
山地地盘松种群间隔10年的环境适应表现对比*
2021-04-22 13:42:46蔺雨阳张帆肖翠杨涛雍小华张春花
西部林业科学 2021年2期
蔺雨阳,张帆 ,肖翠,杨涛,雍小华 ,张春花
(1.四川省攀枝花市农林科学研究院,四川 攀枝花 617000;2.四川外国语大学翻译学院,重庆 401120;3.陕西省林业科学院,陕西 西安 710082;4.四川省攀枝花市盐边生态环境局,四川 攀枝花 617000)
地盘松(Pinusyunnanensisvar.pygmaea)是云南松(P.yunnanensis)由于经特殊的山地环境筛(主要是干旱贫瘠的土壤、屡遭火烧、砍伐和放牧等)的过滤,植株适应环境并协同进化所致,是环境对杂合性种群起直接选择作用而产生的生态小种[1],是种群的多态现象之一[2]。目前云南松生态小种或地理小种的研究主要集中在变种间的对比研究[1-8],侧重于工业用材质量和特征的定性描述研究[2,5-7],对回答生态小种特别成因问题显得不够深入[8-10],在探讨环境适应表现等生态过程和作用机制方面亟待更进一步研究[11-12]。
植物功能性状是植物与其定居、存活、生长和死亡等生态过程联系的系列核心植物属性[13-15],反映了植被对环境变化的响应[16-18]。近年来,随着对功能性状研究的深入,生态学家们从植物功能性状角度出发可以简明扼要地解释一些生态现象[18-19]。功能性状的改变包括形态可塑性、生理可塑性和寿命可塑性等[13-14]。地盘松功能性状的形态可塑性是研究其环境适应策略的重要方面。地盘松作为生态小种,正是通过形态特征的可塑性,表达植物的环境适应策略[15]。蔡年辉等[16]也认为应通过跟踪观察来验证其物理变形,从而探讨在时序上的性状塑造性。参照林业行业标准《LY/T 2812—2017 森林木本植物功能性状测定方法》中性状指标设置原则,由于地盘松干枝特殊的着生关系和盘曲的树体结构形态,设置合适的性状指标非常重要[17]。此外,种群的空间格局也在很大程度上反映了长期的环境适应结果,决定了建群种发展的可能性和经营空间的大小[20],对于帮助揭示环境适应策略多维属性具有重要意义。因而从植物的功能性状和空间格局两个途径入手研究植物的环境适应性策略可以互相补充。
如何选择性状指标反应植物形态可塑性是非常关键的[21]。地盘松地上部分形态不同于其它松类植物生长形态[2],常规的胸径DBH、树高H、冠幅C等性状参数,不能有效反映特殊的着生位置和盘曲或匍匐的树体结构形态。再者“空间代替时间”的方法研究种群的空间格局会引起种群密度低估的现象,不能真实反映种群空间格局随长时间变迁的演变特征。因此,本文通过创造性地设置4个形态参数,来解析地盘松的形态可塑性,进而对比分析2019年与2009年山地地盘松形态性状和空间分布格局的变化,通过环境适应表现剖析其环境适应策略。
1 研究区概况
攀西大裂谷位于云贵川高原横断山脉抬升的地质地貌构造上,是典型的中国西南喀斯特中等发育的区域。这里有着世界上湿润-半湿润、热带-亚热带地貌发育和植被的主要特征[22-24]。本研究区域在川西南攀西大裂谷九道竹林区,地理坐标101°07′57″~101°25′54″E,26°53′30″~27°12′40″N,海拔1 500~3 200 m。土壤主要以山地暗棕壤为主,棕壤和红棕壤也有分布,因枯落物等有机质归还少,腐殖质层瘠薄,淋溶作用强烈,有较为典型的发育中等的喀斯特地貌特征。地形的复杂导致了复杂的小气候特征,多湿冷气候,但总体干湿季节分明,蒸发量大于降雨量。年平均温度13~19 ℃,最热月平均温度达到20 ℃。根据当地气象资料,近10 a气候相对稳定,无明显异变的气象记录(盐边县气象局就近气象观测站提供)。该区域分布有杜鹃(Rhododendronspp.)林、地盘松林和亚高山草甸的多种植物群落[22]。地盘松种群在该研究区明显占优,分布范围和种群数量明显占优,有“万亩盘松”之称。地盘松为建群种的群落中伴生树种主要为高山栲(Castanopsisdelavayi)、滇青冈(Cyclobalanopsisglaucoides)、滇石栎(Lithocarpusdealbatus)等。林下活地被物不发达,下木和草本层多见小铁仔(Myrsinestolonifera)、厚皮香(Ternstroemiagymnanthera)、野拔子(Elasholtziarugulanus)、野青茅(Deyeuxiaarundinacea)、火绒草(Leontopodiumspp.)等。
2 研究方法
2.1 样地设置与调查
2009年在四川省盐边县格萨拉生态旅游区西北九道竹林区地盘松集中分布区开展调查。鉴于地盘松集中分布在高差约200 m的大面积平缓山地上,采用样带法进行抽样调查。沿着海拔梯度每100 m设置一个样带,样带大小50 m×20 m,样带方向与等高线垂直,共设置3个。各样带中以相邻网格法划分为10 m×10 m样方进行每木调查。调查面积3 000 m2,获得地盘松样株信息916株。采集土壤和气候数据,记录其地形地貌信息和采集样方中木本的每木调查(包括地径、冠幅、树高、树干全长,节间长,轮生枝存数,每木相对坐标)数据。地盘松的每木轮生枝存数调查包括活轮生枝存数和枯轮生枝存数,枯轮生枝的记录距树冠长度;地盘松的弯曲或扭曲或匐地的树干全长采用软绳拟合树干弯曲形状后测量取得,分别记录;每木相对坐标值测定时,在样带中纵横每间隔10 m放置铺伸的50 m卷尺一把,测量者持5 m卷尺移动测定每木的相对坐标位置(X,Y),测定一株标注一株,防止重计或漏计。坐标值精确到0.1 m。
2019年,在原调查样带和样方原址的山体上进行新的一期调查,调查方法与原方法相同。调查面积3 000 m2,获得地盘松样株信息859株。样地(样带)信息见表1。

表1 地盘松样地(样带)信息
2.2 形态参数的设置
参考《LY/T2812—2017森林木本植物功能性状测定方法》,充分考虑到部分常规的植物功能性状不能有效揭示地盘松特殊的着生关系和盘曲的结构形态的实际,需要根据研究重点重新构造表型形态指标。针对地盘松的弯曲或扭曲或匍匐的特征,设置表征地盘松个体的弯曲、扭曲程度,铺地占据空间的能力,轮生枝节间长连年增长量。故而创造性的设置如下4个形态参数。
(1)树曲高比数(ratio of height and length,HLR)
是一个无量纲数值,树体弯曲或扭曲或匍匐树干全长和树高的比值,用于表达地盘松扭曲程度,数值越高树体越扭曲变形。计算公式如下。
RHLR=L/H
①
式中:L为树体弯曲或扭曲或匍匐树干全长(m),H为树高(m)。
HLR特点 综合了L和H 2个直接测得的数据所表达的信息,体现了两者的制约关系,相比工程学参数曲率半径表达弯曲程度要更加直接和简易。
(2)幅径截面比数(cross-sectional area ratio of canopy and ground diameter,CARCD)
是一个无量纲数值,冠幅与地径截面积的比值,用于表达地盘松经历逆境表现的生长势,数值越小表示其生境越差,生长越艰难。计算公式如下。
RCARCD=ACanopy/ADiameter=(π·aCanopy·bCanopy)/[π·(1/2·DDiameter)2]
②
式中:ACanopy为树冠截面积(按照椭圆面积计算近似值);ADiameter为地径截面积;DCanopy、DDiameter分别为冠幅与地径的直径;aCanopy、bCanopy分别为冠幅的长轴和短轴。
RCARCD特点 指标中冠幅截面积椭圆法计算可以有效杜绝因铺地程度不同造成冠幅的椭圆偏心率各异进而引入冠幅被过高估的误差;同时,与地径截面积的比值可以排除在地盘松为追逐光照等资源造成树体徒长的情况下生长势高估问题。
(3)活枝存数干高比(ratio of the number of living branches to the difference between the total length of the bent or twisted or creeping trunk and the height of the tree ,RNDLH)
轮生枝活存数和树体弯曲或扭曲或匍匐树干全长与树高之差的比值,用于估计地盘松在变形特征下自然树冠竖向丰度,也侧面反应立地的严酷程度。计算公式如下。
RRNDLH=N/(L-H)
③
式中:N为轮生枝活存数目;L和H同公式①。
RRNDLH特点 弥补了常规调查树冠丰度时忽略竖向丰度的缺陷,并且着重考虑了地盘松特殊的树体形态下特有的竖向丰度信息载有量。
(4)轮生枝间长均数(mean of length between living branches,MLB)
是指树体最近10个(因考察研究对象近10 a的生长量)存在的主干顶枝轮生枝间长的平均值,用于表达地盘松10 a间生长量情况。
MLB特点:在时间跨度上考察连年生长量可以在经历10 a跨度后的轮生枝间变化,用以评估种群轮生枝的环境适应变化情况。避免了当年轮生枝间长带来的数据单一和异常值的干扰。
2.3 形态对比
为了比较2009年和2019年间地盘松形态可塑性,采用方差分析检验了上述形态参数10 a前后的差异。为区分不同树龄个体的差异,按固定径级为序列统计为6组(地径级以3 cm为阶分级编树龄组,树龄组以1至6编号为序)以代替自幼到老的林龄组序列。为更好表述,下文中每两树龄组为龄段(3个生长阶段[23]),即依次为幼林龄段(sapling stage)、中林龄段(juvenile stage)、成熟林龄段(mature stage)[20,23],进一步进行差异检验。
2.4 点格局分析
采用点格局分析法O-ring函数[25],O-ring统计用圆环取代了传统的Ripley’s K函数在进行空间格局分析中使用的圆圈,计算环内点的平均数目,从而孤立了特殊的距离等级[13]。计算公式如下。
④

本研究选择不校正边界效应的函数,采用矩形(analyze data in rectangle)方法研究,空间尺度为0~50 m,步长为 1 m,经 199次Monte Carlo 模拟得到 99% 的置信区间。关于包迹线区间的判定参考董灵波等[27]对点格局的研究方法。
2.5 数据处理
使用SPSS 26.0进行箱线图制作、相关分析和方差分析(AVAVON)。点格局分析使用Diggle等开发的格局分析软件Programita[29]进行,导出结果后的制图在Excel 2013中完成。
3 结果与分析
3.1 形态特征参数的比较
为了弄清楚2009年和2019年间形态特征参数的情况,使用方差分析比较。从表2看出,4个不同的功能性状参数在两次调查时的差异除RNDLH外均达到了极显著水平。经过10 a时间的地盘松种群迁变,前后对比地盘松种群的功能性状属性确实发生了较大的差异。
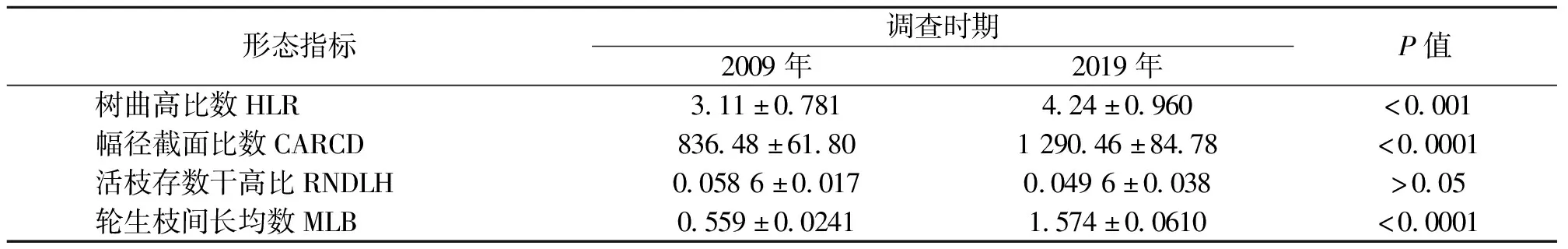
表2 10年前后两期地盘松不同形态指标的对比结果
3.1.1 树曲高比数Highth Length Ratio(HLR)
对地盘松种群个体的HLR进行比较,结果如图1所示。2019年普遍高于2009年。同时在中林龄段的种群,HLR没有明显分异,而在幼林龄段的种群和成熟林龄段的种群的HLR有明显的分异,2019年的HLR明显居高,也就是云南松树体有更加明显的扭曲、弯曲变形。

图1 2019年和2009年在不同林龄组序列上的HLR
3.1.2 幅径截面比数CARCD
同样以地盘松种群的林龄组序列进行统计并比较幅径截面比数CARCD,结果如图2所示。2019年普遍高于2009年。在成熟林龄段的种群,CARCD没有明显分异,而在幼林龄段的种群和中林龄段的种群CARCD有较为明显的分异,2019年明显居高,也就是地盘松种群2019年树体树冠更为丰腴。
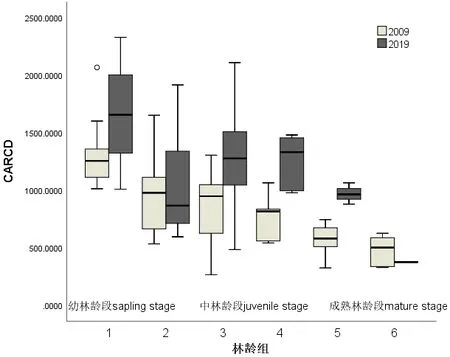
图2 2019年和2009年在林龄组序列上的CARCD
3.1.3 活枝存数干高比RNDLH
如图3所示,在幼林龄段种群的RNDLH有较明显分异(P<0.05),成熟林龄段的种群显著差异(P<0.01)。而在中林龄段的种群的RNDLH没有明显的分异(P>0.05)。中林龄段的种群的树冠在竖向的丰满程度,在经历了10 a的变迁并没有发生明显的变化。
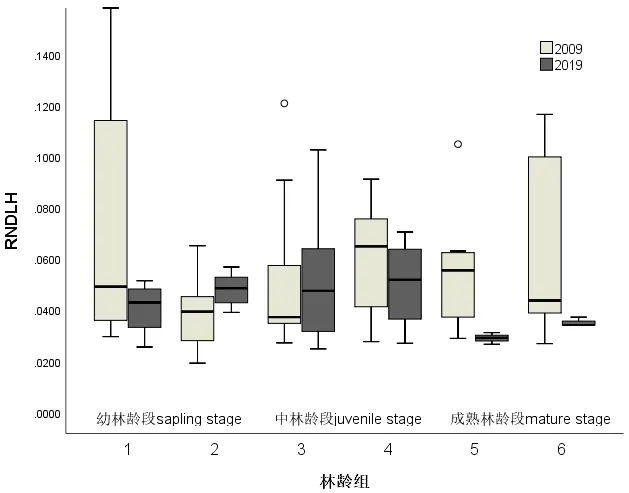
图3 2019年和2009年在林龄组序列上的RNDLH
3.1.4 轮生枝间长均数MLB
2009年的MLB普遍小于2019年(P<0.001)(表2,图4)。在中林龄段的种群,MLB明显分异(P<0.001),而在幼林龄段的种群和成熟林龄段的种群的MLB也明显的分异(P<0.001),2019年的MLB较之2009年的明显徒长。
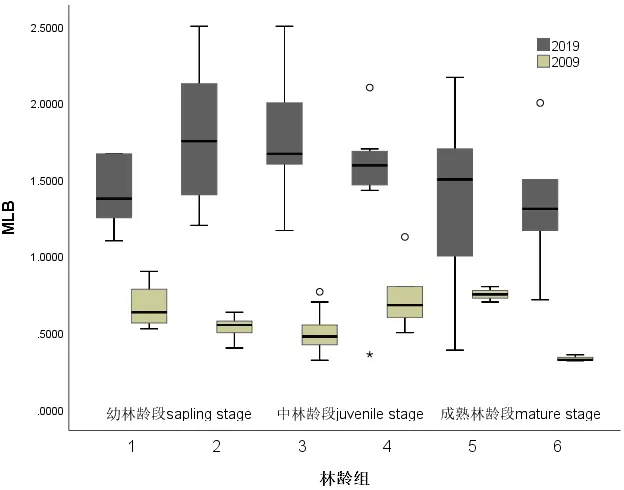
图4 2019年和2009年在林龄组序列上的MLB
3.2 种群点格局特征的比较分析
两期的散点图初测在研究尺度上无明显的聚集分布特征,因此运用O-ring函数中的异质性泊松过程(HP)为零假设模型。
在1~50 m的尺度范围内,地盘松种群主要表现为随机分布格局,同时在1~5 m左右尺度的分布表现有聚集性分布的趋势(图5a,图6a)。2009年和2019年空间格局差异不明显。伴生种高山栲、滇青冈、滇石栋与地盘松种群空间关联分析表明,2009年地盘松与伴生种之间表现为互相独立(图5b),而2019年在0~22 m尺度上表现出较强的正关联(图6b)。
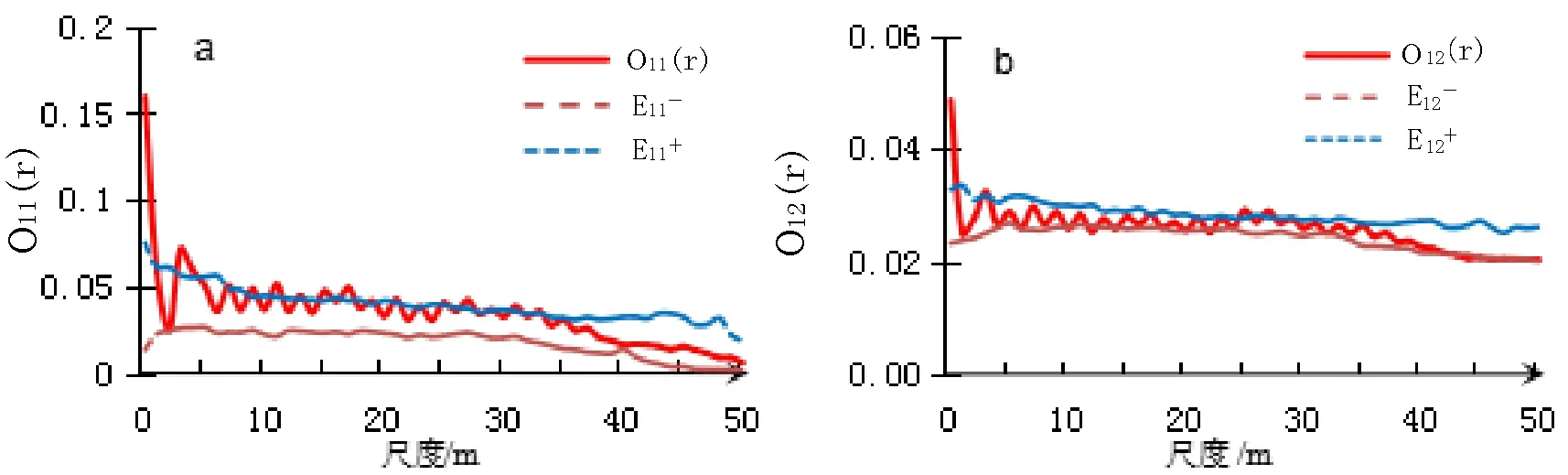
图5 2009年种群点格局和伴生种空间关联
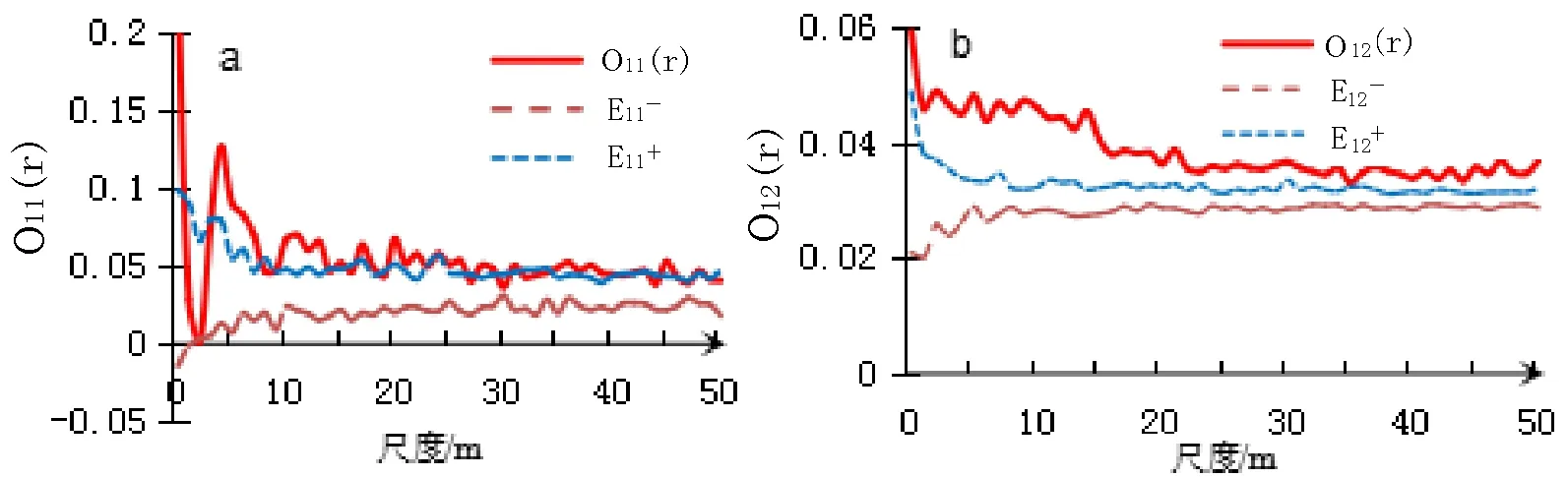
图6 2019年种群点格局和伴生种空间关联
3.3 形态参数与环境因子的相关性
选择复查样地的海拔、土壤水分指标(体积含水率)、土壤有机碳、种群密度、坡度等关键环境因子与形态特征参数HLR、CARCD、RNDLH、MLB,进行相关性分析(皮尔逊系数)。结果显示,各指标之间均未达显著水平(P>0.05),在环境因子和形态参数(表中黑体加粗部分)关系中,可清楚看出存在正相关和负相关的各有10对,多数为不相关或弱相关。其中,HLR与种群密度、土壤体积含水率、土壤有机碳含量都存在强负相关,CARCD与种群密度都存在强负相关,MLB与种群密度存在强正相关性,土壤有机碳含量与MLB、RNDLH存在较强相关性。
4 讨论与结论
4.1 讨论
比较地盘松的功能性状和格局演变的规律,来探索时序上的环境适应策略[30],从地盘松种群在时间跨度(10 a间)和空间尺度(0~50 m尺度序列)两个维度着手进行。重点关注地盘松种群这两个维度与资源获取互相妥协过程和环境适应策略的关系。
4.1.1 地盘松种群形态特征与环境适应的策略
本文比较了10 a间地盘松形态的变化,其中两个调查时期的树曲高比数(HLR)有明显的差异。但中林龄段的种群HLR没有明显分异(表2),而在幼林龄段的种群和成熟林龄段的种群的HLR有明显的分异,2019年的HLR明显居高(图1)。可以初步推断出经历较长时间跨度地盘松种群的盘曲主要发生在幼林龄段和成熟林龄段。此两阶段的形态调整来采取更加积极的环境适应策略,这与其他学者研究植物根部功能性状属性时表现出的不同的主动适应策略相似[30-32]。设置的HLR参数综合了地盘松树体的L和H表达的信息和两者的制约关系,剔出了因树体高生长掩盖的盘曲程度改变。HLR作为特殊树体的形态性状属性表达弯曲程度,相比工程学参数曲率半径更直接简易。HLR与土壤条件优劣成负相关也从侧面印证了其承载的环境适宜信息,是可供后来研究继续参考的指标之一。
地盘松种群的幅径截面比数(CARCD)在10 a前后有明显差异(表2),而且两期的CARCD在种群年龄结构上成明显持续降低的过程(图2)。根据CARCD的特征,地盘松随树龄增长相对生长势在衰弱。该指标充分考虑了冠幅作为与外界进行物质交换和碳水化合物制造机构的集合体,表征其生长势的重要性。同时因地盘松铺地程度不同造成冠幅水平投影的不同程度椭圆化(冠幅偏心率各异)。这种冠幅偏心率各异现象进而会引入冠幅被过高或过低估计的误差。因而CARCD指标中的冠幅通过椭圆计算冠幅截面积是比较可取的。CARCD是冠幅与地径截面积的比,可以表达地盘松经历逆境表现的相对生长势,数值越小表示生长越艰难,因生境或竞争而调整环境适应的策略,碳累积过程在减速。
地盘松种群的RNDLH在较长时间跨度上无明显差异(表2),种群年龄结构的序列上也无明显差异(图3),仅与土壤有机碳相关(表3)。这应该存在2种可能性:1)RNDLH本身设置不合理,L和H的差值没有明显生态学意义;2)地盘松种群以匍地盘曲获取资源和争取生态空间为主,种群竖向丰度本身生态信息过少,不是考察种群环境适应表现的良好指标。

表3 形态特征参数与环境因子的相关性
地盘松种群各林龄组、段的轮生枝间长均数(MLB)无明显差异(图4),但两期差异极显著。因MLB指标相对简单,仅表达轮生枝的连年生长量情况。经对比,2019年期的明显高,可知是近期新生长的较长,稍早则次之。此外,其还和种群密度密切正相关,可判断为环境适应策略性的徒长,以着生更加丰富的针叶进行碳累积。这种因环境适应发生的异速生长现象是否为松柏类植物所特有,目前尚无此类报道,仍需要进一步验证。
文中的4个形态参数与环境因子存在不同程度的相关性。从它们之间的相关特征看基本可印证指标所应承载的生态信息,同时也和指标的环境适宜特征与环境变量的动态关系大体一致。要详细研究几者间的对应关系,需要选择多样点采集数据(本文的固定样地研究并不能有效进行环境解释),再经连续的排序轴展示变量的关系和环境解释。
4.1.2 分布格局与环境适应的动态过程
种群的空间格局在不同发育阶段会表现不同的特征,这与森林群落的自然稀疏过程、干扰格局以及环境的变化有密切关系[31]。调查中发现,2009年和2019年两期的种群数量之差为57株,占种群数量的6.2%。主要表现为更新幼苗略有增加,而成熟林龄段的种群数量则有一定减少,这在格局研究的结果中得到了一定体现——在小尺度上,2019年种群较之2009年的有更大的分布强度。研究采用HP模型(考虑生境异质性),结果显示10 a之间格局无明显变化(图5、图6)。在1~50 m的尺度上主要表现为随机分布,分布强度变化趋势也一致。格局的变迁需要较长的时间跨度,经本研究验证至少应该大于10 a。调查地带凋落物回落腐化,在较长时间的跨度时,受到暴雨季节的地表径流、季节交替的阵风肆虐等因素作用后,土壤在有机质堆积和腐化程度的空间变迁上都发生较大改变[1,24]。这成为研究区微环境格局变化的主要驱动力[22],也是高山或山脊等恶劣立地下的普遍表现。在此微环境动态下,地盘松的种子库和更新幼苗的分布极大的影响了格局的动态[5]。但是在研究比较两期格局的变化后发现其在尺度上总体稳定,仅在1~5 m的小尺度存在一定差异。这和上述微环境的动态变化对更新幼苗、幼树影响有较大关联。在较大尺度上存在的中壮龄种群,则经历了群落的自然稀疏过程后,格局趋于稳定。在经历时间的变迁和风动力周期性的推动,分布格局与微环境格局之间互相妥协并互相牵制,妥协过程也是循环的主要驱动力之一。但这个循环在10 a的时间跨度上,是动态稳定的,如果出现动态变化应该把观测时间进一步拉大到10 a以上甚至更长。这也就反证了种群在短时间跨度内是不发生分布格局较大变化的环境适应表现,即其环境适应策略是不轻易启动分布格局调整的。研究的伴生种开始较大波动,如果推算下一个10 a,格局进一步发生扰动。时间跨度进一步拉大的话,建群种的格局不再稳定或将生存的领地上进行资源的重新配置。但伴生种的数量较少,样本量在对比上显得不足,表达更有价值的生态过程不唯一。下一步研究其更新特征时,可以更加明确其巩固生存的领地能力是否衰减,逆向演替是否会加速,以预测其将会由什么群落来替代。
4.2 结论
根据设置的形态性状参数的意义,经10 a的迁变,地盘松种群的茎干特征更加趋于趴地匍匐形态以占据更优越的生态空间,对领地中的其他物种形成明显的压制优势。特别是中幼林龄种群更趋于丰茂或稳定,树势更为强健。种群的自然树冠的竖向丰度在10 a间却无明显异变,表现稳定。这些确实表达了地盘松种群形态性状在较长时间跨度下形成较为多样化的环境适应表现,是非固化的适应策略应对。而在10 a的时间跨度下,种群格局表现相对稳定。地盘松种群在较长时间跨度下,并不轻易启用改变种群格局分布作为适应策略。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
今日农业(2022年15期)2022-09-20 06:54:16
丽水学院学报(2022年2期)2022-04-19 01:22:10
林业科技情报(2021年3期)2021-09-01 02:01:26
今日农业(2019年13期)2019-08-12 07:59:02
人民调解(2019年5期)2019-03-17 06:55:16
红土地(2018年7期)2018-09-26 03:07:38
中南林业调查规划(2017年1期)2017-12-19 00:58:07
小说月刊(2015年12期)2015-04-23 08:51:10
当代畜禽养殖业(2014年10期)2014-02-27 07:59:49
