
遮光对野鸦椿幼苗生长和光合生理特性的影响*
2021-04-22 13:42陈菊艳龙海燕邓伦秀
西部林业科学 2021年2期
陈菊艳,龙海燕,邓伦秀
(贵州省林业科学研究院,贵州 贵阳 550005)
光照是植物生长发育、形态建成的重要环境因子[1]。光照不足会降低植物的净光合速率,使其生长受阻[2],但是光强较多也并非对植物的生长有利,在强光下,植物光合机构难以全部利用所有的光能,过多的光照将导致光抑制或者光破坏,从而使光合速率降低[3]。在不同的光照条件下,植物大多可以通过形态和生理的可塑性来提高光能利用效率,以避免光抑制或光破坏[4]。通过人工设置不同遮光处理研究光照条件对植物生长发育的影响是常用的方法。因此,选择适宜的光照强度,将有助于珍贵树种的合理开发利用。
野鸦椿(Euscaphisjaponica)为省沽油科(Staphyleaceae)野鸦椿属(Euscaphis)[5-6]的落叶小乔木或灌木[7],其树形优美,春夏之交观其黄白色花朵,秋天观其红果[5,8],是一种集赏花、观果、观叶和观形为一体的优良园林绿化树种。同时,也是一种具有地域特色的多年生、低维护、低成本、3~4季均可观赏的珍贵树种[8],其主要分布于长江以南各省区,分布较为零星广泛[9]。目前,国内对野鸦椿的开发利用还处于起步阶段,尽管在福建、湖南、湖北等地有开发利用[10],但至今还未见其在园林中栽培应用。为充分开发野鸦椿的园林应用价值,本项目通过人工设置不同遮光条件对野鸦椿的生长发育及光合生理特性的研究,分析合理的光照栽培生态指标,寻求促进野鸦椿生长发育最适宜的光照环境,培育优质壮苗,进而为其制定保育策略、规模化引种栽培管理、园林配置应用提供理论依据。
1 材料与方法
1.1 试验地概况
试验地设在贵州省林业科学研究院树木园苗圃地(26°30′N,106°43′E,海拔1 100m)属于亚热带高原季风湿润气候,全年无霜期278 d,年平均降雨量1 198.9 mm,平均相对湿度77%,年平均气温15.2 ℃[11]。试验地气候温和,雨量充沛,较适宜野鸦椿的引种栽培。
1.2 试验材料和试验设计
试验选用的野鸦椿种子,于2017年10月采于贵州省毕节市纳雍县纳雍林场场部工区(26.67°N,105.26°E),采种后及时沙藏,2018年3月播种,发芽后于8月初选取生长健壮、长势一致的苗木移栽于塑料花盆中(上口径28 cm,下口径21 cm,高23 cm),栽培基质选用黄心土︰珍珠岩=4︰1(V︰V)。定植后移入遮光棚内,遮光棚长、宽、高分别为1.5 m×1.0 m×1.0 m,用黑色遮阳网模拟不同光照强度,试验共设置4个光照处理:自然光照(全光照CK);遮光率为光照强度的30%(A处理);遮光率为光照强度的60%(B处理);遮光率为光照强度的90%(C处理)。本试验采用单因素随机区组设计,每盆一株,每处理10盆,每处理3次重复。各处理浇水、施肥、除草管理都相同。
使用便携式光合测定仪(LI-COR)于遮光棚内随机选取5个点测定各处理的光合有效辐射,并计算各处理的遮光率[12]。
遮光率=(U0-U)/U0×100%
式中:U0表示全光照下的光合有效辐射,U表示各处理小区内的光合有效辐射。
1.3 测定指标与方法
1.3.1 生长特性
2019年10月开始各项观测、测定和分析。苗圃内2 a生实生苗,该苗木已按试验设计遮光处理1 a,各种生理、形态变化已稳定,测量其株高、地径、根幅及一级侧根数。
1.3.2 生物量测定
将采集的植株清洗干净,用吸水纸吸干多余水分,称量总鲜重,然后用剪刀将根、茎、叶分别剪下,用天平测量鲜重,然后分别将根、茎、叶装入牛皮纸袋放入烘箱,在105 ℃下杀青30 min,在70 ℃下烘干至恒量,称量干重。
1.3.3 生理指标
(1)光合色素用丙酮-乙醇混合液提取法[13-14];(2)可溶性蛋白含量用考马斯亮蓝法[12]测定;(3)可溶性糖含量用蒽酮比色法[14-16]测定;(4)丙二醛(MDA)含量用硫代巴比妥酸法[14]测定。以上各指标所需材料均在早上8:00对各处理随机采5枚相同部位的成熟叶片,去除中脉和叶柄,将所有叶片剪碎混合均匀测其相应指标,试验用722型分光光度计,每个处理各指标均重复3次,取平均值。
1.3.4 光合日变化
用LI-6400便携式光合作用测定仪(LI-COR Inc.USA),使用开放气路,空气流速为0.5 L/min。7—8月选择晴天少云的天气,从9:00-19:00,每2 h测一个轮回,选择生长旺盛并具有代表性的植株,取其生长良好的成熟叶片测定光合指标,测定指标包括单叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度Gs)、胞间CO2浓度(Ci)等,同时记录叶室内光合有效辐射(PAR)、空气CO2浓度(Ca)、气温(Ta)、叶温(Ti)、大气相对湿度(RH)等环境因子数据。光合作用的气孔限制值(Ls)按Berry和Downton[17]的公式计算。每处理重复3株,每株测定3片叶子。
1.4 数据处理
用Excel 2008软件进行数据的录入和处理,并绘制图、表;用SPSS 18.0软件进行方差分析、相关性分析以及其它统计分析。
2 结果与分析
2.1 遮光对野鸦椿生长特性的影响
遮光对野鸦椿幼苗生长特性的影响见表1。植株的苗高生长量随遮光程度的增强而先升高后降低,其中B处理苗高生长量最大,较CK处理高47.70%(P>0.05),CK处理苗高生长量最低;地径生长量随遮光程度的增强而呈先增加后减小的趋势,其中地径最大的是B处理,较CK增加6.03%,最小的是C处理,较CK减少7.51%,各处理间均无显著差异;叶片数随着遮光程度的增加呈现出先增加后减小的趋势,叶片数较多的是A处理,较少的是CK处理;根长呈现出随遮光程度的增加先增加后减小的趋势,根长最长的是B处理,比CK高16.34%,最短的是C处理,比CK短1.00%,各处理间均无显著差异;根幅也呈现出随遮光程度的增加先增加后减小的趋势,最大的是B处理,最小的是C处理;侧根数呈现出随遮光程度增加不断减小的趋势,各处理间均无显著差异。遮光30%、60%处理下,野鸦椿植株的生长量均高于对照,以B处理下苗高、地径、根长、根幅的增幅最大,遮光90%处理下,地径、根长、根幅和侧根数均小于对照。说明适度遮光有利于野鸦椿幼苗的生长,但是不同遮光处理间各指标差异不显著(P>0.05)。
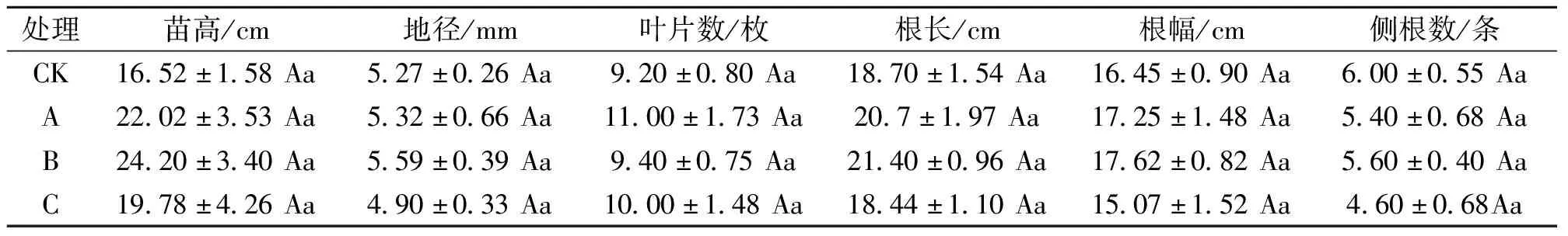
表1 遮光对野鸦椿的生长状况的影响
2.2 遮光对野鸦椿生物量的影响
生物量可以直接表示植物的物质产量[18]。从表2可知,随着遮光程度的不断增加其根鲜重、根干重表现出先增后减的趋势。其中根鲜重最大的是A处理,较CK处理高9.53%(P>0.05),最小的是C处理,较CK处理低267.88%(P<0.01);根干重最大的是B处理,较CK处理高28.48%,最小的是C处理,较CK处理低238.90%,A处理和C处理间有显著差异(P<0.05)、B处理和C处理也有显著差异(P<0.05)。随着遮光程度的不断增加茎鲜重、茎干重也表现出先增后减的趋势,其中茎鲜重最大的是B处理,最小的是C和CK处理,各处理间均无显著差异;茎干重最大的也是B处理,最小的是C处理,各处理间均无显著差异。随着遮光程度的不断增加其叶鲜重、叶干重也表现出先增后减的趋势,其中叶鲜重最大的是B处理,较CK处理高91.89%(P<0.01),最小的是CK处理;叶干重最大的是A处理,较CK处理高58.00%,最小的是CK处理,各处理间均无显著差异(P>0.05)。随着遮光程度的不断增加其总鲜重、总干重也表现出先增后减的趋势,总鲜重、总干重最大的均是B处理,最小的均是C处理;总鲜重最大的比CK高22.62%,最小的比CK低90.33%;总干重最大的比CK高20.69%,最小的比CK低86.86%。不同遮光处理对野鸦椿根、茎、叶和总生物量的影响程度均不同,B处理下茎、叶、总生物量最高,其次是A处理,C处理最低。试验发现A、B处理的植株长势良好,叶色较全光照下深;C处理下叶色较深,但是根、茎积累的生物量却相对较少;CK处理下叶片小,叶色较浅,有的叶片出现卷曲,长势不良。可见适度的遮光有利于野鸦椿幼苗生物量的积累,光照过强或过弱都会导致野鸦椿幼苗生物量的下降。

表2 遮光对野鸦椿单株生物量的影响
2.3 遮光对野鸦椿幼苗生理指标的影响
2.3.1 遮光对野鸦椿光合色素含量的影响
从表3可以看出,遮光处理后,野鸦椿幼苗的叶绿素a(Chla)、叶绿素b(Chlb)及叶绿素总量(Chla+Chlb)的含量在C处理下均达到最大,分别比CK增加63.08%、129.11%、77.78%(P<0.01),表明弱光下野鸦椿幼苗能够通过增加叶绿素含量来提高光捕获能力,从而提高光能利用效率。叶绿素a/b(Chla/Chlb)最大值是CK处理,CK和C处理之间有极显著差异(P<0.01),CK和A、B处理之间有显著差异(P<0.05),总体表现出随遮光的增加而不断减小的趋势,表明叶绿素b含量的增幅大于叶绿素a含量。类胡萝卜素(Caro)的作用主要是防止光照伤害叶绿素,野鸦椿幼苗叶片中类胡萝卜素含量随着遮光程度的不断增加呈先增加后减小的趋势,类胡萝卜素最大的是B处理,较CK处理高72.96%(P<0.01),最小的是CK处理,CK和A处理之间无显著差异,其他各处理间有极显著差异(P<0.01)。

表3 不同光照条件下野鸦椿叶片的光合色素含量
2.3.2 遮光对野鸦椿叶片可溶性蛋白和可溶性糖含量的影响
野鸦椿叶片的可溶性蛋白含量在遮光条件下显著低于全光照生长条件下,与全光照相比降幅分别为42.20%、43.50%、68.28%,A和B处理之间无显著差异(P>0.05),其他各处理均有极显著差异(P<0.01)。总的来说在全光照条件下蛋白质含量相对较高,遮光处理下含量较低,表明光照强度的减弱,影响叶片内蛋白质的积累,从而导致野鸦椿叶片蛋白质含量降低。
不同遮光处理下,野鸦椿幼苗叶片可溶性糖变化见图2。随着遮光程度的增加,野鸦椿叶片可溶性糖含量呈现先升后降低的趋势。其中可溶性糖含量最大的是A处理,较CK处理高57.90%(P<0.01);最小的是B处理,较CK处理低11.62%(P<0.01),各光强下均有显著性差异(P<0.01)。表明适度的遮光有利于叶片中可溶性糖含量的积累,遮光强度较低时不利于可溶性糖含量的产生与积累。可能是因为适度的遮光处理后光合速率增强,促进糖的合成积累,而随着遮光程度的加强,其光合速率降低,抑制了糖的合成与积累。
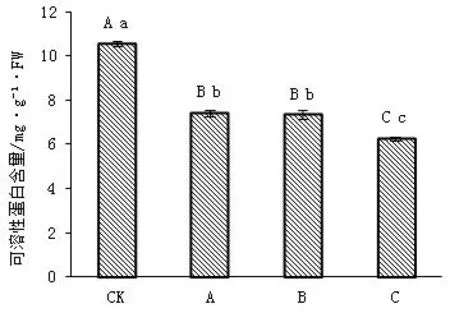
图1 遮光对野鸦椿幼苗叶片可溶性蛋白含量的影响
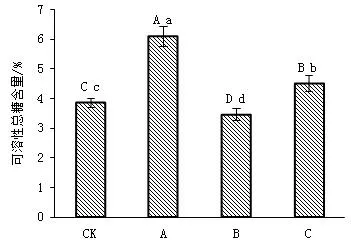
图2 遮光对野鸦椿幼苗叶片可溶性糖含量的影响
2.3.3 遮光对野鸦椿MDA含量的影响
图3表明,在A、B处理下,叶片中丙二醛(MDA)的含量显著低于CK处理,分别比CK处理下降了21.94%、4.73%,CK和A处理有极显著差异(P<0.01),CK和B处理有显著差异(P<0.05),C处理比CK处理下高111.02%。MDA是膜脂过氧化物,表示膜脂过氧化强度和膜系统受伤害程度[19]。结果表明,遮光30%、60%条件下减少了野鸦椿生物膜的伤害,对野鸦椿幼苗生长有利,而遮光90%环境下对野鸦椿产生了不同程度的伤害,对野鸦椿的生长不利。

图3 遮光对野鸦椿幼苗叶片MDA含量的影响
2.4 遮光对野鸦椿光合作用日变化的影响
2.4.1 遮光条件下环境因子日变化
从光合有效辐射(PAR)的变化来看(图4),全光照条件下15:00时PAR出现峰值为1 202.93 μmol/(m2·s),遮光条件下PAR显著下降,均在13:00时达最高值,分别为A处理495.60 μmol/(m2·s),B处理194.18 μmol/(m2·s),C处理50.69 μmol/(m2·s)。遮光对大气CO2浓度变化影响不大,各处理下Ta和TL呈现出相似的变化趋势,在13:00时遮光处理下Ta和TL显著低于对照(图5)。遮光影响了苗木生长的小环境,有利于维持植物的水分含量和环境中的相对湿度,进而影响苗木的生长。

图4 不同光照条件下野鸦椿光合有效辐射和大气CO2浓度日变化
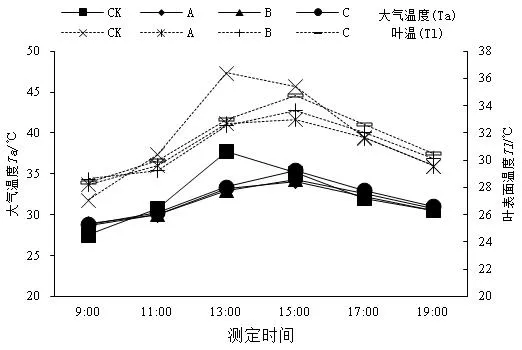
图5 不同光照条件下野鸦椿大气温度和叶表面温度日变化
2.4.2 遮光对野鸦椿光合作用参数的影响
(1)遮光对净光合速率(Pn)的影响 不同光照环境下野鸦椿叶片Pn的日变化趋势显著不同(图6)。在全光照(CK)处理下Pn呈双峰曲线,高峰分别在11:00时[峰值为12.43 μmol/(m2·s)]和15:00时[峰值为4.28 μmol/(m2·s)],第二个峰值约为第一个峰值的65.57%;在13:00时有明显的“午休”现象。各个遮光处理下叶片Pn均为单峰曲线,高峰均出现在13:00时,各处理的峰值分别为A处理16.14 μmol/(m2·s)、B处理13.60 μmol/(m2·s)、C处理11.10 μmol/(m2·s),均没有出现“午休”现象,以遮光30%处理的Pn较高,表明野鸦椿适度遮光后有利于光合作用的增强。

图6 不同光照条件下野鸦椿净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、气孔限制值(Ls)的日变化
(2)遮光对气孔导度(Gs)的影响 气孔导度(Gs)反映了大气 CO2和水汽与植物叶片传导、交换能力的高低[20]。由图6可知,13:00时之前各处理叶片Gs较高,后期逐渐下降,整个Gs日变化中,除了CK处理9:00时较高之外,其他时间段A处理的Gs一直高于其他各处理,说明A处理的光强有利于野鸦椿叶片气孔的调节,也有利于光合作用效率的提高。各处理Gs日变化均表现为上午的Gs高于下午,下午由于高光、高温和低湿的作用,促使野鸦椿叶片部分气孔关闭,气孔导度不断的降低,致使净光合速率也不断下降。
(3)遮光对胞间CO2浓度(Ci)的影响 遮光处理下胞间CO2浓度的日变化曲线可以看出(图6),遮光处理下野鸦椿叶片从9:00开始其Ci便一直下降,到13:00时达到日渐最低值,然后又迅速升高,呈“V”型,而对照CK处理下从9:00时开始下降,11:00达到第一个最低值,然后又升高,接着又降低,到15:00达到第二个低值,呈“W”型,基本上与净光合速率变化相反。
(4)遮光对蒸腾速率(Tr)的影响 不同光环境下野鸦椿幼苗的Tr的日变化见图6。A处理蒸腾速率日变化呈“单峰”变化曲线,最高峰出现在13:00,其他各处理呈“双峰”变化曲线,最高峰出现在9:00和13:00,但是总体表现是上午的蒸腾速率高于下午的蒸腾速率,上午随着光合有效辐射强度的不断增强,温度也不断升高,蒸腾速率也不断地增大。13:00以后,蒸腾速率与净光合速率都出现下降。
(5)遮光对气孔限制值(Ls)的影响Ls反映了植物叶片对大气CO2相对利用效率的大小[21]。不同处理下野鸦椿幼苗叶片的气孔限制值(Ls)的日变化见图6。各遮光处理下Ls呈“单峰”曲线,Ls的日变化与Ci的日变化相反,当Ci达到低谷时,Ls达到最高值,全光照下呈“双峰”曲线,峰值出现在11:00和15:00,第一个峰值明显高于第二个峰值,从各个时间段比较来看,A处理下Ls测定值要大于其他处理。15:00以后野鸦椿的光能利用效率都很低。
3 讨论与结论
植物的生长发育与其所处的光照条件密切相关,不同的光照条件可以引起植物的可塑性反应。一般地,在一定的遮光条件下阴性植物往往会达到最佳生长状态,即正常高度,不徒长等[22]。在轻度遮光时,植物可以通过生长发育和形态特征调节以主动适应环境,如加快株高生长、扩大叶面积等;在重度遮光时,生长发育和形态特征会受到严重阻碍,表现出植株瘦弱、茎干藤蔓化、叶色发黄及死亡等现象[22]。本研究表明,苗高、地径、根长、根幅均在遮光60%条件下较大,叶片数在遮光30%处理下最多,这表明遮光30%、60%条件对野鸦椿幼苗的生长有更大的促进作用。过度的遮光使野鸦椿苗木出现一定的徒长,茎干细弱,叶片变薄,有“倒伏”现象出现,且分生芽数没有遮光30%、60%条件下多,但是在3种遮光条件下均无死苗,未出现黄化和瘦弱等不适应现象,说明野鸦椿能在适度的遮光环境下生长。
光合作用是植物积累有机物和增加自身生物量的最根本途径[23],植物对光的喜好程度不同,不同的光照环境对植物生长有很大的差异,适度地遮光处理对有些植物的生长也是非常有利的,并且能够增加生物量,提高品质[24]。从干物质含量来看,遮光30%、60%环境下野鸦椿积累较多生物量,可见适度的遮光有利于野鸦椿生物量的积累。
本研究表明,随着遮光程度的不断增加,叶绿素a、叶绿素b含量和总叶绿素含量不断增加,说明在遮光环境下野鸦椿幼苗叶片更绿。光合色素含量的增加可以显著增强光系统吸收、传递和转化低密度光量子的能力[12],这表明遮光环境下野鸦椿幼苗能通过增加叶绿素a、叶绿素b和总含量来提高光捕获能力,进而提高对光能的利用效率。遮光后,野鸦椿叶片的叶绿素a/b低于全光照,大大增强了利用蓝紫光的能力,遮光通常会降低红光吸收比例,增高蓝光的吸收比例[25],所以植物能通过降低叶绿素a/b来适应荫蔽的环境[26]。类胡萝卜素(Caro)的作用是防止叶绿素光损伤,野鸦椿幼苗叶片中Caro含量随着遮光程度的不断增加呈先增加后减小的趋势,在遮光60%处理下含量最高,在全光照条件下最低,说明在遮光60%的环境下有利于提高野鸦椿叶片吸收和传递光能的能力,充分利用弱光条件下的光能。
可溶性蛋白质含量的高低与植物的衰老密切相关,含量越高对植物生长越有利,含量降低是对植物逆境伤害的表现[27]。较高光强有利于野鸦椿叶片可溶性蛋白的合成和积累,较低的光强不利于野鸦椿叶片可溶性蛋白含量增加,其中以遮光90%环境下受影响最大。可溶性糖是植物生长中许多生理生化反应和重要能量来源的基础[28]。试验表明,全光照条件下野鸦椿叶片可溶性蛋白含量较大,遮光30%条件下可溶性糖含量积累较多,说明适度遮光对调节植物细胞组织渗透势、保持膜的完整性、增强植物抗性起重要作用[29]。
丙二醛是膜脂过氧化物,具有细胞毒性[30]。MDA较多的积累,这表明植物组织的保护能力较弱[31],植物的抗逆性和抗衰老能力也较弱[32]。本试验中,遮光90%环境条件下植物叶片的丙二醛含量明显高于其他各处理条件,表明重度遮光环境下发生了较严重的膜脂过氧化,植物遭到了逆境胁迫。全光照条件下,野鸦椿叶片丙二醛含量高于遮光30%、60%处理,说明在全光照条件下野鸦椿生长也受到了一定程度的影响,遮光30%和遮光60%处理下丙二醛含量较低,膜脂过氧化程度最轻。
本研究表明,野鸦椿在全光照环境下净光合速率日变化呈双峰曲线,峰值分别出现在11:00和15:00,11:00的净光合速率[峰值为12.43 μmol/(m2·s)]高于15:00时的净光合速率[峰值为4.28 μmol/(m2·s)],净光合速率在13:00左右出现低谷,有光合“午休”现象,可能是因为增加光照强度,植物大量利用空气中的CO2使其浓度下降,导致部分气孔关闭,使其净光合速率降低,也可能是出现了光抑制以及光呼吸的加强导致净光合速率降低[33]。胞间CO2浓度(Ci)降低和气孔限制值(Ls)增大,为气孔限制,Ci增大和Ls减小,则为非气孔限制[34]。本试验中,不同光照条件下野鸦椿叶片的Ci和Ls日变化有显著不同。全光照条件下在13:00明显下降,Ci持续增加和Ls明显下降。所以,此时“光合午休”现象的出现主要原因是非气孔限制造成的,即午间光照强度过高导致大气相对湿度较低,大气温度和叶片表面温度的升高,造成植物暗呼吸和光呼吸的上升,进而抑制了全光照环境下野鸦椿净光合速率的升高,导致 “午休”现象的产生。各遮光处理下叶片净光合速率均为单峰曲线,高峰均出现在13:00,各处理的峰值分别为A处理16.14 μmol/(m2·s)、B处理13.60 μmol/(m2·s)、C处理11.10 μmol/(m2·s),均没有出现“午休”现象,即使中午前后Ci有所降低,野鸦椿Pn保持在相对较高的水平,有利于野鸦椿积累更多的光合产物,说明遮光可避免野鸦椿的“光合午休”现象,但只有适度遮光才能提高其净光合生产力。以遮光30%环境下有较高的光合速率,但随着光照强度的的进一步减弱,光合速率反而降低,说明遮光程度太重(即遮光90%),使光照强度明显不足,即使在这种环境下野鸦椿叶片有较高的叶绿素含量,仍然不足以使光合速率提高,光照强度成了限制野鸦椿光合速率的重要因子。
综上所述,野鸦椿具有较好的生理调节机制来适应不同的光环境,在贵州地区的自然光强的中午对野鸦椿生长有一定的负效应,但是其对适度遮光又有较好的适应性,可避免光合“午休”的出现,重度遮光会制约野鸦椿幼苗的健康生长,因此在育苗生产中,建议有效辐射强度保持在遮光率为30%~60%的范围,可培育出优良健壮的苗木。野鸦椿在园林中进行配置应用时,宜采用与其他乔、灌、草结合形成人工植物群落,以使植株间互相遮挡阳光,减轻“光合午休”,降低强光对野鸦椿的光伤害,提高树丛内或群落内的湿润度,降低呼吸蒸腾量,为野鸦椿的正常生长创造必备的生态环境条件。
猜你喜欢
中国机械工程(2022年8期)2022-05-09
儿童时代·幸福宝宝(2021年1期)2021-03-29
小资CHIC!ELEGANCE(2019年40期)2019-12-10
音乐教育与创作(2019年8期)2019-05-16
中成药(2018年8期)2018-08-29
新课程·下旬(2017年7期)2017-08-14
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年6期)2015-02-06
西北林学院学报(2015年5期)2015-01-03
