
A comparison of zooplankton assemblages in Nansi Lake and Hongze Lake, potential influences of the East Route of the South-to-North Water Transfer Project, China*
2021-04-14 05:53KandathilRadhakrishnanDIVYAShashaZHAOYushunCHENFeiCHENG
Kandathil Radhakrishnan DIVYA , Shasha ZHAO, Yushun CHEN , Fei CHENG , ,
Lei ZHANG2, Jiao QIN1, Thundiparambil Sathrajith ARUNJITH 1, V. Bjorn SCHMIDT1,Songguang XIE1,2
1 Key Laboratory of Aquatic Biodiversity and Conservation of Chinese Academy of Sciences, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
2 Key Laboratory of Sustainable Fisheries and Environmental Protection for Lakes of Northern Jiangsu, Huai’an Research Center, Institute of Hydrobiology, Chinese Academy of Sciences, Huai’an 223002, China
Abstract Nansi Lake and Hongze Lake are both water storage lakes along the East Route of the South-to-North Water Transfer project (ESNT). Frequent changes in hydrologic properties are responsible factors for controlling the zooplankton community assemblages in both lakes, so we studied the possible influence of water transfer and environmental factors on zooplankton community structure and abundance.Zooplankton assemblages were investigated seasonally for one year in both lakes; a total of 133 and 122 zooplankton taxa were identified in Nansi Lake and Hongze Lake, respectively. The most dominant rotifer species were littoral, e.g., Keratella tecta, Keratella valga and Lecane lunaris in Nansi Lake and Brachionus angularis, Brachionus forficula and Polyarthra vulgaris in Hongze Lake. Comparatively, Nansi Lake had a higher Shannon-Wiener diversity index value (5.13), while Hongze Lake had a higher species richness index (4.21). The average number of zooplankton across seasons in Nansi Lake (protozoa: 774±63 ind./L,rotifers: 4 817±212 ind./L, cladocerans: 896±14 ind./L, copepod: 435±42 ind./L) was comparatively lower than Hongze Lake (protozoa: 1 238±63 ind./L, rotifers: 6 576±112 ind./L, cladocerans: 1 013±20 ind./L,copepod: 534±25 ind./L). Canonical correspondence analysis identified diff ering environmental gradients that were most responsible for influencing zooplankton communities in the two lakes (Hongze Lake: NH 4-N,total nitrogen, transparency and pH; Nansi: pH, temperature and total phosphorus). Frequent changes related to water transfer in lakes favoured the diversity of rotifers and protozoa communities. Zooplankton habitat preference, changes in community structure and opportunistic peaks and extinction of certain taxa were also observed in the study lakes.
Keyword: zooplankton assemblage; water transfer; Hongze Lake; Nansi Lake
1 INTRODUCTION
Inter-basin water transfer has been applied around the world to alleviate water shortages in waterdeficient areas. The difference in hydrologic properties between donor and recipient systems influences both abiotic (physical and chemical) and biotic (species interactions, habitat, species composition of plants and animals) features of the connected ecosystems(Gruberts et al., 2007; Kufel and Leśniczuk, 2014;Winemiller et al., 2015). Several studies have shown positive eff ects of water transfers, through decreasing phytoplankton concentrations and improvement of water quality in downstream reservoirs (Padisák et al., 2000; Hu et al., 2010; Zhai et al., 2010). Other studies have reported negative implications of water transfer such as fluctuations of the nutrient influx in recipient systems, including a significant increase in silica, total iron and chlorophyll a content (Matsumura-Tundisi and Tundisi, 2005; Fornarelli and Antenucci,2011). Water transfer can also negatively aff ect fish and invertebrate communities through increasing the likelihood of species invasions, possibly through the transfer of alien species and toxic cyanophytes into recipient systems (Matthews et al., 1996; Snaddon and Davies, 1998).
Zooplankton communities provide an ideal indicator of human disturbances in lake monitoring programs (Stemberger et al., 2001; Dodson et al.,2005). In lacustrine habitats, zooplankton community structure and seasonal dynamics are determined by a variety of factors such as geographic location, the lake area, water depth, water current velocity, and presence of macrophytes. Lake morphological characteristics such as surface area and shape influence species composition and richness of zooplankton, mainly based on the available limnetic and littoral habitats in the lake (Fryer, 1985; Gasith and Gafny, 1990; Dodson, 1991, 1992; Karatayev et al., 2005; Padial et al., 2009). Additionally, fluctuations in hydrological properties can either directly or indirectly aff ect biological communities such as zooplankton (Wolcox and Meeker, 1992; Betsill and Van Den Avyle, 1994; Poff et al., 1997; Gruberts et al., 2007; Leira and Cantonati, 2008). Biotic factors(both bottom-up and top-down factors) (Carpenter et al., 1985; McQueen et al., 1986; Northcote, 1988;Vanni, 1988) and abiotic factors like water chemistry can also act as driving forces for structuring zooplankton communities (Johannsson et al., 1991;Pourriot et al., 1994).
In recent centuries, many aquatic sources have been strongly transformed as a result ofintensive engineering and water diversion, and these kinds of activities caused deterioration of biodiversity. The zooplankton assemblages, as well as the other biotic communities that inhabit the aquatic ecosystems,constitute an important element of the food chain, and are eff ective indicators of the trophic conditions.Moreover, zooplankton communities are sensitive to anthropogenic impacts and their study may be useful in the prediction oflong-term changes in lake ecosystems. To understand the ecological function in lakes, suffi cient knowledge is required for all biotic elements, including zooplankton. The present study aims to identify the trends of zooplankton diversity and distribution in Nansi Lake and Hongze Lake and their relationships with environmental factors(physical and chemical features of the water). Two questions were answered: How did the water transfer aff ect the zooplankton community structure and distribution in water storage lakes (Nansi anid Hongze) along the East Route of the South-to-North Water Transfer Project (ESNT)? Which environmental factors were responsible for temporal changes of zooplankton community in both lakes?
2 MATERIAL AND METHOD
2.1 Study area
Nansi Lake and Hongze Lake are water storage lakes along the ESNT. The ESNT diverts water from the lower reaches of the Changjiang (Yangtze) River at Jiangdu near Yangzhou, Jiangsu Province, to the North China Plain, using the Grand Canal as the main conveyance channel. The ESNT also intersects and passes through Nansi and Hongze Lake during the transfer process. The ESNT was expected to increase the water levels of Nansi and Hongze Lake by 0.5 m,and the annual water exchange rate was expected to be 1.2 times after it became fully operational (Zhang,2009). The first full East Route transfer was done in 2013; during normal operation water passes through Hongze Lake and is conveyed to Nansi Lake. Nansi Lake has a dividing dam between the upper and lower lakes, and the water collects first in lower Nansi Lake(Weishan) and then is lifted and moved to upper Nansi Lake. Comparatively, water depth is higher in Hongze Lake than in Nansi Lake. Additionally, water flow may cause there to be less macrophytes in Hongze Lake due to stronger water currents during transfer,while in Nansi Lake water is first collected at the dam leading to comparatively weaker currents and more extensive macrophyte coverage.
Nansi Lake (34°36′N, 117°12′E) (Fig.1a) is located in the north of the Huaihe River basin in Shandong Province, China. It is a relatively large, shallow lake,with a total surface area (combined upper and lower lakes) of 1 266 km2and an average depth of 1.5 m.According to lake morphology, most of the area of Nansi Lake is covered by vegetation, providing more habitat heterogeneity for species colonization.
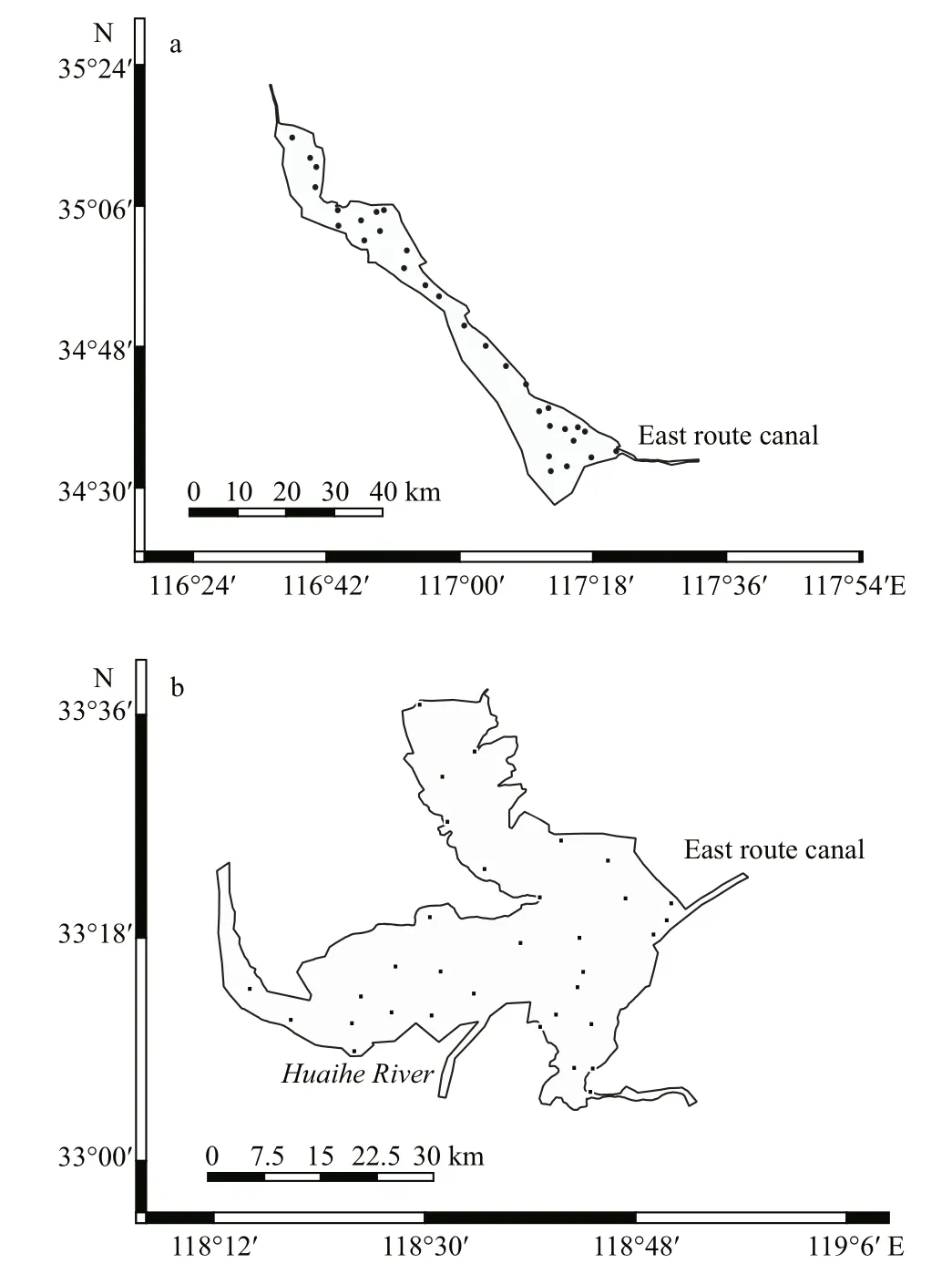
Fig.1 Map of the Nansi Lake (a) and Hongze Lake (b)showing the distribution of sampling stations
Hongze Lake (33°18′27″N 118°42′36″E) (Fig.1b)is the fourth largest freshwater lake in China, in the middle reach of the Huaihe River in Jiangsu Province with a total surface area of 1 960 km2. The mean water depth of the lake is 1.77 m, with the deepest sections being around 4.37 m (Chu, 2001). The water volume is about 27.9×108m3. The average elevation of the lake basin is around 4-8 m higher than the eastern plain of China. Approximately 73% of the lake water is contributed by the inflowing Huaihe River (Yang,1993). Hongze Lake is a transitional lake where water levels often undergo large fluctuations both annually and seasonally (Wang and Chen, 1999). Overall, the macrophyte coverage in Hongze Lake is very low,although there remains some coverage in the northeast and southwest sections. Therefore, comparatively, the open water area of Hongze Lake is higher than Nansi Lake (Liu et al., 2009; Lin et al., 2017).
2.2 Physico-chemical environmental analysis
Physico-chemical environmental variables were measured for each sample site during zooplankton sampling. Water temperature (°C), pH, and dissolved oxygen (DO) (mg/L) were measured in-situ using a Yellow Springs Instrument (YSI proplus probe).Transparency (m) and water depth (m) were measured with a Secchi disk. Water samples (from 0.5 m below the surface) were also obtained during zooplankton sampling. Water samples were analyzed to determine total phosphorus (TP, stannous chloride method, mg/L), nitrate nitrogen (NO3-N, nitrate selective method, mg/L), chemical oxygen demand(COD, open reflux method, mg/L) (APHA, 1992),and total nitrogen (TN, potassium persulfate digestion and ultraviolet spectrophotometry method,mg/L), ammonia nitrogen (NH4-N, Nessler’s reagent spectrophotometry method, mg/L) (Huang et al.,1999).
2.3 Zooplankton sampling and analysis
Zooplankton was sampled at 31 sampling stations in Nansi Lake and 33 sampling stations in Hongze Lake covering the whole area for both lakes (Fig.1).Sampling was carried out seasonally (spring, summer,autumn and winter) in April, July, and October of 2015, and January 2016. Sampling was carried out both qualitatively and quantitatively. Qualitative samples were collected using conical nets of 23-μm mesh for protozoa and rotifers (fixed at 1% Lugol’s solution) and of 64-μm mesh for crustaceans (fixed at 4% formalin) by horizontally towing at the surface at approximately 0.5 m depth, at a speed of 1-1.5 m/s,for approximately 5 min. Zooplankton were identified in the lab to the lowest possible taxonomic level,usually to species or genus (Chiang and Du, 1979;Shen and Song, 1979; Tai and Chen, 1979; Tai and Song, 1979; Shen, 1983; Shiel, 1995) at 10×, 40×,and 100× magnifications using an Olympus compound microscope.
Quantitative sampling was performed using a 5-L modified Patalas water sampler. For quantitative samples of smaller organisms (e.g., protozoa and rotifers, nets of 23 μm mesh), 5 L of water was filtered and fixed in 1% Lugol’s solution. For crustaceans(nets of 23-μm mesh), 20 L of water was filtered and preserved with 4% formalin. Zooplankton (rotifers,protozoans, cladocerans and copepods) samples were counted using Sedgewick-Rafter counting chambers.The enumeration of specimens in the samples was done using subsamples of 0.1 mL for protozoans and rotifers and 1 mL for crustaceans. Dominant species were calculated as those species occurring with more than 10% of the total individuals of a sample according to (Patalas, 1971). Habitat preference of zooplankton species was classified as limnetic, littoral, or limnetic/littoral (Tai and Chen, 1979; Shen, 1983; Smirnov and Timms, 1983; Koste and Tobias, 1987; Koste and Shiel, 1991).
2.4 Statistical analysis
The datasets were checked with the Shapiro-Wilk test, and then, if necessary, square root (physicochemical environmental variables) or log ( x+1)(zooplankton density) transformed to achieve normality. To analyze differences between the lakes, a two-way analysis of variance (ANOVA) was performed for zooplankton density and a Friedman test was performed for physico-chemical parameters.All the above statistical analyses were conducted using SPSS, Version 20 (IBM Chicago, IL). Analyses were also conducted to calculate species richness,evenness and Shannon-Wiener diversity indices of zooplankton (rotifers, protozoans, cladocerans, and copepods) of the two lakes using software package Primer 5 (Clarke and Warwick, 2001).
Finally, to elucidate the relationship between the zooplankton composition and environmental conditions,a canonical correspondence analysis (CCA) was conducted for each lake (Ter Braak and Verdonschot,1995). Species abundances were square root-transformed and down-weighted for rare species, following the method of (Lepš and Šmilauer, 2003). The forward selection of CCA, which is analogous to stepwise multiple regression, was used to determine the significance ( P <0.05) of environmental variables that could explain the variation in the species data. The significance of these variables was assessed using Monte Carlo permutation tests (with 999 unrestricted permutations). All the ordinations were performed using CANOCO version 4.5 (Ter Braak and Smilauer, 2002).
3 RESULT
3.1 Zooplankton community structure and density
Total of 133 zooplankton species were identified in Nansi Lake, and 122 were identified in Hongze Lake.Zooplankton species (either limnetic or littoral)appeared in the category of sporadic (<10%) in both lakes (Table 1). Rotifers contributed as the major grouping for the total zooplankton diversity with afrequency of occurrence of more than 50%. In contrast with diversity patterns of protozoa and Rotifera,crustacean zooplankton in the two lakes had low species diversity. In Nansi Lake and Hongze Lake,rotifers and protozoans contributed more to the total zooplankton density ( P <0.05), (Fig.2).
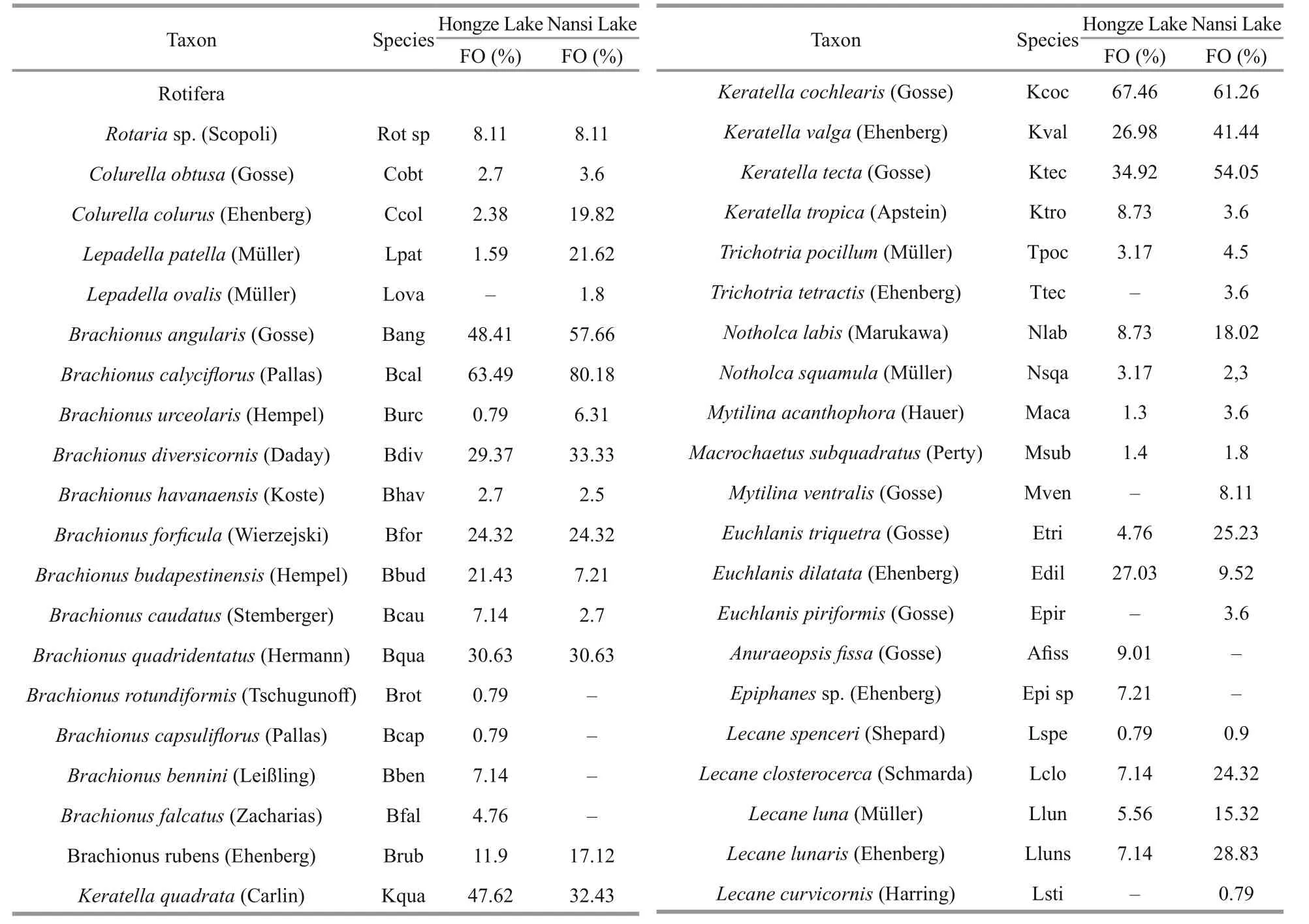
Table 1 Zooplankton taxa recorded in Nansi Lake and Hongze Lake with frequency of occurrence (FO) (%) of species
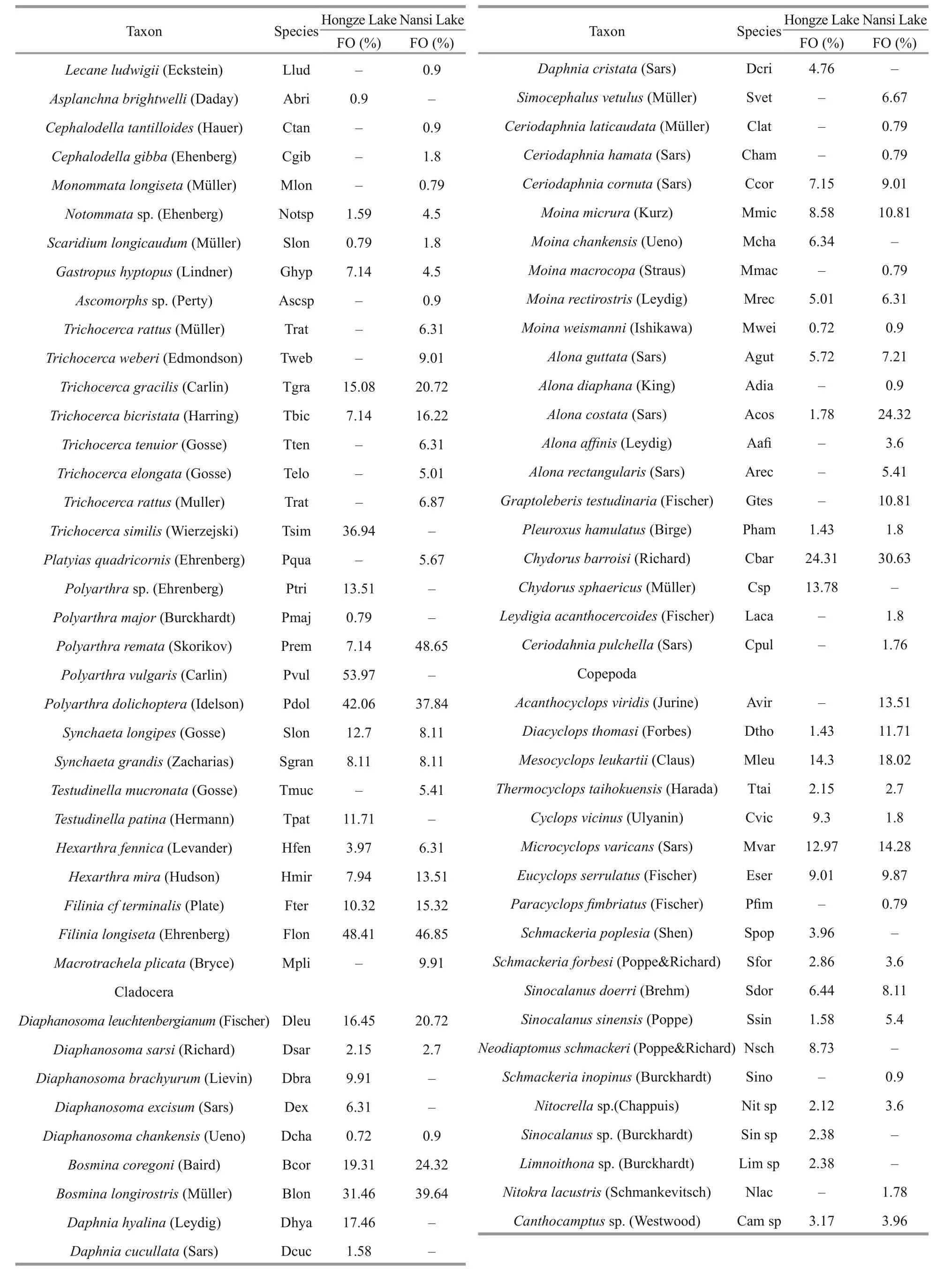
Table 1 Continued
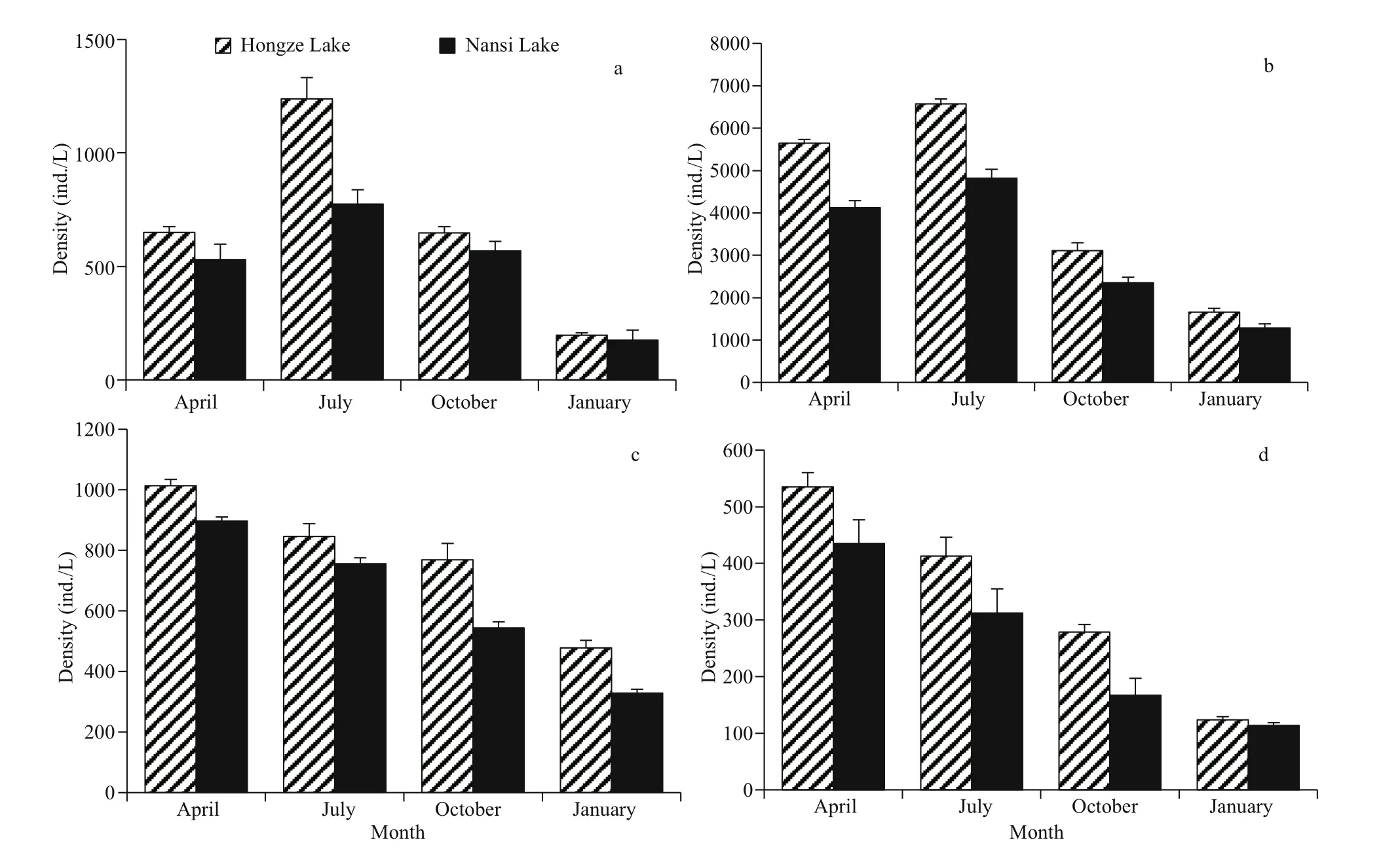
Fig.2 Comparison of temporal changes of zooplankton density in Nansi Lake and Hongze Lake

Fig.3 Average density of dominant zooplankton species in Nansi Lake (a) and Hongze Lake (b)
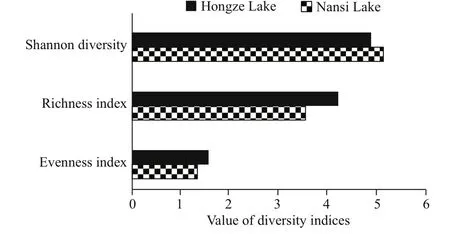
Fig.4 Zooplankton diversity indices (the richness index,evenness index, and Shannon index) in Nansi Lake and Hongze Lake
3.2 Dominant species and diversity indices
In Nansi Lake, the most dominant species were Keratella tecta, Keratella valga, and Lecane luna for rotifera (Fig.3a). In Hongze lake, the most dominant species were limnetic, e.g., Brachionus angularis,Brachionus forficula, and Polyarthra vulgaris for rotifer (Fig.3b). Shannon-Wiener diversity index (5.13)was higher in Nansi Lake than in Hongze Lake (4.87),while the species richness index was higher (4.21) in Hongze Lake than in Nansi Lake (3.54) (Fig.4).
3.3 Factors related to seasonal dynamics of zooplankton assemblages
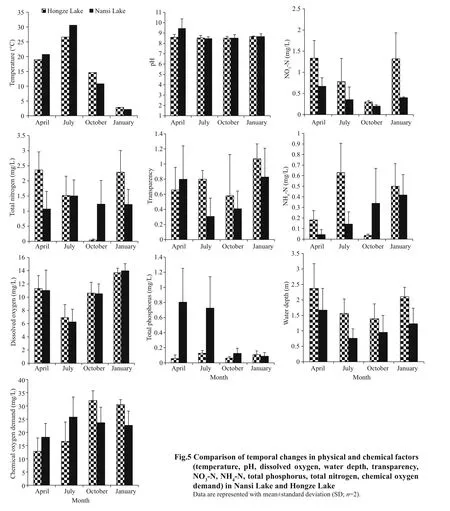
Figure 5 shows seasonal variation of physicochemical factors between Nansi Lake and Hongze Lake. In Nansi Lake, CCA identified that the environmental variables pH, total phosphorus and temperature contributed significantly to zooplankton assemblages ( P <0.05) (Fig.6a, Table 2). CCA axis 1(λ: 0.08) and CCA axis 2 (λ: 0.05) explained 63.3%and 36.7% of the total variance of the species environmental relationship respectively. The correlation effi ciencies of the first two axes were 0.72 and 0.57, respectively. The first axis was highly correlated with temperature and pH; the second axis was highly correlated to total phosphorus. The dominant species such as Ceriodaphnia cornuta, and K. tecta were associated with higher temperature values, species such as B. forficula were associated with high pH, and species such as Arcella vulgaris,L. luna, and K. valga were associated with low pH. In addition, the dominant species Moina macrocopa and Diaphanosoma leuchtenbergianum were correlated with total phosphorus.
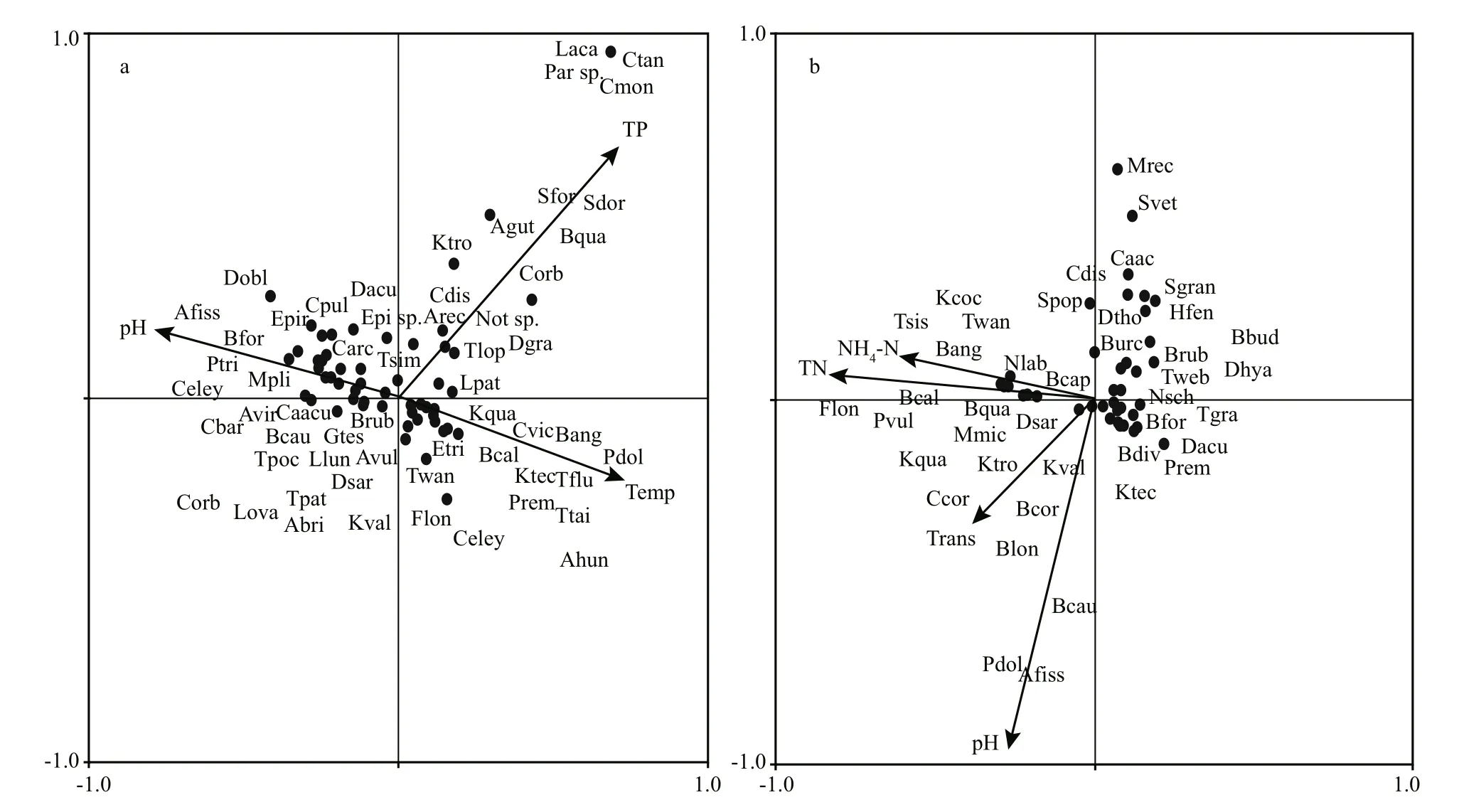
Fig.6 Canonical correspondence analysis biplots showing the relationships between zooplankton species and environmentalvariables in Nansi Lake (a) and Hongze Lake (b)
For Hongze Lake, CCA identified the environmental variables NH4- N, total nitrogen, transparency and pH as contributing significantly to zooplankton assemblages ( P <0.05). CCA axis 1 (λ: 0.08) and CCA axis 2 (λ: 0.06) explained 58.2% and 41.8% of the total variance of the species environmental relationship respectively. The correlation effi ciencies of the first two axes were 0.79 and 0.68, respectively. The first axis was highly correlated with NH4-N, total nitrogen and transparency; the second axis was highly correlated with pH. The dominant species such as B. angularis,P. vulgaris, Tintinnopsis sinensis, Tintinnopsis wangi,and Moina micrura were associated with higher levels of NH4-N and total nitrogen, whereas species such K. tropica, C. cornuta, Bosmina coregoni, and B. longirostris were associated with high transparency.In addition, the species Anuraeopsis fissa andPolyarthra dolichoptera were more strongly correlated with pH. However, in the ordination plot of environmental variables and community structure,few species were scattered near the origin, which represents average values for environmental variables for these species (Fig.6b; Table 2).
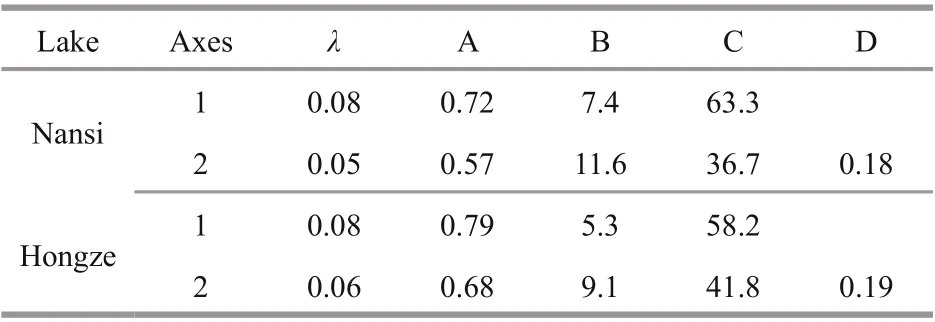
Table 2 Summary statistics for the first two axes of CCA performed between environmental variables and zooplankton species for the Nansi Lake and Hongze Lake
4 DISCUSSION
Water levels in Nansi and Hongze Lakes were strongly influenced by the operation of the East Route of the South-to-North Water Transfer project. The ESNT likely led to lower transparency, increased nutrients and more fluctuations in water level, which could have significantly altered abiotic and biotic factors in the lakes. More than 50% of zooplankton species (either limnetic or littoral) appeared within the category of sporadic in lakes. Water level fluctuations due to water exchange aff ected the zooplankton assemblage by the appearance or disappearance of certain taxa. Opportunistic peaks and declines of certain zooplankton taxa were likely correlated with the changing lake habitats and food resources aff ected by water transfer. Therefore, water level fluctuations may influence extreme habitat alterations in these ecosystems (Balkić et al., 2018).
The comparative analysis of zooplankton community assemblages showed that a large degree of variation occurred concerning species composition,habitat preference, and density of zooplankton in Nansi Lake and Hongze Lake. Protozoans and Rotifera species diversity and abundance were a characteristic feature of the two lakes. The current study found 133 zooplankton species in Nansi Lake and 122 in Hongze Lake. When compared to previous studies, there were 249 species reported in the early 1980’s in Nansi Lake, and then species richness drastically declined to 28 species in 2002 and 48 species in 2008 (Gong et al., 2010). The zooplankton richness was gradually recovered with 163 species in 2012 (Chen et al., 2016). Zooplankton species richness was again reported to have decreased during surveys from 2011-2015. A total of 58 species of zooplankton were observed in 2011, 62 in 2012, 67 in 2013-2014 and 65 in 2015 (Meng et al., 2017). The data of the present study have shown a substantial decrease in the abundance of zooplankton communities in Nansi Lake. The contribution of pelagic species decreased, and the proportion oflittoral species of zooplanktons increased. The decreased diversity of pelagic species is probably a consequence of the leaching action of flowing water(Havel et al., 2000). A key factor influencing the occurrence of particular species in the littoral zone and, in consequence, the diversity of communities colonizing this zone, is the availability of diverse habitats (Balayla and Moss, 2003). Yang (2003)reported 91 species of zooplankton and an average abundance of 1 458 ind./L in Hongze Lake. Du et al.(2014) reported 34 rotifer species comprised with the dominant species Keratella cochlearis, Asplanchna priodonta, Polyarthra dolichoptera, Brachionus calyciflorus, and Keratella valga were collected in 2011. Among the total zooplankton species, most of the species were limnetic, and remaining taxa were either littoral or common in both habitats. Therefore,the qualitative structure of zooplankton of Hongze Lake was significantly comprised oflimnetic species.However, an increase in the percentage of pelagic rotifers in the total density of zooplankton was also observed in Hongze Lake. The greatest increase was noted in the limnetic species, B. angularis, B. forficula,P. vulgaris, and K. cochlearis, although these taxa are ubiquitous in many waters and are most common in flowing ecosystem (Ejsmont-Karabin and Kruk,1998; Kobayashi et al., 1998; Czerniawski and Domagała, 2010; Czerniawski and Pilecka-Rapacz,2011). This phenomenon is connected with the short generation cycle of this group and the possibility to satisfy the threshold food concentration in scanty river waters (Baranyi et al., 2002). However, the largest changes in habitat preference of the species,especially limnetic ones clearly indicate the eff ect of the water transfer on the lakes. Based on the morphology and habitat types of Hongze Lake, it may be subjected to more ecological eff ects of water transfer. Following ESNT, the downstream regions of Hongze Lake could be experiencing hydrological instability from high variability of water current in the lotic water transfer system. A second factor may be aff ecting the Rotifera richness in the pelagic zone, as a result of washout of aquatic macrophytes by the currents. Another factor could be that the ESNT connection between Hongze Lake and Changjiang River and nearby lakes through canals will allow recruitment of pelagic species to the lakes. Such plankton exchange could especially influence patterns of flowing water species such as K. cochlearis.
Protozoans and rotifers were the most abundant zooplankton group in both lakes. More abundant populations of rotifer genera ( Lecane, Keratella in Nansi Lake, Brachionus and Polyarthra in Hongze Lake) and protozoans ( Diffl ugia and Arcella in Nansi Lake and Tintinnopsis in Hongze Lake) were found during summer coinciding with lower water levels.Succession of the planktonic species, especially rotifers and protozoans, are determined by the environment with plentiful food resources of algae and detritus (Auer et al., 2004; Wallace et al., 2006;Sodré-Neto and Araújo, 2008). The dominant planktonic groups had opportunistic life history traits,with short life cycles and resilience to variation in the hydrologic regime. When compared to protozoans and rotifers, copepods and cladocerans tended to have low diversity and density in the studied lakes. The more abundant cladoceran genera ( Diaphanosoma,Moina, and Ceriodaphnia in Nansi Lake; Moina in Hongze Lake) were developed during the summerautumn seasons at lower water levels. The largebodied cladoceran Daphnia was also found absent in Nansi Lake. In lower water levels, nutrient concentrations may be high, and a strong prevalence of few species preferring higher biogen concentrations,combined with a decrease in chemical components,may have caused an eradication of more sensitive taxa (Sampaio and López, 2000). In lakes, fish predation on large crustaceans may lead to a shift towards smaller crustaceans and rotifers (Brooks and Dodson, 1965; Korponai et al., 1997). The dominant copepod genera ( Diacyclops and Microcyclops in Nansi Lake; Mesocyclops and Sinocalanus in Hongze Lake) occurred during spring and summer seasons,respectively. Higher temperatures during warmer months accelerate the growth of phytoplankton productivity that exceeded the growth of certain species. Dominance of cyclops and sinocalanus might be influenced by the availability of food resources in particular seasons (Simões et al., 2013).
Zooplankton community structure and their seasonal dynamics in the lacustrine environment are driven by a variety of factors such as lake morphology,hydrological properties, climate conditions, and anthropogenic activities (Fryer, 1985). CCA analysis identified diff ering significant environment variables that explained maximum variability in zooplankton community structure in the two lakes. These results confirm numerous former studies and show that abiotic factors such as pH, nutrients (N and P),temperature, and turbidity concentrations are major driving forces that influence directly or indirectly the zooplankton assemblage (Bērzinš and Pejler, 1989;Pinel-Alloul et al., 1990; Dodson, 1992; Jeppesen et al., 1994; Derry et al., 2003). Temperature significantly explained zooplankton variation in Nansi Lake, with P. dolichoptera, Filinia terminalis, and K. tecta being associated with increasing temperature. Rotifers generally have an extensive tolerance to temperature,but in separate lakes, they are restricted with temperature differences (Bērzinš and Pejler 1989). In the present study, F. terminalis and P. dolichoptera were associated with high temperature, but these species usually are considered to be “winter species”,preferring temperatures below 10 °C (Carlin, 1943;Galkovskaya et al., 2006). However, other former studies also observed the occurrence of these species at higher temperatures in small lakes and ponds(Bērzinš and Pejler, 1989). Higher temperatures can theoretically promote comparable benefits for small zooplankton species exhibiting strategy life history traits and growth patterns (May, 1983; Hessen et al.,1995; Bunioto and Arcifa, 2007). B. calyciflorus was found to favor low temperatures in this study, but not in other studies (Bērzinš and Pejler, 1989; Xiang et al., 2010; Ji et al., 2013), K. quadrata preferred low temperatures in our study as well as in other studies(May, 1983; Galkovskaya et al., 2006; Wen et al.,2011). These diff ering results could be explained in that temperature alone does not generally determine when and where a species occurs, as there is a combination of other biotic and abiotic factors that contribute along with temperature (Bērzinš and Pejler,1989). In addition, Daphnia hyalina, and Cyclops vicinus peaked at low temperatures, while other species such as Thermocyclops taihokuensis and Ceriodaphnia cornuta peaked at relatively high temperatures. Early spring species Cyclops vicinus and Daphnia hyalina normally prefer temperatures below 25 °C, attaining a higher growth rate and development at this temperature (Chiang and Du,1979; Maier, 1989). Seasonal changes in temperature likely attributed to the changes in species composition and density of species (Tackx et al., 2004). Hongze Lake dominant species C. cornuta, Bosmina coregoni and Bosmina longirostris and Keratella tropica were associated with lower transparency, which may provide better shelter for this zooplankton than in clear water (Dodson, 1990; Wissel and Ramacharan,2003). In Hongze Lake, dominant species T. wangi,T. sinensis, K. cochlearis, B. angularis, and P. vulgaris were correlated to NH4-N and total nitrogen, while in Nansi Lake Diffl ugia gramen, Centropyxis orbicularis, B. calyciflorus, and Keratella tropica were correlated to total phosphorus. Nutrients including NH4-N, NO3-N, total nitrogen, and total phosphorus can indirectly aff ect protozoans and rotifers via trophic cascades. Rotifers are unselective micro filtrates, feeding on particles in the range of 0.5 to 20 μm with a diff erential ability to feed on bacteria, protozoans, heterotrophic flagellates, and numerous algae, including pico- and nanophytoplankton (Arndt, 1993). In Nansi Lake,dominant species such as Moina macrocopa,Diaphanosoma leuchtenbergianum, and the copepods Sinocalanus doerri and Schmackeria forbesi were associated with total phosphorus concentration. Total phosphorus influences the growth of crustaceans by reflecting edible food materials for them. Cladocerans share available food with rotifers, and the majority of cladocerans are filter feeders (Gilbert, 1966; Yan,1986; Pinto-Coelho et al., 2005).
5 CONCLUSION
Our study demonstrated that the differences in the zooplankton assemblages are likely due to water level fluctuations caused by water transfer, and some environmental conditions are likely responsible for biotic changes and ecosystem functioning in the studied freshwater lakes. Water exchange significantly aff ected the lake morphology due to water level fluctuations being responsible for the habitat preference of the zooplankton community structure in studied lakes. The stated decrease in taxonomic diversity of crustacean plankton fauna and the prevalence of specific zooplankton taxa, likely correlated with the change oflake habitats and food resources in the studied lakes. Further studies are needed to predict the influence of the ESNT on the hydrological and biotic community (phytoplankton,zooplankton, and fishes) in Nansi and Hongze Lakes.
6 DATA AVAILABILITY STATEMENT
All data generated or analyses during this study are included in this published article.
7 ACKNOWLEDGMENT
We thank the Institute of Hydrobiology for hosting this research work. We would also like to thank SONG Yiqing for supporting statistical analysis on the manuscript.
8 DISCLOSURE STATEMENT
There are no conflicts ofinterest arisen by authors.
 Journal of Oceanology and Limnology2021年2期
Journal of Oceanology and Limnology2021年2期
- Journal of Oceanology and Limnology的其它文章
- Predicting sediment flux from continental shelfislands,southeastern China*
- Laboratory simulation of dissolved oxygen reduction and ammonia nitrogen generation in the decay stage of harmful algae bloom*
- Development of high-resolution chloroplast markers for intraspecific phylogeographic studies of Phaeocystis globosa*
- Effects ofiron and humic acid on competition between Microcystis aeruginosa and Scenedesmus obliquus revealed by HPLC analysis of pigments*
- Effect of river plume on phytoplankton community structure in Zhujiang River estuary*
- Exploring the sublethal genotoxic effects of class II organophosphorus insecticide quinalphos on freshwater fish Cyprinus carpio