
不同分解等级的兴安落叶松倒木细菌多样性分析
2021-04-22 13:20杨立宾崔福星黄庆阳朱道光
中南林业科技大学学报 2021年4期
杨立宾,崔福星,黄庆阳,朱道光,徐 飞
(1.西华师范大学,四川 南充 637002;2.黑龙江省科学院 自然与生态研究所,黑龙江 哈尔滨 150040)
倒木(Fallen log)是粗木质残体(Coarse woody debris,CWD)的重要组成部分,是生态系统中的一个生态单位[1],是森林生态系统中结构和功能的重要组成元素[2];其在森林生态系统的更新与演替,生物多样性的维持与保育,物质的循环以及能量的传递等方面起着非常重要的作用[3]。倒木的物种不同、径级不同、生态系统中立地条件以及分解者的差异等都能够影响倒木的腐烂程度和分解过程。微生物不仅仅是倒木的主要分解者,同时也是消费者,能够敏感、直观的反映影响倒木分解速率的各种环境因素。有研究指出,不同的分解时间(分解阶段)和不同部位(树皮、边材、心材)的倒木分解具有不同的微生物组成和微生物量[4-6]。现阶段更多的研究集中在真菌类群[7-8],但真菌并不是参与倒木分解的唯一微生物类群[9]。细菌普遍存在于倒木上[10],并与该环境下的真菌群落相互作用共同参与木材的分解[11]。有研究表明,倒木的分解改变了细菌的丰富度,倒木的pH 值、碳氮比、含水量、养分等对细菌的群落有影响[12],而细菌在倒木分解的过程中能够改变倒木的结构、破坏完整性使其更加透水等[13]。过去由于研究手段的局限导致对微生物认识的不完整,目前关于参与倒木分解的细菌群落的生态习性、多样性组成以及与真菌之间的相互作用等许多问题并不是十分明确,还需进一步研究与探讨。
兴安落叶松是我国寒温带森林生态系统的主要建群种,是大兴安岭林区重要的森林资源树种。目前有关兴安落叶松林倒木研究的报道主要集中在倒木储量、养分含量、分解速率及温室气体排放等方面的相关研究,而对兴安落叶松倒木分解过程微生物的分解者,特别是针对细菌的相关研究未见报道。因此,为明确细菌在倒木分解不同阶段的多样性和组成,为评价细菌在倒木分解过程中的作用和贡献,本研究以大兴安岭兴安落叶松林倒木为研究对象,采用高通量测序的方法研究不同分解阶段倒木上的细菌群落的结构特征、组成、多样性与倒木理化性质特征之间的关系,以期为深入理解我国大兴安岭地区寒温带兴安落叶松林倒木分解过程及影响因素提供科学依据。
1 材料与方法
1.1 研究区域概况
本实验的研究区域位于我国寒温带大兴安岭地区的黑龙江省呼中国家自然保护区内,实验样 地(51°49′01″~51°49′19″N,122°59′33″~123°00′03″E)位于2014年建成的大兴安岭兴安落叶松林25 hm2国际性监测样地内。监测样地的地形平缓,海拔为847~974 m,年平均气温为-4℃,年平均降水量为458.3 mm,年平均相对湿度为71%,年平均蒸发量为911 mm。监测样地的土壤为棕色针叶林土,土壤的厚度为5~25 cm。监测样地的植被群落结构比较单一,是以兴安落叶松为主要优势种的北方明亮针叶林;植被中包括4 种乔木,37 种灌木,127 种草本,分别隶属于21 科39 属。实验样地中的乔木为兴安落叶松Larix gmelinii和白桦Betula platyphylla,主要灌木包括杜香Ledum palustre、笃斯越桔Vaccinium uliginosum等,主要草本包括舞鹤草Maianthemum bifolium、地榆Sanguisorba officinalis等[14]。
1.2 研究方法
根据闫恩荣等[15]的倒木分解等级标准将其划分为5 个腐烂等级,分别用Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ级表示。2019年6月11日在样地中选择杜香-兴安落叶松林阳坡地势平坦的样方,调查选取立地条件相似且储量最大的倒木(径级为20~30 cm)[16],分别取5 个分解等级各3 株倒木混合样品(因兴安落叶松Ⅳ级、Ⅴ级分解等级的倒木无法准确确定树皮、边材和心材,所以从Ⅰ~Ⅴ级倒木均采用树皮、边材和心材混合取样;对于Ⅰ、Ⅱ、Ⅲ级分解等级的倒木,径向截取2 cm 的木段,Ⅳ、Ⅴ级分解等级的倒木取大于2 cm 长度且等体积的样品装入取样袋),去除表面的泥土、根系和苔藓等附着的物质,做好标记后装于无菌样品袋内低温保存,尽快带回实验室进行分析。将每个样品分成两部分,一部分低温研磨、混匀后于-80℃中储存,用于细菌多样性测定;另一部分用于基质物质[18]和理化性质测定。
16S rDNA 基因组提取采用试剂盒(Carlsbad,CA,USA)法提取。以50 ng DNA 为模版,采用(338F/806R)通用引物进行扩增。PCR 扩增体系为:5×FastPfu 缓冲液4 μL,2.5 mmol·L-1的dNTP 2 μL,5 μmol·L-1的Primer F 0.8 μL,5 μmol·L-1的Primer R 0.8 μL,5 U·μL-1的Polymerase 0.4 μL,BSA 0.2μL,DNA 模板10 ng,补超纯水(dd H2O)至20 μL。扩增条件为:95℃预变性3 min,95℃变性30 s,45℃退火30 s,72℃延伸30 s,5 个循环,二次扩增95℃ 20 s,55℃退火20 s,72℃延伸30 s,20 个循环,最后72℃延伸5 min。用2%的琼脂糖电泳检测相同样本混合后的PCR 产物,使用AxyPrep DNA Gel Extraction Kit (Axygen Biosciences,Union City,CA,USA) 凝胶回收试剂盒纯化PCR 产物[14]。将纯化扩增产物等摩尔混合,构建Miseq 文库,使用Illumina MiSeq 平台测序[17]。物种分类采用silva132/16s_bacteria 的细菌数据库进行比对和鉴定,生物信息学分析采用上海美吉生物医药科技有限公司的微生物多样性云分析V4.0 平台。
1.3 统计分析
生物信息学分析由上海美吉生物医药科技有限公司的微生物多样性云分析V4.0 平台协助完成。使用Qiime 软件进行数据去杂。以97%相似性为标准划分操作分类单元(Operational taxonomic units,OTU),使用Usearch 去除嵌合体。采用RDP classifier 贝叶斯算法对OTU 代表序列进行分类学分析,置信度阈值为0.7。采用mothur 软件进行α 多样性指数分析和rarefaction 分析[14],分别用Coverage 指数、Ace 指数、Shannon 指数和Simpson 指数表示测序深度、物种的多度、丰富度和均匀度。β 多样性基于Bray-Curtis 距离计算并进行PCoA 分析,用FastUniFrac 分析得到样本间距离矩阵。环境因子关联分析采用db-RDA 分析和相关性 Heatmap 分析(采用Pearson 法)。使用Excel 2007 和 SPSS 19.0 软件对实验数据进行统计分析,采用单因素方差分析(ANOVA)和多重比较法(LSD)对倒木的总碳、总氮、含水率、纤维素、半纤维素、木质素含量的差异和显著性进行分析。
2 结果与分析
2.1 倒木养分含量分析
从表1可以看出,随着腐烂程度的增强,倒木的含水率从Ⅰ到Ⅴ级分解等级显著升高。倒木总氮含量从Ⅰ级到Ⅴ级分解等级逐渐升高,Ⅰ级至Ⅳ级分别与Ⅴ级存在显著性差异。总碳含量从Ⅰ级到Ⅴ级逐渐降低,Ⅱ、Ⅲ、Ⅳ级之间无显著性差异,但与Ⅰ级和Ⅴ级存在显著性差异。碳氮比从Ⅰ级到Ⅴ级逐渐降低,其中Ⅱ、Ⅲ、Ⅳ级之间无显著性差异,但与Ⅰ级和Ⅴ级的存在显著性差异。

表1 不同分解等级的倒木理化性质比较†Table 1 Comparison of physical and chemical properties of fallen wood with different decomposition levels
倒木纤维素的质量分数从Ⅰ到Ⅴ级逐渐降低,Ⅰ级和Ⅱ级之间无显著性差异,Ⅲ、Ⅳ级和Ⅴ级之间存在显著性差异。倒木半纤维素的质量分数从Ⅰ到Ⅴ级逐渐降低,变化趋势与纤维素相同,Ⅰ到Ⅴ级之间呈显著性差异。倒木木质素的质量分数从Ⅰ到Ⅴ级逐渐升高,Ⅰ级和Ⅱ级之间无显著性差异,Ⅲ级和Ⅳ级之间无显著性差异,Ⅴ级倒木的木质素质量分数最高,并与各组之间存在显著性差异。
2.2 测序结果及取样深度验证
倒木样品的细菌测序通过优化过滤后得到的有效序列总数为864 035 个,有效的碱基数为376 325 347 个,有效序列平均长度为435.54 bp。倒木样品的细菌测序序列经拆分、去冗余后在97%相似度下进行OTU 聚类,共得到1 360 种OTUs。
用Sobs 多样性指数构建稀释曲线,能够表征实际观测到的物种数目,当曲线趋于平坦时,表示测序的数据量合理。如图1所示,不同分解等级倒木样品细菌的稀释性曲线均趋于平缓和饱和,说明供试样品的取样合理,测试数据量合理,置信度较高,能够真实的反映不同分解等级倒木样本的细菌群落结构特征。

图1 相似度为0.97 条件下样品的稀释性曲线Fig.1 Dilution curve of samples with similarity of 0.97
如图2所示,不同分解等级倒木细菌OTUs 总数Ⅴ级>Ⅳ级>Ⅰ级>Ⅱ级>Ⅲ级,分别为870、721、651、581、530 种。5 个分解等级倒木细菌共同所有的OTUs 数目为204 种,独有的OTUs Ⅰ级为52 种,Ⅱ级为19 种,Ⅲ级为27 种,Ⅳ级为96 种,Ⅴ级为338 种。从各级分解等级倒木细菌的OTUs 总数和独有数上来看,各组之间存在较大不同。

图2 各分解等级倒木上细菌的OTUs 比较Fig.2 OTUs comparison of bacteria on fallen wood with different decomposition levels
2.3 倒木上的细菌群落分布特征分析
在相似水平为97%的条件下,对OTUs 的代表序列做分类学分析,共鉴定20 门47 纲126 目222 科377 属637 种。在门分类水平上分属于酸酐菌门Acidobacteria、放线菌门Actinobacteria、拟杆菌门Bacteroidete、绿弯菌门Chloroflexi、蓝细菌门Cyanobacteria、厚壁菌门Firmicutes、浮霉菌门Planctomycetes、变形菌门Proteobacteria、疣微菌门Verrucomicrobia、Patescibacteria和WPS-2等菌门。从图3可以看出,变形菌门和放线菌门为各分解等级倒木细菌的主要共有优势菌。此外,Ⅰ级分解等级倒木上的细菌主要优势种还包括拟杆菌门和疣微菌门;Ⅲ级还包括酸酐菌门;Ⅴ级还包括厚壁菌门。各分解等级倒木细菌在门分类水平上除变形菌门和浮霉菌门无差异外,其余各门存在显著性差异,其中WPS-2 各级为极显著差异。

图3 门水平上细菌的组成Fig.3 Bacteria composition of fallen wood with five decomposition grades at the phylum taxonomic level
2.4 Alpha 多样性指数分析
对不同分解等级的倒木细菌16S rDNA 的Alpha 多样性指数进行ANOVA 分析,从表3可以看出各分解等级的Coverage 指数Ⅰ级至Ⅳ级均与Ⅴ级存在显著性差异,但数值都趋近于1,说明样本中序列被测出的概率高,反映本次测序结果能够代表样本中微生物的真实情况。细菌Ace 指数Ⅴ级指数最高,Ⅲ级指数最低,各级之间存在显著性差异,说明各级分解等级倒木样品中的细菌物种组成总数存在差异。Shannon 指数Ⅱ级与其它各级之间分别存在显著性差异,说明其与各组之间的丰富度不同。Simpson 指数Ⅱ级与Ⅰ、Ⅲ、Ⅳ级存在显著性差异,但与Ⅴ级无差异,说明其优势种群的生态优势度较低,也说明其均匀度最高。

表2 相似度为0.97 条件下各倒木样品多样性指数分析Table 2 Diversity index analysis of fallen wood samples of different decomposition grades with similarity of 0.97
2.5 Beta 多样性分析
基于Bray-Curtis 距离对不同分解等级的倒木上细菌群落组成进行主坐标PCoA(Principal coordinates analysis)分析,从图5可以看出,各组相同标识的样品之间相对距离比较近,说明各组内的样品相似性较高。在X 轴上各组样品分为两大组,第一组在X 轴的负方向上(Ⅰ级和Ⅱ级,Ⅲ级和Ⅳ级),第二组在X 轴的正方向上(Ⅴ级);此外第一组的Ⅰ级和Ⅱ级共同在第三象限且被置信椭圆囊括,第二组中的Ⅲ、Ⅳ级共同在第二象限且Ⅲ级和Ⅳ级之间距离较小,说明Ⅰ级和Ⅱ级的细菌组成更加相似,Ⅲ级和Ⅳ级的细菌组成相似。第一轴的解释率为33.04%,第二轴的解释率为28.89%,累计解释率为61.93%。
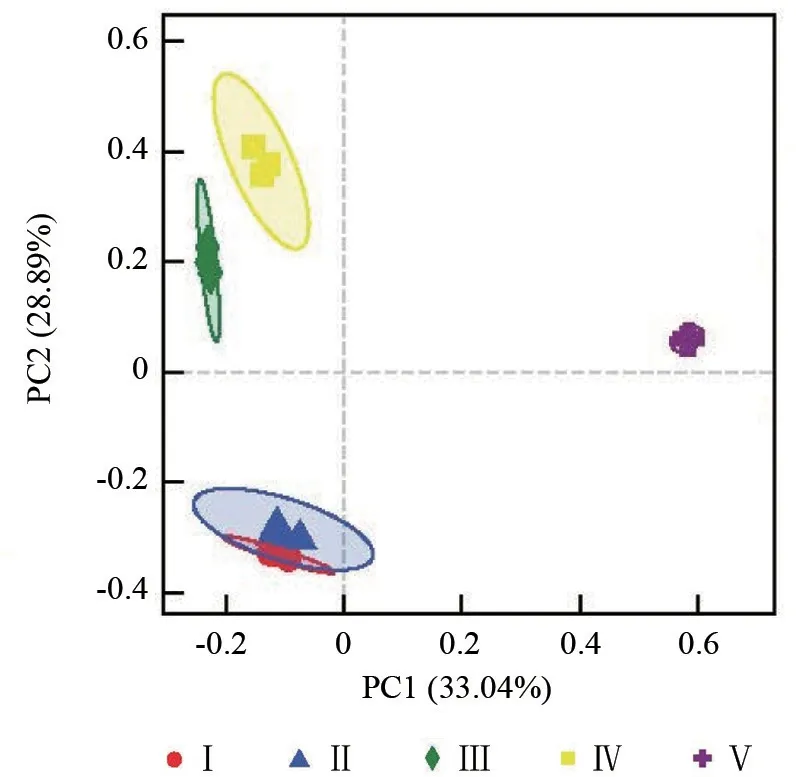
图4 各分解等级倒木的细菌Beta 多样性分析Fig.4 Beta diversity analysis of bacteria on fallen wood at different decomposition levels
2.6 冗余分析
将97% 相似性的样本OTU 表做DCA 判别,分析结果中Lengths of gradient 第一轴的数值≥3.5,因此选择CCA 进行分析和讨论。CCA排序结果从图5可以看出,两轴的累计贡献率为48.11%,第一轴解释细菌群落变化的27.42%,第二轴解释了20.69%。全部样品为3 组,Ⅰ级和Ⅱ级为一组处于第二象限,Ⅲ级和Ⅳ级为一组处于第三象限,Ⅴ级处于第一象限。

图5 基于倒木细菌OTU 水平和养分因子的冗余分析Fig.5 CCA based on the bacteria OUT level and nutrient factors of fallen wood
相关性的Heatmap 分析是通过计算环境因子与物种间的相关性系数(Pearson 相关系数、Spearman 等级相关系数等),并通过Heatmap 图直观展示数值。不同的颜色变化反映二维矩阵或表格中的数据信息,颜色深浅表示数据值的大小。本研究选取各分解等级倒木的理化性质与细菌在门水平上的Pearson 相关系数进行相关性 Heatmap分析,从图6可以看出,半纤维素含量与放线菌门极显著正相关(R=0.727 6),与酸杆菌门(R=0.591 4)和WPS-2(R=0.559 1)显著正相关,与绿弯菌门极显著负相关(R=-0.709 9);木质素含量与厚壁菌门极显著正相关(R=0.883 3),与拟杆菌门显著负相关(R=0.551 0)。优势菌门中,变形菌门与总氮含量极显著负相关(R=0.811 4),与碳氮比极显著正相关(R=0.792 9);放线菌门与半纤维素含量极显著正相关(R=0.727 6),与碳氮比显著正相关(R=0.532 1)。

图6 基于倒木细菌门水平和养分因子的相关性Heatmap 图Fig.6 Heatmap plot based on the correlation between bacteria phylum level and nutrient factors of fallen wood
3 讨 论
本研究显示,5 个分解等级倒木的理化性质在不同分解等级间均存在显著的差异。含水率从分解开始到后期依次降低,这与前人的研究结果一致[19-20],可能是由于倒木分解的进行,倒木密度变小,孔隙度增大吸水量增加,从而导致倒木的含水率增加。全氮的质量分数从分解初期到分解末期逐渐升高,总碳的质量分数从分解初期到分解末期逐渐降低,这与辛魏巍等[19]、Harmon 等[21]、马豪霞等[22-23]以及康玲等[24]的研究结果相类似,产生这样结果的可能原因包括降水、生物寄生[21]以及细菌的固氮等[25-26];随着分解的进行,不同类群的微生物侵入倒木,微生物的固氮作用、动物昆虫的粪便等增加了氮元素的输入,而微生物呼吸作用的增强增加了微生物对碳的分解,同时淋溶作用也加大了碳的损失[23]。碳氮比数值从分解初期到分级末期逐渐降低,这与Kielak 等[27]和马豪霞等[23]的研究认为随着分解的进行碳氮比降低的结果相同,碳氮比数值降低说明倒木中难分解的物质含量减少[28],在分解的过程中较容易分解的半纤维素、纤维素先被分解,留下较难分解的甲壳质和木质素进行后续分解[9],此外Kielak等[27]研究认为氮可能是分解过程中限制因子,这可能与微生物类群对碳和氮的需求不同而产生差异,这还需进一步的深入研究。
森林倒木分解首先从易分解的糖类开始到纤维素、半纤维素再到木质素等较难分解的物质[9],本研究中倒木的纤维素含量从Ⅰ级到Ⅴ级逐渐降低,木质素含量从Ⅰ级到Ⅴ级逐渐升高,这与都很多研究结果相类似[20,29]。针叶树倒木中的半纤维素成分主要是通过木材褐腐菌进行分解[9],分解过程通常伴随着纤维素的分解;本研究中半纤维素相对含量的变化趋势与纤维素相类似,从Ⅰ级到Ⅴ级逐渐降低。倒木基质分解的先后顺序、分解速率等与参与分解的微生物类群相关,参与分解的先驱微生物类群首先侵入,通常从易分解的物质开始分解,相比木质素来说纤维素和半纤维素较容易分解,所以随着分解的进行,纤维素和半纤维素的含量逐渐降低,木质素的相对含量升高。
倒木的分解受众多生物学和非生物学因素的影响,而同时倒木的径级、腐烂程度(分解等级)也能够影响微生物的多样性[29],同时为不同种类的微生物占据各自生态位提供生境[30]。有研究表明:木材上的微生物类群包括木材腐朽菌、木材软腐菌、木材变色菌、污染性霉菌、细菌和放线。细菌经常存在于木质部中,成为分解的先驱微生物;这部分细菌主要包括厚壁菌门的芽孢杆菌属Bacillus、假单孢杆菌属Pseudomonas、梭菌属Clostridium和欧氏杆菌属Erwinia,拟杆菌门的拟杆菌属Bacteroides,放线菌门的棒杆菌属Corynebacterium,变形菌门的黄单孢杆菌属Xanthomonas和不动杆菌属Acinetobacter,以及一些硫还原细菌和甲烷细菌等[9]。本研究中分解等级为Ⅰ级的倒木上以变形菌门、放线菌门、拟杆菌门和疣微菌门为主要优势类群,这与池玉杰等[9]研究的先驱微生物类群相类似。研究中不同分解等级倒木上的细菌优势菌门为变形菌和放线菌,这与Kielak 等[27]、Zhang 等[32]的研究结果相类似。此外在Ⅲ级分解等级的倒木上除变形菌门和放线菌门为优势菌门外,酸酐菌门的相对丰度为0.125 9,同为优势菌门;这与Kielak 等的研究结果相类似,这可能受腐烂木材低磷的环境所影响[33-34]。
Kielak 等在研究樟子松木材分解的过程中发现,分解的过程分为木材腐烂早期、中期和后期;在木材分解的早期细菌群落呈随机分布[35],中期和后期均符合生态位模型(Ecological niche modeling,ENM)。本研究中从Alpha 多样性指数上可以看出,各分解等级倒木细菌的Shannon 指数和Simpson 指数变化不大,而Ace 指数随着分解的进行,呈现先降低后升高的波动情况,且各级之间都存显著性差异;这样的结果表明倒木上细菌的物种的丰富度和均匀度(优势菌群的优势度)并未出现较大变化,说明这种变化并不是取代性或者颠覆性的,而是一个具有连续性的、发展性的演替过程,这与Prewitt 等[36]和Schowalter等[37]的针对倒木真菌的相关研究类似。此外,在Beta 多样性的PCoA 分析和环境因子关联分析中,也显示出倒木分解过程所被划分了3 个阶段,并且每个分解阶段以及各阶段的细菌类群都受倒木理化性质的影响。这也符合预测物种组成的变化不是随机的,而是与环境变量的变化有关的生态位理论。然而影响倒木分解的不仅仅是环境因子的变化,还可能包括其它生物类群(真菌、土壤动物、昆虫等)[9,38]、环境因素及其之间的互相作用等关系;况且本研究中只测定了有限数量的影响因子,所以影响分解的因素更复杂也更难以确定,还需进一步的研究。
4 结 论
大兴安岭寒温带兴安落叶松不同分解等级的倒木分解过程中,变形菌门和放线菌门为各分解等级倒木细菌的主要共有优势类群,其中放线菌门的相对丰度存在显著性差异。Alpha 多样性指数中Ace 指数先下降后升高且存在显著性差异;Beta 多样性和冗余分析将分解过程中的细菌分为3 组,其中优势菌门中的变形菌门与总氮含量极显著负相关,与碳氮比极显著正相关,放线菌门与半纤维素含量极显著正相关,与碳氮比显著正相关。碳氮比是大兴安岭寒温带兴安落叶松倒木分解过程中影响细菌组成和多样性的重要因素。
猜你喜欢
安徽农业科学(2022年20期)2022-11-11
科学技术与工程(2022年26期)2022-11-01
中国农学通报(2022年14期)2022-06-01
野生动物学报(2022年2期)2022-05-16
毛纺科技(2022年3期)2022-04-07
黑龙江大学自然科学学报(2022年1期)2022-03-29
计算机仿真(2021年7期)2021-11-17
疯狂英语·读写版(2020年11期)2020-12-21
电子制作(2019年24期)2019-02-23
中国知识产权(2018年12期)2018-12-29
