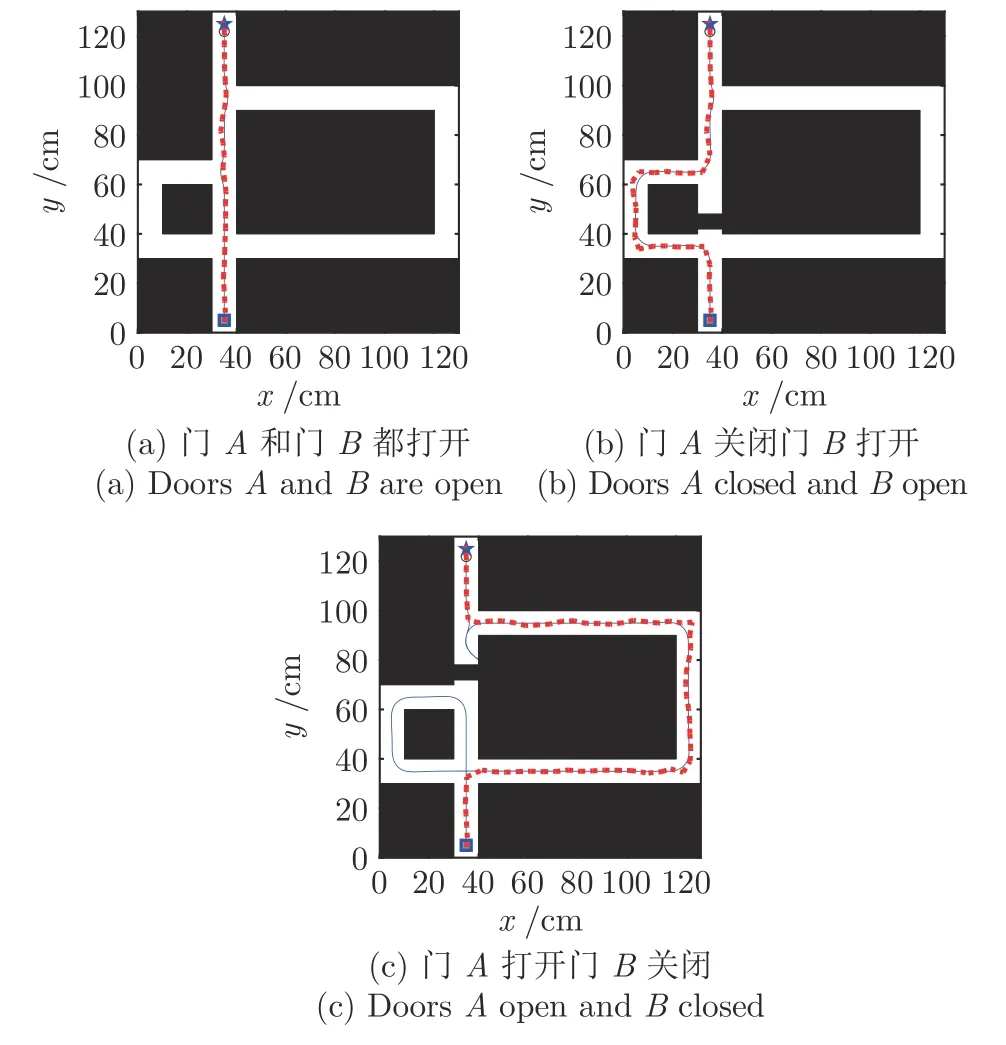
图18 动态窗口法和本文方法导航结果对比图Fig.18 Comparation of navigation results between dynamic window approach and DGP-PCCMM
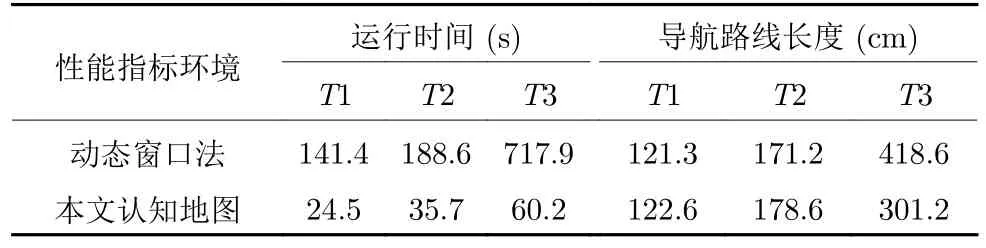
表3 本文与动态窗口法对比Table 3 Comparison between dynamic window approach and our method
4.2 与其他认知地图对比讨论
Martinet 等[20]提出了前额皮质柱神经网络模型,通过构建位置细胞和皮质柱的分层网络再现Tolman 绕道实验.文献[20]和本文都能对绕道实验进行再现,文献[20]展示了皮质柱网络结构从海马输入中学习稀疏的拓扑度量表示,而本文侧重基于海马体位置细胞进行认知地图的构建与更新,属于对老鼠空间导航机制的不同研究方向的探索.
Erdem 等[35]提出了基于前向轨迹探针的目标导向的空间导航模型.通过随机探索,可以创建由位置细胞和前额皮质细胞组成的地图,实现目标导向的行为.然而,正如文献[35]的讨论中所说,何时引入位置细胞来表达空间环境的位置信息是一个重要的问题,文献[35]中的位置细胞是通过特定的伪随机方法来产生的,存在不能很好表达环境的问题.本文采用类似自组织图的“感知—响应”结构,通过竞争与动态增长获得位置细胞,可以实现对环境的良好表达,两种方法获得的认知地图如图19 所示,图19(a)为文献[35]得到的认知地图,其中圆圈表示位置细胞,虚线表示得到的导航路径;图19(b)为本文方法得到的认知地图,其中星号表示位置细胞,星号及其之间的连线表示得到的认知地图.通过图19 可知,文献[35]得到的认知地图在某些位置处没有位置细胞表示,如最左边一列,而在某些位置处有多余的位置细胞表示,如从左边数第三列可行通路,不能生成对整个环境的完整表达;相比较而言,本文方法可以获得对整个环境的一个均匀完整的表达,很好地解决了文献[35]提出的如何引入位置细胞来表达空间环境这一重要问题.
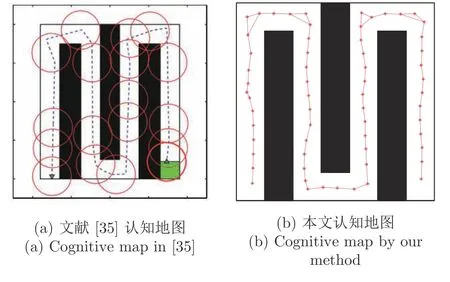
图19 Erdem 认知地图与本文认知地图对比Fig.19 Comparison between different cognitive maps
概括来说,本文认知地图构建方法有以下优点:1) 具有类脑仿生特性与一定的生物合理性.与栅格地图和一般的拓扑地图相比,本文认知地图的构建和更新过程与海马体的一次学习或快速学习机制相一致,更符合动物对环境的认知过程.2) 能够适应动态环境(如门的开闭).与动态窗口法相比,在遇到动态障碍后,本文方法能动态更新认知地图,并利用学习到的认知地图指导导航,耗时短且能实现全局最优.3) 能够降低建图的复杂性,并通过构建Tolman 迷宫、Hairpin 迷宫地图进行验证.与四叉树栅格地图相比,本文方法不仅适用于障碍密度差异大的环境,而且在障碍分布差异小且障碍物较多的环境中也表现出较好的性能.
5 结论
本文用机器人再现动物绕道实验,是通过仿生使机器人更加智能化的一个初步探索.本文受海马体位置细胞在特定位置放电的启发,通过构建动态增减的位置细胞认知地图,再现了Tolman 老鼠绕道实验.目前研究的重点集中在对认知地图的构建上,所提出的构建方法具有以下优点:1) 具有类脑仿生特性与一定的生物合理性;2) 能够适应动态环境(如门的开闭);3) 能够降低建图的复杂性.实验结果表明,机器人漫游和导航的过程中能动态形成和更新认知地图,能像Tolman 老鼠那样,利用认知地图进行自主环境探索、认知与导航,完成绕道取食.
本文属于对动物导航过程中大脑运作机制的一个探索,对海马体位置细胞的建模是后续研究动物导航过程中可能涉及到的内嗅皮层、丘脑、纹状体等之间联合建模的基础.本文简单表明了海马体位置细胞的场景记忆功能,对机器人场景记忆的仿生建模有待进一步的研究.将来期望进一步完善机器人对Tolman 绕道实验的再现,使机器人实现更仿生(如将障碍物撤除后机器人重新发现捷径等)的环境认知.
猜你喜欢
科技创新与应用(2021年31期)2021-11-09
小猕猴智力画刊(2019年9期)2019-11-08
文理导航·科普童话(2016年7期)2017-02-04
学苑创造·A版(2016年9期)2016-10-10
文理导航·科普童话(2016年4期)2016-05-31
儿童故事画报·智力大王(2015年12期)2016-01-23
儿童故事画报·智力大王(2015年2期)2015-05-20
弹箭与制导学报(2015年1期)2015-03-11
雷达学报(2014年4期)2014-04-23
哈尔滨工程大学学报(2013年5期)2013-06-05
