
稻蟹生态种养模式优质食味粳稻的稻米营养品质分析*
2021-04-15 00:00李跃东田春晖董立强孙富余
中国生态农业学报(中英文) 2021年4期
马 亮, 李跃东, 田春晖, 张 悦, 张 睿, 董立强, 孙富余**
稻蟹生态种养模式优质食味粳稻的稻米营养品质分析*
马 亮1, 李跃东1, 田春晖2, 张 悦1, 张 睿1, 董立强1, 孙富余2**
(1. 辽宁省水稻研究所 沈阳 110101; 2. 辽宁省农业科学院植物保护研究所 沈阳 110161)
本研究选用优质食味粳稻品种‘辽粳433’和‘五优稻4号’(稻花香2号)为材料, 设置了稻蟹生态种养(ecological rice-crab, ERC)和水稻单植(rice monoculture, RM)2个模式, 测定与稻米品质相关的蛋白质含量、直链淀粉含量和矿物质元素含量, 采用非靶向代谢组学方法分析两个模式之间与营养相关的差异代谢产物, 从多个方面分析稻蟹生态种养的稻米营养品质, 为稻蟹生态种养模式的应用推广提供理论依据。结果表明, 与RM相比, ERC的‘辽粳433’和‘五优稻4号’糙米蛋白质含量显著降低6.75%和10.11%, Fe含量显著增加, 为RM的2.03倍和4.99倍, Se含量显著增加, 为RM的3.11倍和5.74倍, Pb含量显著降低35.11%和44.35%。采用GC-MS检测到233种代谢产物, 利用偏最小二乘法(PLS-DA)分析发现, ERC和RM糙米的代谢产物差异显著, ERC诱导‘辽粳433’和‘五优稻4号’的4-氨基丁酸显著上调为RM的2.11倍和11.99倍, 抗氧化性物质α-生育酚、β-羟肉豆蔻酸以及奎宁酸分别显著上调为RM的1.62倍和2.22倍、12.64倍和20.48倍以及14.25倍和4.67倍, 己糖醇显著上调为RM的14.36倍和15.14倍, 半乳糖醇显著上调为RM的3.13倍和2.87倍。此外, ERC的‘辽粳433’产量降低286.0 kg∙km−2, ‘五优稻4号’产量增加344.5 kg∙km−2, 均与RM差异不显著。综上所述, 稻蟹生态种养模式在稳产的基础上可以增加优质食味粳稻稻米的营养价值。
稻蟹生态种养; 代谢组学; 营养品质; 产量; 矿物质元素; 食味品质
中国是世界上最早进行稻田养殖的国家[1], 稻田养殖的方式大体可划分为稻鱼、稻蟹、稻虾3种主要类型[2]。其中稻蟹种养模式在中国已经有40多年的历史[3], 是中国北方稻作区一种重要的种养模式[4], 在辽宁省约占水稻()种植面积的10%左右, 其中盘山县经过多年的稻蟹种养实践形成了具有特色的“盘山模式”[5]。
传统的水稻种植模式为了追求产量, 大量施用化肥和农药, 导致土壤结构和土质被破坏[3], 加大环境污染。稻蟹生态种养模式是根据稻蟹共生互利原理[6], 利用螃蟹在田间爬行防治病虫杂草, 利用螃蟹的排泄物为水稻提供天然营养物质, 减少了农药和化肥的使用量, 增加土壤有效矿物质元素[7], 土壤有机碳和碳水化合物的含量[8], 调节水稻生长的田间小气候[9]和微生物多样性格局[10]。因此, 稻蟹种养模式是一种保护土壤环境和增加经济效益的生态种养模式[11-13]。
稻田生态种养可以降低稻米的直链淀粉和蛋白质含量, 改善稻米品质[14-15], 但稻米营养品质内涵丰富, 还包括有益的矿物质元素(铁、硒等)、糖醇类物质、有机酸类物质、酚类化合物等。代谢组学(metabolomics)可以从生物学的角度对植物中的所有内源代谢物进行定性定量的研究。沈枫等[16]运用非靶向代谢组学进行了‘辽粳433’和‘越光’糙米代谢产物分析, 评价‘辽粳433’和‘越光’稻米营养价值。Heuberger等[17]利用代谢组学选择了营养价值更高和具有保健功能的水稻品种。以往的稻蟹种养模式对稻米品质的研究缺乏系统性, 本研究以优质食味粳稻‘辽粳433’和‘五优稻4号’(稻花香2号)为材料, 在分析稻蟹生态种养模式对水稻产量、蛋白质含量、矿物质元素含量影响的基础上, 采用非靶向代谢组学方法分析稻蟹生态种养模式与水稻单一种植模式之间稻米中与营养相关的代谢产物差异, 基于多个方面解析稻蟹生态种养模式的稻米营养品质, 为提升稻蟹生态种养模式的综合效益和模式推广提供理论依据。
1 材料与方法
1.1 试验材料
供试水稻品种为优质食味粳稻‘辽粳433’和‘五优稻4号’(稻花香2号), 种子来源于辽宁省水稻研究所。螃蟹为中华绒螯蟹(), 来源于盘山县河蟹研究所。
1.2 试验区域与试验设计
试验于2019年4—10月在辽宁省盘锦市盘山县坝墙子镇姜家村(40°40′~41°27′N、121°31~122°28′E)北方稻蟹种养示范基地进行。土壤基本理化性质为有机质16.8 g∙kg−1、全氮10.8 g∙kg−1、碱解氮 112.51 mg∙kg−1、速效磷 10.95 mg∙kg−1、速效钾131.5 mg∙kg−1、pH 6.8。
试验设置稻蟹生态种养模式(ERC)和水稻单一种植模式(RM)两个处理, 每个处理设置3次重复, 每个重复面积为200 m2, 重复之间有挡板进行隔离。水稻株距均为30 cm×20 cm。稻蟹生态种养模式(ERC)采用插秧时一次性侧深施用稻蟹田专用肥900 kg∙hm−2(含14%的N、5%的P和6%的K), 上一年的水稻秸秆还田和河蟹粪便为补充肥料。病虫草害主要依靠河蟹生物除草、理化诱控防治二化螟、减氮控病。螃蟹苗为5250只∙hm−2(80只∙kg−1), 于水稻返青后(6月10日)投放稻田。投放第2 d开始投喂螃蟹专用饲料, 投放量为螃蟹体重的3%~5%。螃蟹于9月15日起捕, 水稻于10月15日收获。为了贴近生产, 本研究选用农民传统种植模式, 即水稻单一种植模式(RM)为对照, 该模式是两次施肥, 底肥为750 kg∙hm−2(含250 g∙kg−1的N、150 g∙kg−1的P和120 g∙kg−1的K), 分蘖期追肥总量为尿素150 kg∙hm−2、硫酸铵150 kg∙hm−2、硫酸钾112.5 kg∙hm−2。分蘖盛期20%氯虫苯甲酰胺悬浮剂150 g∙hm−2进行二化螟防治, 孕穗期喷施75%三环唑可湿性粉剂375 g∙hm−2进行稻瘟病防治。
1.3 测定指标
10月15日进行水稻收获, 自然风干晾晒5 d进行稻米代谢产物检测, 自然风干15 d进行稻米品质和产量性状分析。
品质性状: 采用凯氏定氮法测定蛋白质含量[18], 利用620 nm比色测定直链淀粉含量[19]。同时测定糙米率、精米率和垩白率, 每个处理3次重复。
食味值: 采用日本佐竹satake食味仪进行稻米食味值测定, 每个处理测定3次重复。
微量元素: 采用火焰原子吸收光谱[20]对糙米中Ca、P、K、Mg、Fe、Zn、Se、Cr和Pb的含量进行测定, 测定在日立Z-2000型原子吸收分光光度计上完成, 每个处理设3次重复。
代谢产物: 采用气相色谱-质谱(GC-MS)方法进行糙米代谢产物检测, 具体步骤如下: 1.5 mL离心管中放入60 mg糙米冷冻磨碎的粉末样本、两颗钢珠、360 μL的冷甲醇和40 μL的内标(L-2-氯-苯丙氨酸, 0.3 mg∙mL–1, 甲醇配置), 在–80 ℃冰箱中放置2 min后研磨机研磨2 min, 超声提取30 min。加入200 μL的氯仿, 混匀, 再加入400 μL的水, 超声提取30 min。再12 000 r∙min–1, 4 ℃离心10 min, 取300 μL的上清液装入玻璃衍生瓶中。用冷冻浓缩离心干燥器挥发样品。向玻璃衍生小瓶中加入15 mg∙mL–1甲氧胺盐酸盐吡啶溶液80 μL, 涡旋振荡 2 min后, 于振荡培养箱中37 ℃进行肟化反应, 肟化时间为90 min。样品取出后再加入含1% TMCS的BSTFA衍生试剂80 μL和正己烷20 μL, 涡旋振荡 2 min后, 于70 ℃反应60 min。取出样本后, 在室温放置30 min, 使用安捷伦公司(Agilent Technologies Inc. CA, UAS)的7890B-5977A GC/MSD气质联用仪对样品进行代谢产物检测。采用MS-DIAL方法将GC-MS的原始数据进行定性分析, 采用内标法进行GC-MS的原始数据的归一化定量分析。每个处理6次重复。每6个样品中插入1个质控(quality control, QC)样本, 评价整个实验过程中系统质谱平台的稳定性。
产量性状: 每个重复5点取样, 测定经济性状, 包括有效穗数、一次枝梗数、穗粒数、结实率、千粒重和产量。
1.4 统计分析
运用Excel和SPSS 20.0等软件进行品种性状、微量元素含量和产量性状的方差分析。代谢产物数据通过在线软件进行PCA (principal components analyses)、PLS-DA (partial least-squares discriminant analysis)和差异代谢产物分析。
2 结果与分析
2.1 稻蟹生态种养模式对稻米食味品质的影响
与水稻单一种植模式(RM)相比, 稻蟹生态种养模式(ERC)降低了‘辽粳433’的垩白粒率、蛋白质含量和直链淀粉含量, 增加了糙米率、精米率和整精米率, 其中蛋白质含量显著降低6.75%, 其他性状差异不显著。ERC降低了‘五优稻4号’的垩白粒率、蛋白质含量和直链淀粉含量, 增加了糙米率、精米率和整精米率, 其中蛋白质含量显著降低10.50%, 垩白率显著降低了64.82%, 其他性状差异不显著(表1)。与RM相比, ‘辽粳433’和‘五优稻4号’的ERC食味值显著增加5.00%和4.28%, 说明稻蟹种养显著增加了稻米的食味水平。

表1 稻蟹生态种养模式对水稻品种‘辽粳433’和‘五优稻4号’碾磨品质和食味品质的影响
*和**分别表示稻蟹生态种养和水稻单一种植间在<0.05和<0.01水平差异显著。* and ** mean significant difference between ecological rice-crab and rice monoculture at<0.05 and<0.01 levels, respectively.
2.2 稻蟹生态种养模式对稻米矿物质元素含量的影响
与水稻单一种植模式(RM)相比, 稻蟹生态种养模式(ERC)极显著增加了‘辽粳433’和‘五优稻4号’稻米中元素Fe和Se含量, Fe含量为RM的2.03倍和4.99倍, Se含量为RM的3.11倍和5.74倍; ERC极显著(显著)降低了‘辽粳433’和‘五优稻4号’稻米中重金属元素Pb的含量, 分别降低35.11%和44.35%; 提高了P、K和Ca的含量, P含量分别提高8.47%和26.87%, K含量分别提高33.29%和24.24%, Ca含量分别提高27.96%和10.08%(表2)。

表2 稻蟹生态种养模式对水稻品种‘辽粳433’和‘五优稻4号’矿质元素含量的影响
*和**分别表示稻蟹生态种养和水稻单一种植间在<0.05和<0.01水平差异显著。* and ** mean significant difference between ecological rice-crab and rice monoculture at<0.05 and<0.01 levels, respectively.
2.3 稻蟹生态种养模式与水稻单一种植模式糙米代谢产物的差异
2.3.1 糙米代谢产物多变量分析
对‘辽粳433’和‘五优稻4号’的稻蟹生态种养模式(ERC)和水稻单一种植模式(RM)下糙米进行GC-MS检测, 均定性到233种代谢物, 其中, 有机酸59种, 碳水化合物48种, 氨基酸类37种, 核苷酸类11种, 次生代谢物36种, 醇类及酯类18种, 其他24种。
PCA分析可见(图1a), 所有的质控样品(QC)聚在一起, 表明所有检测具有良好的分析稳定性和试验重现性。PCA分析并未很好地将两个品种ERC和RM的代谢产物分离开, 分析原因可能是PCA分析不仅对同一个品种的两个处理之间代谢产物进行比较分析, 同时两个品种之间也进行了比较分析。将‘辽粳433’的ERC和RM以及‘五优稻4号’的ERC和RM分别进行PLS-DA分析, 结果表明, ERC和RM的糙米代谢产物分别位于主成分1的左右两边, 主成分1分别解释了所有变量的20.3%和25.4%(图1b-c)。模型累积解释率中发现,2值为0.9387和0.8281,2值为0.6620和0.5293, 表明模型的拟合准确性较好, 说明ERC显著改变了‘辽粳433’和‘五优稻4号’的代谢产物组成。
2.3.2 糙米中与营养相关的差异代谢产物分析
利用PLS-DA分析的变量权重值(VIP>1.0)进行代谢产物差异分析, ‘辽粳433’的稻蟹生态种养模式(ERC)与水稻单一种植模式(RM)之间有47种代谢产物差异显著, ‘五优稻4号’的ERC与RM之间有54种代谢产物差异显著(图2)。两组分析中共有变化趋势一样的代谢产物13种(图3), 包括4-氨基丁酸(GABA)、α-生育酚、β-羟肉豆蔻酸、半乳糖醇、葡萄糖-6-磷酸、谷氨酸、谷氨酰胺、己糖醇、甘露酸、N-乙酰-D-己糖胺、油酸、苯乙胺和奎宁酸。
ERC诱导糙米显著变化的13种代谢产物均与稻米品质有关, 其中氨基酸类物质3种, 谷氨酰胺显著上调为RM的2.97倍和2.01倍, 4-氨基丁酸显著上调为RM的2.11倍和11.99倍, 谷氨酸显著下调为RM的32%倍和8%倍; 糖醇类物质2种, 己糖醇显著上调为RM的14.36倍和15.14倍, 半乳糖醇显著上调为RM的3.13倍和2.87倍; 多酚类物质2种, 奎宁酸显著上调为RM的14.25倍和4.67倍, β-羟肉豆蔻酸显著上调为RM的12.64倍和20.48倍; 维生素类物质2种, α-生育酚显著上调为RM的1.62倍和2.22倍, 油酸显著降低为RM的0.52倍和0.11倍; 有机酸类物质2种, 葡萄糖-6-磷酸显著上调为RM的1.91倍和4.37倍, 甘露酸显著下调为RM的0.06倍和0.90倍; 胺类物质2种, 苯乙胺显著上调为RM的5.15倍和7.97倍, N-乙酰-D-己糖胺显著上调为RM的2.23倍和1.63倍。
A: ‘辽粳433’的ERC; B: ‘辽粳433’的RM; C: ‘五优稻4号’的ERC; D: ‘五优稻4号’的RM; QC: 质控样品。A: ‘Liaojing 433’ of ERC; B: ‘Liaojing 433’ of RM; C: ‘Wuyoudao No.4’ of ERC; D: ‘Wuyudao No.4’ of RM; QC: quality control sample.
A: ‘辽粳433’的ERC; B: ‘辽粳433’的RM; C: ‘五优稻4号’的ERC; D: ‘五优稻4号’的RM. A: ‘Liaojing 433’ of ERC; B: ‘Liaojing 433’ of RM; C: ‘Wuyoudao No.4’ of ERC; D: ‘Wuyudao No.4’ of RM.
A: ‘辽粳433’的ERC; B: ‘辽粳433’的RM; C: ‘五优稻4号’的ERC; D: ‘五优稻4号’的RM. A: ‘Liaojing 433’ of ERC; B: ‘Liaojing 433’ of RM; C: ‘Wuyoudao No.4’ of ERC; D: ‘Wuyudao No.4’ of RM.
2.4 稻蟹生态种养模式对‘辽粳433’和‘五优稻4号’产量构成因素的影响
产量构成因素分析表明(表3), 与水稻单一种植模式(RM)相比, ‘辽粳433’的稻蟹生态种养模式(ERC)降低了有效穗数和千粒重, 增加了枝梗数、穗实粒数和结实率, 其中枝梗数显著增加3.65%, 其他性状差异不显著。‘五优稻4号’的ERC降低了有效穗数, 增加了枝梗数、实粒数、千粒重和结实率, 显著(极显著)增加了5.85%的枝梗数和10.41%的千粒重, 其他性状差异不显著。与RM相比, ‘辽粳433’的ERC产量降低了286 kg·km–2, ‘五优稻4号’的ERC产量增加344.5 kg·km–2, 但差异不显著。
3 讨论
代谢组学是研究生物胁迫、水稻品种分类和水稻米质[21-22]的重要手段。可以通过PCA和PLS-DA分析进行数据中的相似性模式分析[23]和进行组之间的分离[24]。本研究通过PCA和PLS-DA分析发现, 稻蟹生态种养显著改变了糙米的代谢产物组成, 诱导糙米中多种与人类身体健康和抗氧化活性有关的代谢产物显著变化。己糖醇具有一定抗癌作用, 半乳糖醇具有润肠通便作用。GABA虽为非必需氨基酸, 但它被认为是一种具有生物活性的植物代谢物, 发芽糙米因含有较高的GABA被称为具有保健功能的大米[25]。维生素和酚类是有益的代谢物, 可以保护膜蛋白和DNA[26]。α-生育酚不仅是抗氧化物质[27]和脂溶性维生素[28], 同时在植物个体发育代谢过程中起着关键作用。从小分子角度分析表明, ERC显著改变了稻米中抗氧化活性物质, 增加了稻米的营养价值。
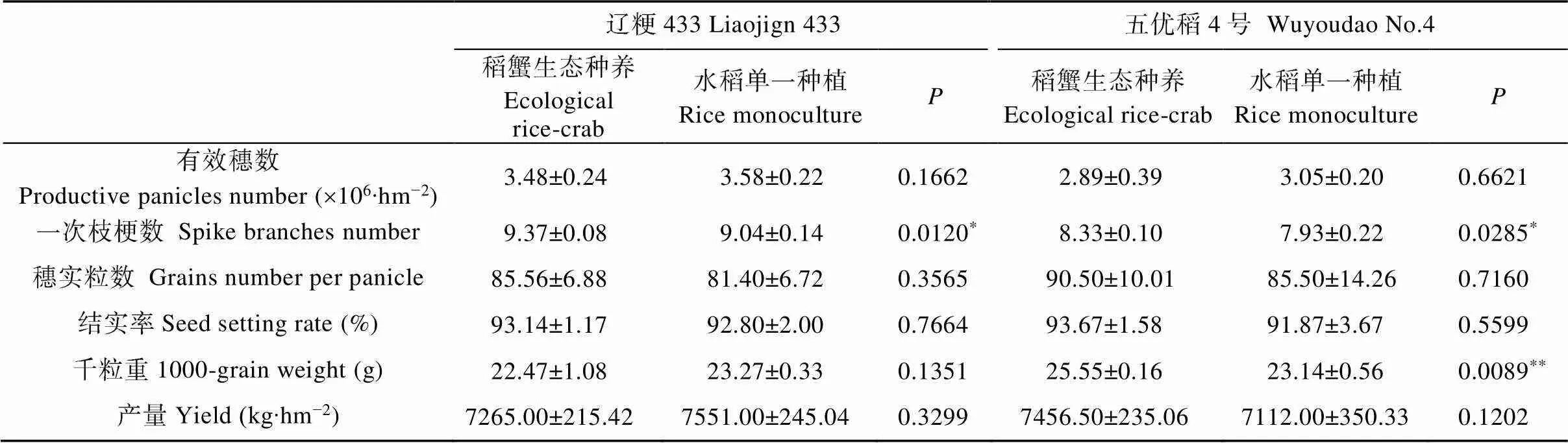
表3 稻蟹生态种养对水稻品种‘辽粳433’和‘五优稻4号’产量构成因素影响
*和**分别表示稻蟹生态种养和水稻单一种植间在<0.05和<0.01水平差异显著。* and ** mean significant difference between ecological rice-crab and rice monoculture at<0.05 and<0.01 levels, respectively.
微量元素含量是评价食品食用安全的一项重要指标, 稻米中Fe含量可以通过转基因技术得以提高[29], 富硒大米因其营养价值较高备受人们瞩目[30]。本研究发现, 稻蟹生态种养增加了‘辽粳433’和‘五优稻4号’稻米中有益矿物质含量, Fe和Se含量成倍增加。分析原因, 一方面可能是河蟹在稻田爬行增加水稻根系从土壤中吸收有效态的矿物质元素的量[10]; 另一方面, 葡萄糖-6-磷酸是植酸中肌醇环唯一来源[31], 代谢组学分析发现稻蟹生态种养的‘辽粳433’和‘五优稻4号’的葡萄糖-6-磷酸显著上调为单一种植的2.23倍和1.63, 抑制稻米中植酸含量, 降低了与植酸络合的Fe和Zn等金属离子, 增加了稻米中这些矿物质元素的含量。具体原因有待进一步分析。稻蟹生态种养和水稻单一种植模式下稻米中Pb和Cd的含量均符合食品安全国家标准[32], 但是, 稻蟹生态种养显著降低了Pb含量, 增加了的稻米食用安全性。
稻米品质和产量除了和自身遗传因素有关, 还与外界环境条件和栽培措施有一定关系。氮肥施用方法和施用量在一定程度上影响水稻的品质。张欣等[33]的研究结果表明, 与常规施肥相比, 施用猪粪有机肥与生物肥能显著提升稻米的食味品质。本研究结果也证明这一点, 稻蟹种养模式显著提高稻米的食味品质和营养品质, 与该模式为水稻提供的肥料以生物有机肥和螃蟹粪便为主有一定关系。与水稻单一种植相比, ‘辽粳433’和‘五优稻4号’稻蟹生态种养可以达到稳产效果, 分析原因一方面是由于螃蟹取食减少水稻无效分蘖, 合理调节植株氮素分配并促进稻粒氮素吸收; 另一方面是稻蟹生态种养模式利用一次性侧深施有机稻蟹专用肥, 减少施用化肥产生氨态氮, 同时, 螃蟹取食稻田杂草、水生生物、专用配合饲料等过腹粪便还田, 满足了水稻生育期对养分的需求, 提高水稻各器官氮素运转酶活性[34]和氮肥利用率[35]。该模式充分利用稻田种养生态系统中有利因子, 减施肥药, 最终实现水稻稳产和增加品质的目的。
4 结论
稻蟹生态种养模式在保证优质食味粳稻‘辽粳433’和‘五优稻4号’产量的前提下, 降低稻米蛋白质含量, 增加了稻米食味值; 显著增加稻米中Fe含量, 降低了有毒物质Pb含量, 提高了稻米的食用安全性;增加了有益物质GABA和α-生育酚含量, 提高了稻米的抗氧化能力。因此, 稻蟹生态种养模式具有重要的生态意义、经济意义和推广价值。
[1] 王强盛, 王晓莹, 杭玉浩, 等. 稻田综合种养结合模式及生态效应[J]. 中国农学通报, 2019, 35(8): 46–51 WANG Q S, WANG X Y, HANG Y H, et al. Combination modes and ecological effects of planting-breeding ecosystem in rice field[J]. Chinese Agricultural Science Bulletin, 2019, 35(8): 46–51
[2] 管卫兵, 刘凯, 石伟, 等. 稻渔综合种养的科学范式[J]. 生态学报, 2020, 40(16): 5451–5464 GUAN W B, LIU K, SHI W, et al. Scientific paradigm of integrated farming of rice and fish[J]. Acta Ecologica Sinica, 2020, 40(16): 5451–5464
[3] 王昂, 王武, 马旭洲. 稻蟹共作模式的发展历程和前景展望[J]. 吉林农业科学, 2013, 38(3): 89–92 WANG A, WANG W, MA X Z. Development and prospects on rice-crab model[J]. Journal of Jilin Agricultural Sciences, 2013, 38(3): 89–92
[4] 王昂, 戴丹超, 马旭洲, 等. 稻蟹共作模式对土壤微生物量氮和酶活性的影响[J]. 江苏农业学报, 2019, 35(1): 76–84 WANG A, DAI D C, MA X Z, et al. Effects of rice-crab culture system on soil microbial biomass nitrogen and soil enzymes activities[J]. Jiangsu Journal of Agricultural Sciences, 2019, 35(1): 76–84
[5] 胡宗云, 杨培民, 李文宽, 等. 稻蟹共作模式下稻蟹产出与土壤理化性质的研究[J]. 河北渔业, 2015, (5): 5–7 HU Z Y, YANG P M, LI W K, et al. Study on the output of rice crab and the physical and chemical properties of soil under the mode of crab co cropping[J]. Hebei Fisheries, 2015, (5): 5–7
[6] 陈玥, 牛世伟, 邹晓锦, 等. 稻蟹联合种养对稻田生态环境的影响[J]. 辽宁农业科学, 2018, (3): 30–34 CHEN Y, NIU S W, ZOU X J, et al. Effect of rice-crab co-culture on environment of rice field[J]. Liaoning Agricultural Sciences, 2018, (3): 30–34
[7] 林利红, 王延平, 王帅. 稻田养蟹对水稻土壤理化性质的影响研究[J]. 辽宁农业科学, 2016, (3): 43–47 LIN L H, WANG Y P, WANG S. Effects of rice-crab model on physical and chemical properties of rice soil[J]. Liaoning Agricultural Sciences, 2016, (3): 43–47
[8] YAN Y, LIU M D, YANG D, et al. Effect of different rice-crab coculture modes on soil carbohydrates[J]. Journal of Integrative Agriculture, 2014, 13(3): 641–647
[9] 陈灿, 黄璜, 丁姣龙, 等. 稻田综合种养稻米质量特性形成及调优机理探究[J]. 作物研究, 2019, 33(5): 392–397 CHEN C, HUANG H, DING J L, et al. Study on the formation and optimization mechanism of rice quality characteristics in comprehensive planting and breeding model of rice field[J]. Crop Research, 2019, 33(5): 392–397
[10] 马晓慧, 车喜庆, 王井士, 等. 稻蟹共作与常规稻田蜘蛛群落组成及多样性分析[J]. 中国生态农业学报(中英文), 2019, 27(8): 1157–1162 MA X H, CHE X Q, WANF J S, et al. The structure of spider communities in crab paddies and conventional paddies[J]. Chinese Journal of Eco-Agriculture, 2019, 27(8): 1157–1162
[11] 汪倩, 胡庚东, 宋超, 等. 基于Ecopath评估蟹-稻复合生态系统营养结构和能量流动[J]. 生态学报, 2020, 40(14): 4852–4862 WANG Q, HU G D, SONG C, et al. Evaluation of nutritional structure and energy flow of crab-rice complex ecosystem based on an Ecopath model[J]. Acta Ecologica Sinica, 2020, 40(14): 4852–4862
[12] 闫志利, 林瑞敏, 牛俊义, 等. 我国稻蟹共作技术研究的现状与前景展望[J]. 北方水稻, 2008, 38(2): 5–8 YAN Z L, LIN R M, NIU J Y, et al. Current status and prospectives of rice-crab production technique research in China[J]. North Rice, 2008, 38(2): 5–8
[13] 陈飞星, 张增杰. 稻田养蟹模式的生态经济分析[J]. 应用生态学报, 2002, 13(3): 323–326 CHEN F X, ZHANG Z J. Ecological economic analysis of a rice-crab model[J]. Chinese Journal of Applied Ecology, 2002, 13(3): 323–326
[14] 安辉, 刘鸣达, 王厚鑫, 等. 不同稻蟹生产模式对稻蟹产量和稻米品质的影响[J]. 核农学报, 2012, 26(3): 581–586 AN H, LIU M D, WANG H X, et al. Effects of different rice-crab production models on rice-crab yield and quality[J]. Journal of Nuclear Agricultural Sciences, 2012, 26(3): 581–586
[15] 陈灿, 黄璜, 郑华斌, 等. 稻田不同生态种养模式对稻米品质的影响[J]. 中国稻米, 2015, 21(2): 17–19 CHEN C, HUANG H, ZHENG H B, et al. Effects of different mode of ecological planting and raising on rice quality[J]. China Rice, 2015, 21(2): 17–19
[16] 沈枫, 蒋洪波, 刘博, 等. 优质食味粳稻辽粳433和越光糙米代谢产物差异分析[J]. 中国水稻科学, 2020, 34(4): 359–367 SHEN F, JIANG H B, LIU B, et al. Difference of metabolites in brown rice between Liaojing 433 and Koshihikari with good eating quality[J]. Chinese Journal of Rice Science, 2020, 34(4): 359–367
[17] HEUBERGER A L, LEWIS M R, CHEN M H, et al. Metabolomic and functional genomic analyses reveal varietal differences in bioactive compounds of cooked rice[J]. PLoS One, 2010, 5(9): e12915
[18] 张杰, 郑蕾娜, 蔡跃, 等. 稻米淀粉RVA谱特征值与直链淀粉、蛋白含量的相关性及QTL定位分析[J]. 中国水稻科学, 2017, 31(1): 31–39 ZHANG J, ZHENG L N, CAI Y, et al. Correlation analysis and QTL mapping for starch RVA profile properties and amylose and protein contents in rice[J]. Chinese Journal of Rice Science, 2017, 31(1): 31–39
[19] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 15683–2008 大米直链淀粉含量的测定[S]. 北京: 中国标准出版社, 2009 General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of China. GB/T 15683–2008 Rice—Determination of Amylose Content[S]. Beijing: China Standard Press, 2009
[20] 张标金, 罗林广, 魏益华, 等. 镉锰互作对水稻幼苗镉和矿物质积累的影响[J]. 江西农业学报, 2015, 27(4): 8–11 ZHANG B J, LUO L G, WEI Y H, et al. Effects of cadmium and manganese interaction on cadmium and mineral elements accumulation in rice seedlings[J]. Acta Agriculturae Jiangxi, 2015, 27(4): 8–11
[21] 雷刚, 黄英金. 代谢组学在水稻研究中的应用进展[J]. 中国农业科技导报, 2017, 19(7): 27–35 LEI G, HUANG Y J. Application progress of metabolomics in rice research[J]. Journal of Agricultural Science and Technology, 2017, 19(7): 27–35
[22] CALINGACION M N, BOUALAPHANH C, DAYGON V D, et al. A genomics and multi-platform metabolomics approach to identify new traits of rice quality in traditional and improved varieties[J]. Metabolomics, 2012, 8(5): 771–783
[23] ALONSO A, MARSAL S, JULIÀ A. Analytical methods in untargeted metabolomics: State of the art in 2015[J]. Frontiers in Bioengineering and Biotechnology, 2015, 3: 23
[24] JUNG Y, AHN Y G, KIM H K, et al. Characterization ofspecies using1H NMR- and GC-MS-based metabolite profiling[J]. Analyst, 2011, 136(20): 4222–4231
[25] 丁俊胄, 杨特武, 周强, 等. 厌氧胁迫对发芽糙米中γ-氨基丁酸含量变化的影响[J]. 中国粮油学报, 2015, 30(2): 6–10 DING J Z, YANG T W, ZHOU Q, et al. γ-Aminobutyric acid content of brown rice induced by hypoxia stress during germination[J]. Journal of the Chinese Cereals and Oils Association, 2015, 30(2): 6–10
[26] SZETO Y T, KWOK T C Y, BENZIE I F F. Effects of a long-term vegetarian diet on biomarkers of antioxidant status and cardiovascular disease risk[J]. Nutrition, 2004, 20(10): 863–866
[27] SADIQ M, AKRAM N A, ASHRAF M, et al. Alpha- tocopherol-induced regulation of growth and metabolism in plants under non-stress and stress conditions[J]. Journal of Plant Growth Regulation, 2019, 38(4): 1325–1340
[28] TIGU F, ZHANG J L, LIU G X, et al. A highly active pantothenate synthetase fromenables the production of D-pantothenic acid with high productivity[J]. Applied Microbiology and Biotechnology, 2018, 102(14): 6039–6046
[29] 刘巧泉, 姚泉洪, 王红梅, 等. 转基因水稻胚乳中表达铁结合蛋白提高稻米铁含量[J]. 遗传学报, 2004, 31(5): 518–524 LIU Q Q, YAO Q H, WANG H M, et al. Endosperm-specific expression of the ferritin gene in transgenic rice (L.) results in increased iron content of milling rice[J]. Acta Genetica Sinica, 2004, 31(5): 518–524
[30] 姜超强, 沈嘉, 祖朝龙. 水稻对天然富硒土壤硒的吸收及转运[J]. 应用生态学报, 2015, 26(3): 809–816 JIANG C Q, SHEN J, ZU C L. Selenium uptake and transport of rice under different se-enriched natural soils[J]. Chinese Journal of Applied Ecology, 2015, 26(3): 809–816
[31] RABOY V, GERBASI P. Genetics of-inositol phosphate synthesis and accumulation[J]. Subcellular Biochemistry, 1996, 26: 257–285
[32] 国家卫生和计划生育委员会, 国家食品药品监督管理总局. GB 2762–2017 食品安全国家标准食品中污染物限量[S]. 北京: 中国标准出版社, 2017 National Health and Family Planning Commission of the People’s Republic of China, National Medical Products Administration. GB 2762–2017 National Food Safety Standard of the People’s Republic of China[S]. Beijing: China Standard Press, 2017
[33] 张欣, 施利利, 刘晓宇, 等. 不同施肥处理对水稻产量、食味品质及蛋白质组分的影响[J]. 中国农学通报, 2010, 26(4): 104–108 ZHANG X, SHI L L, LIU X Y, et al. Effect of different fertilizer treatments on rice yield, grain quality and protein fraction content[J]. Chinese Agricultural Science Bulletin, 2010, 26(4): 104–108
[34] 陈香碧, 胡亚军, 秦红灵, 等. 稻作系统有机肥替代部分化肥的土壤氮循环特征及增产机制[J]. 应用生态学报, 2020, 31(3): 1033–1042CHEN X B, HU Y J, QIN H L, et al. Characteristics of soil nitrogen cycle and mechanisms underlying the increase in rice yield with partial substitution of mineral fertilizers with organic manure in a paddy ecosystem: A review[J]. Chinese Journal of Applied Ecology, 2020, 31(3): 1033–1042
[35] 徐敏, 马旭洲, 王武. 稻蟹共生系统水稻栽培模式对水稻和河蟹的影响[J]. 中国农业科学, 2014, 47(9): 1828–1835 XU M, MA X Z, WANG W. Effects of different cultivation patterns on rice yield and crab in rice-crab culture system[J]. Scientia Agricultura Sinica, 2014, 47(9): 1828–1835
Nutritional quality of japonica rice with good taste quality in an ecological rice-crab mode*
MA Liang1, LI Yuedong1, TIAN Chunhui2, ZHANG Yue1, ZHANG Rui1, DONG Liqiang1, SUN Fuyu2**
(1. Liaoning Rice Research Institute, Shenyang 110101, China; 2. Institute of Plant Protection, Liaoning Academy of Agricultural Sciences, Shenyang 110161, China)
In this study, we investigated the effects of an ecological rice-crab mode on the nutritional value of rice using two cropping patterns, ecological rice-crab (ERC) and rice monoculture (RM). Two japonica rice cultivars with good taste quality, ‘Liaojing 433’ and ‘Wuyoudao No.4’ (Daohuaxiang No.2), were selected as the study materials, and the rice contents of protein, amylose, and mineral elements, which are related to rice quality, were measured. Differences in the nutrition-related metabolites of ERC and RM were analyzed using non-targeted metabolomics. This study aimed to provide a theoretical basis for the application and popularization of the ecological rice-crab mode. The results showed that compared with RM, the protein content of brown rice of ‘Liaojing 433’ and ‘Wuyoudao No.4’ under the ERC mode significantly decreased by 6.75% and 10.11%, respectively, and Pb content decreased by 35.11% and 44.35%, respectively. The Fe content significantly increased by 1.03 and 3.99 times, respectively, and the Se content increased by 2.11 and 4.74 times, respectively. Two hundred and thirty-three metabolites were detected by gas chromatography-mass spectrometry (GC-MS). Significant differences in the metabolites were found between ERC and RM brown rice by partial least-squares discriminant analysis (PLS-DA). Compared with RM, the content of 4-aminobutyric acid in ‘Liaojing 433’ and ‘Wuyoudao No.4’ under the ERC mode increased significantly by 1.11 and 10.99 times, respectively; and the contents of antioxidant substances, such as alpha-tocopherol, beta-hydroxamic acid, and quinic acid, significantly increased by 0.62 and 1.22 times, 11.64 and 19.48 times, and 13.25 and 3.67 times, respectively. The hexitol content increased by 13.36 and 14.14 times, respectively, and the galactitol content increased by 2.13 and 1.87 times, respectively. Rice yield of ‘Liaojing 433’ under the ERC mode decreased by 286.0 kg∙km−2, whereas that of ‘Wuyoudao No.4’ under the ERC mode increased by 344.5 kg∙km−2. However, there were no significant differences in rice yield between ERC and RM. In conclusion, the ERC mode can increase the nutritional value of high-quality japonica rice without reducing its production.
Ecological rice-crab mode; Metabolomics; Nutritional quality;Yield; Mineral elements;Taste quality
10.13930/j.cnki.cjea.200441
马亮, 李跃东, 田春晖, 张悦, 张睿, 董立强, 孙富余. 稻蟹生态种养模式优质食味粳稻的稻米营养品质分析[J]. 中国生态农业学报(中英文), 2021, 29(4): 716-724
MA L, LI Y D, TIAN C H, ZHANG Y, ZHANG R, DONG L Q, SUN F Y. Nutritional quality of japonica rice with good taste quality in an ecological rice-crab mode[J]. Chinese Journal of Eco-Agriculture, 2021, 29(4): 716-724
S511.4

* 国家重点研发计划项目(2018YFD0200200)和辽宁省农业科学院院长(青年)基金(2020QN2411)资助
孙富余, 主要从事农作物病虫害绿色防控及食品安全控害生产技术研究。E-mail: laassfy@163.com
马亮, 主要从事水稻病虫害防控和优质栽培技术研究。E-mail: malhd@126.com
2020-06-11
2020-10-30
* This work was supported by the National Key R&D Program of China (2018YFD0200200) and the Dean Fund of Liaoning Academy of Agricultural Sciences (2020QN2411).
, E-mail: laassfy@163.com
Jun. 11, 2020;
Oct. 30, 2020
猜你喜欢
美食(2022年5期)2022-05-07
中国农业科学(2022年7期)2022-04-14
早期教育(家庭教育)(2021年11期)2021-12-17
少儿科学周刊·儿童版(2021年21期)2021-12-11
中国农业科学(2021年6期)2021-03-25
红楼梦学刊(2020年2期)2020-02-06
亲子(2014年4期)2014-05-10
食品工业科技(2014年13期)2014-03-11
饮食科学(2009年5期)2009-05-14
祝你幸福·午后版(2008年10期)2008-09-20
