
沙质海岸基干林木麻黄幼林模拟抚育预测
2021-04-08 04:16:56尤龙辉叶功富程分生余锦林
福建农林大学学报(自然科学版) 2021年2期
杨 洋, 尤龙辉, 叶功富, 聂 森, 程分生, 余锦林
(1.福建农林大学林学院,福建 福州 350002;2.福建省林业科学研究院,福建 福州 350012;3.福州市林业局自然保护地规划发展中心,福建 福州 350007)
光合作用、蒸腾作用和呼吸作用是林木冠层的三大功能,而冠幅又是描述林木冠层的重要指标之一[1],传统的森林经营培育,常利用冠幅等因子反映林分立木竞争及树木活力[2].随着计算机模拟技术在林业生产经营中的应用,对森林进行可视化模拟经营的理论和方法也日趋深化和成熟[3],冠幅是森林经营效果评价、生产潜力预估及养分循环等研究中重要的可视化因子[4-7].许多研究者认为,林木冠幅与胸径关系密切,而且通过监测林分冠幅,可以进一步估算林分胸高断面积、林分蓄积量等重要生产指标[8-10].因此,利用模型描述复杂的林分生长规律,实现森林动态变化模拟,已成为林业研究的一个重要方向[11].例如王成德[12]利用树冠外轮廓模型对桉树和杉木人工林进行三维可视化模拟,构建了一个能够服务于桉树和杉木人工林小班经营模拟的辅助决策系统;梅光义[13]对杉木人工林建立冠高、冠幅、胸径等系列生长模型,利用Forestsimulator系统模拟不同经营措施对杉木林分的蓄积量、生物量和景观质量的影响.目前,描述林木冠幅与胸径的关系模型包括线性和非线性模型两大类,研究表明[14-15],在一定生长阶段内,林木冠幅与胸径线性关系显著;而董晨等[2]、符利勇等[16]则指出,利用非线性理论模型描述林木冠幅与胸径间的生长规律更符合生物学现实意义.
木麻黄(Casuarinaequisetifolia)是一种分布于我国东南沿海和台湾地区重要的海岸带防护林树种,具有耐盐碱、抗风沙,生长迅速等优良特点.自20世纪50年代以来,在我国东南沿海地区大面积推广种植,具有良好的防风沙、抗台风和风暴潮等重要作用,成为海岸区域经济生产建设和安全的重要生态屏障[17].但是一直以来,海岸木麻黄防护林的抚育管理较粗放,例如前沿基干林带为了提高造林存活率,常采用密植的方式进行造林[18],成林后一般不进一步抚育管理,但随着木麻黄的生长,树种间开始产生相互竞争互抑互害的现象,若未能及时把握抚育间伐时机,将不利于树木的径向生长,致使树干纤细,抗风能力弱,受极端强风天气如台风的影响,容易造成毁灭性灾害[19].罗冠勇等[20]、陈绶柱等[21]研究也表明,立木胸径与抗风性能的相关性最高,立木胸径越大,抗风能力越强.
精准调控林分密度,是提高木麻黄基干林林分生产力及其防风效能的重要手段.前期密植的基干林虽然能提高造林成活率,但成林后随着林木的生长,单木相互间竞争加剧,林木冠幅和胸径生长受到严重抑制,且林分横断面形状由于高林分密度形成的紧密结构,遇到强风天气时,一方面纤细的树干容易发生大面积折干现象,严重损毁基干林带;另一方面不透风的基干林带会迫使强风越过基干林冠层,在林带背面形成强风湍流,危害后沿农田[22].适当抚育间伐不但能增加基干林透风系数,发挥林带纵深优势,化整为零、以疏代堵,使进入林带的强风以树叶摩擦、树干摆动等形式逐渐消耗[23],而且能促进林木径向生长,提升其抗御风沙危害的能力[24].因此,利用计算机技术精准模拟抚育预测不同林分株数密度幼林抚育间伐年限及其胸径、冠幅生长量,可为评估木麻黄基干林防风效能及抗御台风灾害天气能力提供理论依据.
鉴于此,本研究以福建省平潭岛幸福洋6 a生木麻黄基干林为研究对象,利用6种典型的冠幅—胸径关系模型对林分进行拟合,筛选最优模型,再根据前人总结的木麻黄胸径生长模型,采用Matlab GUI设计程序,对不同林分株数密度基干林的冠幅—胸径生长动态进行模拟抚育间伐,并分别预测其最有利于木麻黄单木径向生长的再次郁闭年限,为提升木麻黄基干林带应对极端强风天气能力及实现木麻黄防护林精准抚育管理提供参考.
1 研究区与研究方法
1.1 研究区概况
福建省平潭岛(25°15′—25°45′N,119°32′—120°10′E)属南亚热带半湿润海洋性季风气候,为福建省少雨区之一,常年平均年降水量约1 200 mm.春雨季(2—4月),占年降水量的26%;梅雨季(5—6月),占36%;台风雨季(7—9月),占26%;少雨季(10月至次年1月),占12%.夏季和冬季以偏南风为主,年均最大风速6.9 m·s-1,出现≥8级风力的天数为84.5 d.每年受台风的袭击与影响多达5~7次.海岸带土壤以滨海性潮风沙土为主,包括红壤性风积沙土、泥炭性风积沙土和潮积沙土,滨海沙土水肥含量低,其中全氮含量1.68 g·kg-1,水解性氮含量14.61 g·kg-1,速效钾含量23.74 g·kg-1,有效磷含量35.31 mg·kg-1,有机质含量2.14 g·kg-1,土壤体积含水率6.2%.岛内常见植被种类主要以木麻黄(Casuarinaequisetifolia)、黑松(Pinusthunbergii)、湿地松(Pinuselliottii)和台湾相思(Acaciaconfusa)为主(图1).
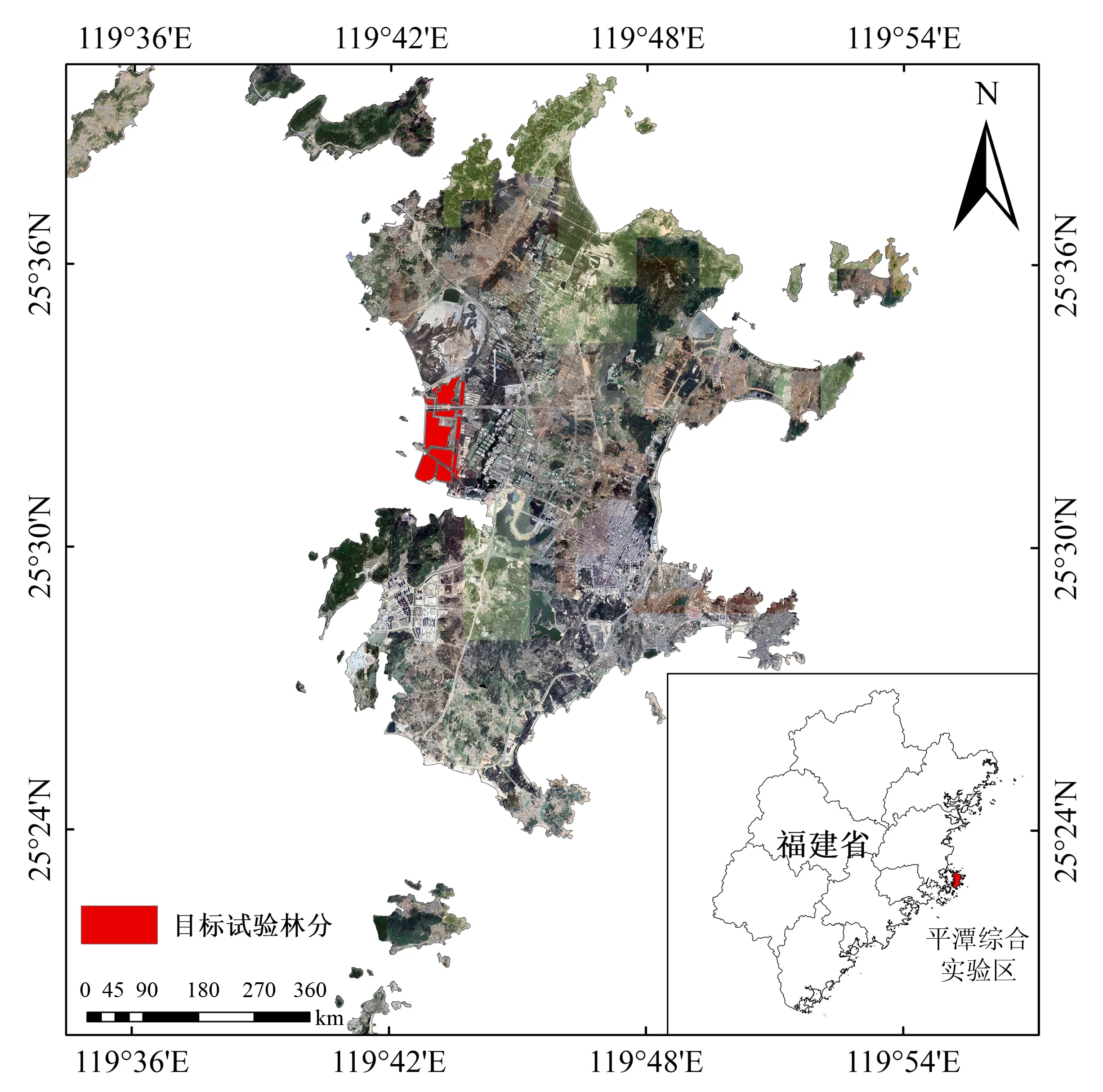
图1 研究区及样地位置Fig.1 Study area and sample plot location
1.2 样地调查与数据整理
2019年5月,在平潭岛幸福洋依次对林分株数密度>3 000株·hm-2、2 000~3 000株·hm-2和<2 000株·hm-2(3种林分密度类型为沿海木麻黄基干林常用的造林密度[25-26])的6 a生木麻黄基干林,分别布设20 m×30 m的标准调查样地各5块,共计15块,总调查面积9 000 m2.试验林标准调查样地的确定和选择满足以下条件:(1)覆盖该地区不同的林分立地条件;(2)标准调查样地之间的距离均大于50 m;(3)林相相对整齐且远离林分边缘地带.然后对各样地进行每木检尺(胸径>1 cm起测),记录树高、胸径、冠幅(包括东西和南北冠幅)、活枝下高、树木健康状况(是否有病虫害)及样地每木相对坐标,并绘制于坐标纸上,本次共计调查木麻黄样木2 135株,样地林分统计信息详见表1.
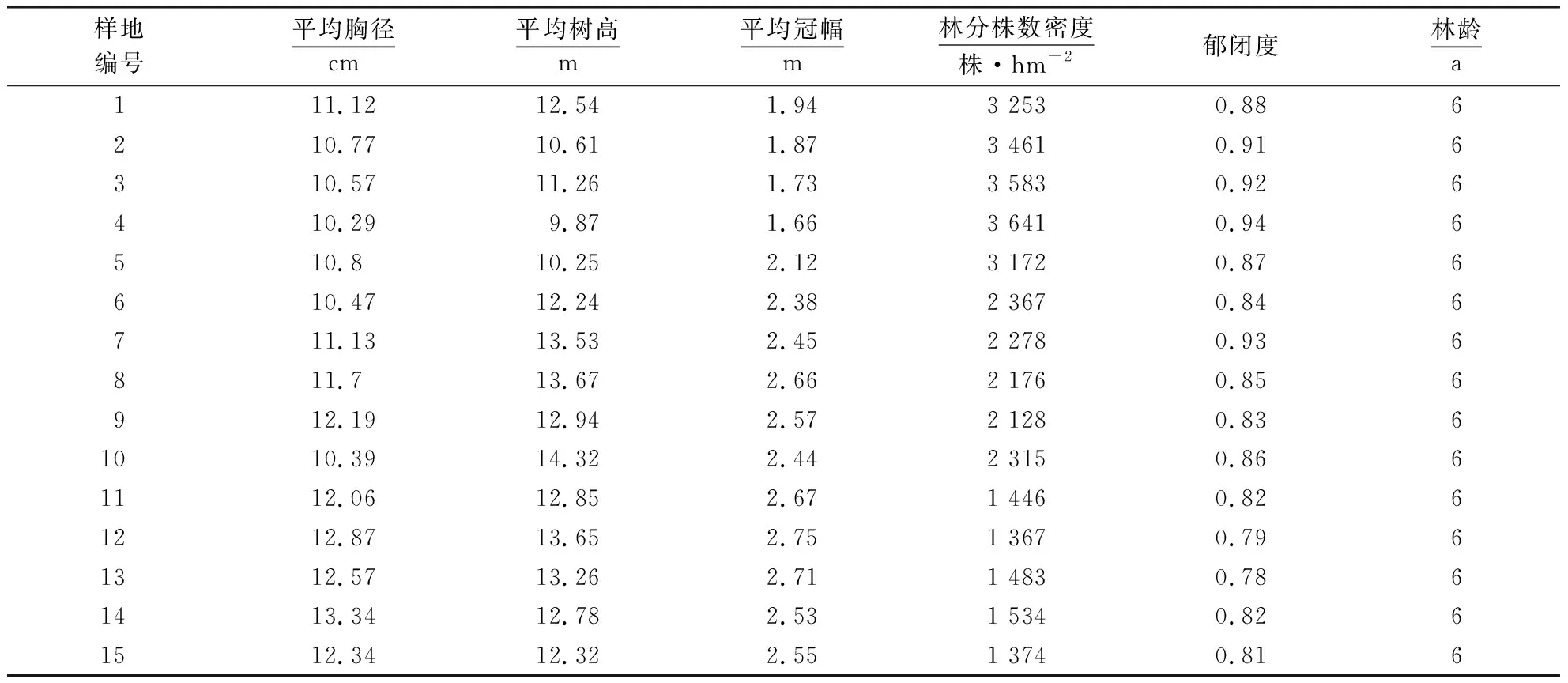
表1 木麻黄模拟抚育目标林分样地基本情况Table 1 Basic situation of sample plot of C.equisetifolia plantation simulated tending target forest
将调查数据依据8∶2的原则分成相互独立的两部分[27],即建模数据与检验数据,用以拟合和检验试验林冠幅—胸径关系模型.其中,用于模型拟合的样木共计1 685株,而用于模型检验的样木共计450株(表2).
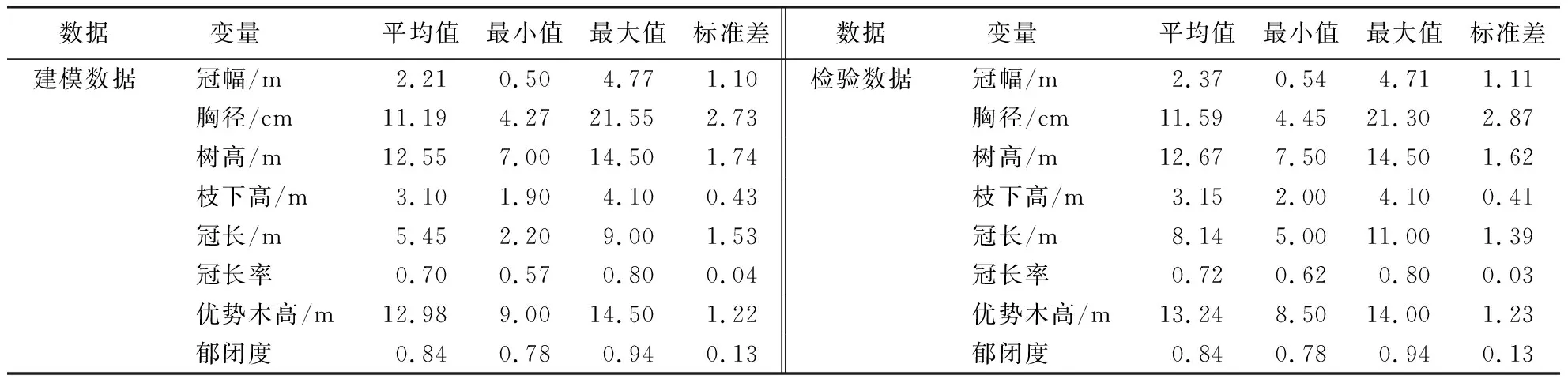
表2 木麻黄模拟抚育目标林分基本统计特征Table 2 Basic statistical characteristics of simulated tending target stand of C.equisetifolia
1.3 木麻黄胸径生长数学模型
采用叶功富等[28]对福建东南沿海强风区不均性风积沙土的木麻黄解析木建立的胸径生长模型,作为计算机模拟抚育程序设计的基础模型之一,其表达式如下:
(1)
其中,DBH(diameter at breast height)为木麻黄胸径,t为树龄.
1.4 冠幅—胸径关系模型的拟合与检验
选用6种典型的冠幅—胸径关系模型[29-31],根据平均绝对误差(MAE)和剩余均方根误差(RMSE)、决定系数(R2)和变异系数(CV)等检验指标,对各模型计算结果的可靠性和准确性进行评价和检验,选择精度最高的模型作为计算机实现可视化模拟预测的基础模型.其中,MAE、RMSE和CV的值越小,R2的值越大,说明基础模型的计算精度越高.模型表达式见表3.
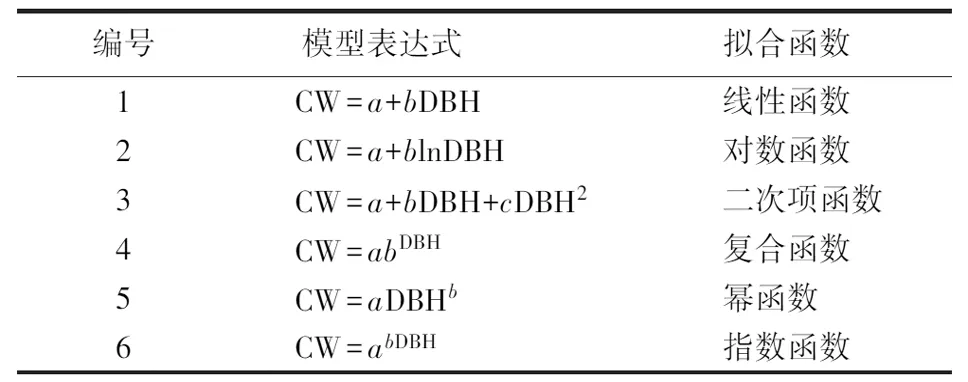
表3 冠幅—胸径模型表达式Table 3 Expression of crown width-DBH model
其中,CW(crown width)为木麻黄冠幅,DBH(diameter at breast height)为木麻黄胸径.
(2)
(3)
(4)
(5)
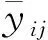
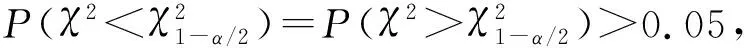
1.5 模拟抚育与预测
调查样地每木冠幅(因南北冠径和东西冠径较接近,取其平均值)及坐标位置,利用Matlab R2017a GUI绘制树冠垂直投影图,再将木麻黄胸径生长模型和筛选的冠幅—胸径关系模型通过GUI回调函数进行程序设计,通过控制郁闭度,构建模拟抚育预测系统(以下简称“系统”).模拟抚育时,系统根据输入的林分调查信息(包括树高、胸径、冠幅、坐标及健康状况等),以《森林抚育规程》为标准,优先伐除样地内枯病腐木以及树冠重叠率较高[33]且树高、胸径较小的被压木.
研究表明,在木麻黄干材积累阶段,郁闭度为0.6~0.8时,最有利于林分单木胸径的生长[34-36],而基干林郁闭度低于0.6时,会降低木麻黄的保存率[29].因此,目标试验林模拟抚育后郁闭度以0.6~0.7为宜,并模拟预测其郁闭度达到0.8~0.9时的再次郁闭年限及林分平均胸径和冠幅增长率.
1.6 数据处理
采用Microsoft Excel 2016整理数据,IBM SPSS25.0模型拟合分析与检验,Origin9.5作图,Matlab R2017a GUI编程.
2 结果与分析
2.1 直径分布
目标试验林胸径分布在4.1~21.8cm的区间内,平均胸径为11.44 cm,标准差为4.10(图2).试验林的直径分布接近正态分布,峰值在10~12径阶.总体而言,数据符合建模要求.

图2 胸径分布直方图Fig.2 Histogram of DBH distribution
2.2 冠幅—胸径模型拟合、检验与最优模型筛选
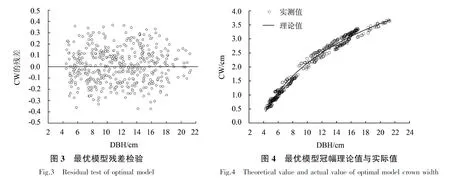
由表4可知,模型2的MAE、RMSE和CV均较小,R2最大,且较符合生物学现实意义(即幼林前期郁闭度较低,冠幅随着胸径生长快速增加,林分郁闭后,单木间开始互抑互害,冠幅生长逐渐放缓,林分进入干材累积阶段,而对数函数模型能较好地解释该生物学现象),说明该冠幅—胸径关系模型拟合效果最好,其表达式为:CW=-2.558+2.037 ln DBH.模型参数详见表5.
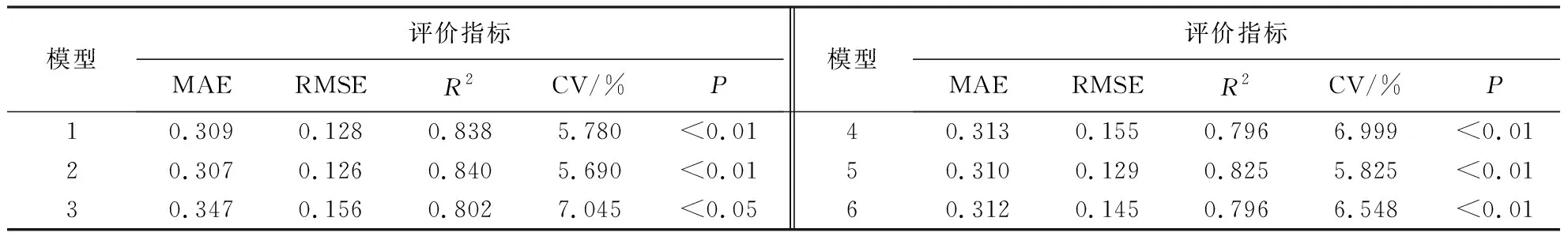
表4 模型评价指标Table 4 Model evaluation index
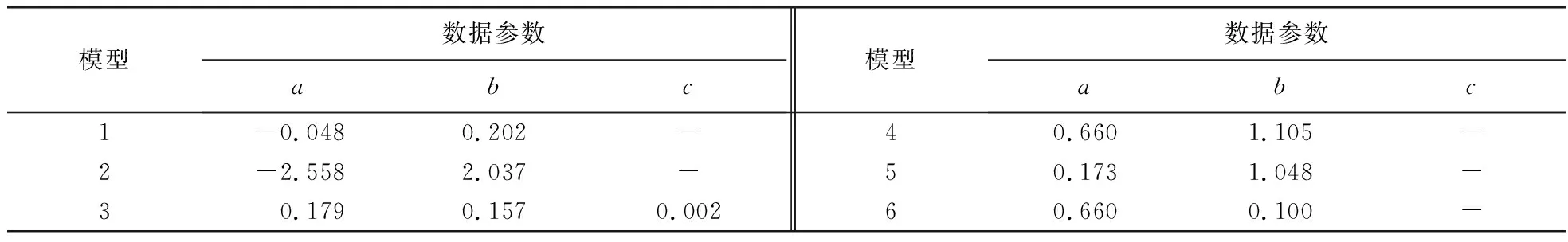
表5 模型数据参数1)Table 5 Model data parameters
2.3 最优模型的假定性检验
由图3可以看出,最优模型的残差结果在横轴上下均匀分布,且标准化残差值均处于(-0.3,0.3)区间内,包含于统计规定范围[-2,2]的区间,因此,筛选的最优模型假定成立,且模型参数估计无偏.
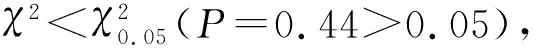
2.4 典型模拟抚育预测试验样地的选取
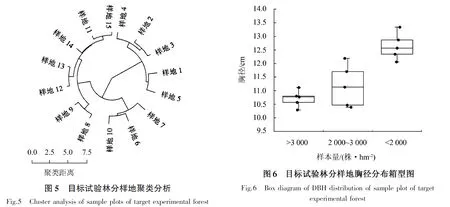
根据木麻黄林分树高、胸径、冠幅、林分株数密度和郁闭度等因子,对15个调查样地进行聚类分析(图5),样地1~5、样地6~10和样地11~15总体上可以分成3个组别.由图6可以看出,样地2、样地7和样地11的平均胸径最接近各组别胸径的平均值,因此,本文筛选样地2、样地7和样地11作为可视化模拟抚育预测的对象.
2.5 模拟抚育与预测
选取样地2(林分株数密度>3 000株·hm-2)、样地7(林分株数密度2 000~3 000株·hm-2)和样地11(林分株数密度<2 000株·hm-2)3种类型试验林分,采用设计的系统进行模拟抚育,抚育效果如图7所示.
由图7和表6可知,样地2林分株数密度较高,林木冠幅重叠率较高,胸径较小,互抑互害现象较严重,因此,需要进行强度间伐,以郁闭度0.65为标准(下同),伐除枯病腐木及林木树冠重叠率较高且树高、胸径较小的被压木.利用系统模拟抚育后,保留木株数密度降为1 916株·hm-2,胸高断面积降为14 743 cm2·hm-2,林分株数抚育强度和胸高断面积抚育强度分别为44.80%和27.80%;平均胸径增长了12.5%.
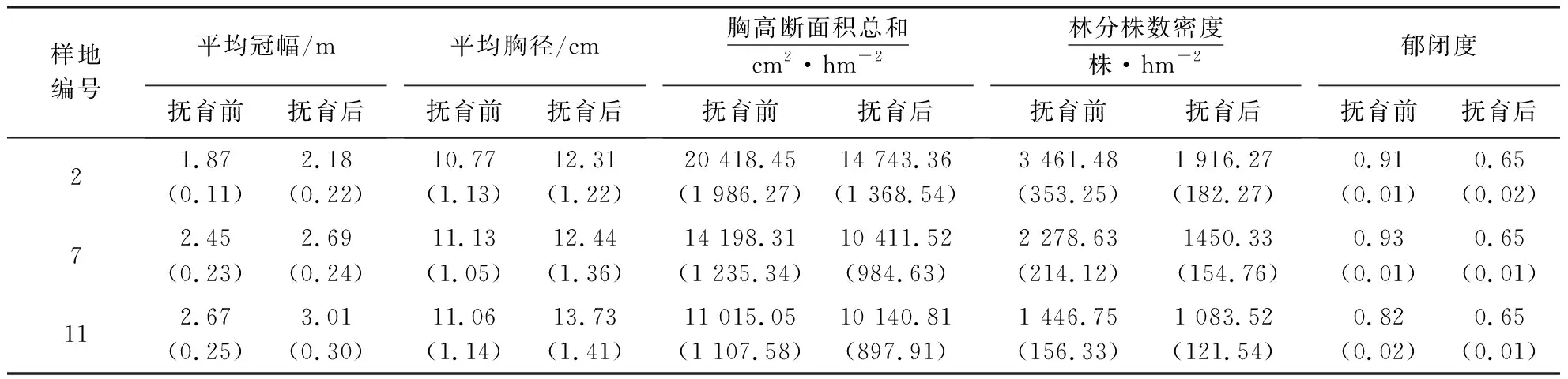
表6 不同林分株数密度木麻黄林分抚育前后冠幅胸径对比1)Table 6 Comparison of crown diameter at breast height of C.equisetifolia stands with different initial planting densities before and after tending
样地7虽然树冠重叠率较低,但郁闭度较高,为促进单木径向生长,需进行中度间伐.伐后保留木株数密度降为1 450株·hm-2,胸高断面积降为10 411.52 cm2·hm-2,林分株数抚育强度和胸高断面积抚育强度分别为36.50%和26.68%;平均胸径增长了10.5%.
样地11林木胸径结构分布较均匀,主要是进行生长伐,伐除林分中的枯病腐木,减少病虫害的传播.伐后保留木株数密度降为1 083株·hm-2,胸高断面积降为10 140.81 cm2·hm-2,林分株数抚育强度和胸高断面积抚育强度分别为25.30%和8.1%;平均胸径增长了19.5%.
如图7和图8所示,样地2在首次模拟抚育后第6年郁闭度达到了0.80,第9年达到了0.90;样地7首次模拟抚育后第4 a郁闭度达到了0.81,第6年达到了0.92;样地11首次模拟抚育后第5年郁闭度达到0.82,第7年达到了0.93.因此,林分株数密度>3 000株·hm-2的6年生木麻黄基干林可在抚育后第6年进行再次抚育,最晚不超过9年;林分株数密度2 000~3 000株·hm-2的林分可在第4年进行再次抚育,最晚不超过第6年;林分株数密度<2 000株·hm-2的林分可在第5年进行再次抚育,最晚不超过第7年.

图7 不同林分株数密度木麻黄林分抚育前后俯视图对比Fig.7 Comparison of top view of C.equisetifolia stands with different initial planting densities before and after tending

图8 不同林分株数密度木麻黄林分模拟抚育后俯视图对比Fig.8 Comparison of top views of C.equisetifolia stands with different initial planting densities after simulated tending
由表7可知,样地2抚育间伐后第6年平均冠幅增长了31.65%,平均胸径增长了16.73%;样地7抚育间伐后第4年平均冠幅增长了11.89%,平均胸径增长了24.03%;样地11在抚育间伐后第5年平均冠幅增长了13.95%,平均胸径增长了19.08%.样地2平均冠幅增长率高于样地7和样地11,而平均胸径增长率则表现为样地7较高,样地2和样地11较低.
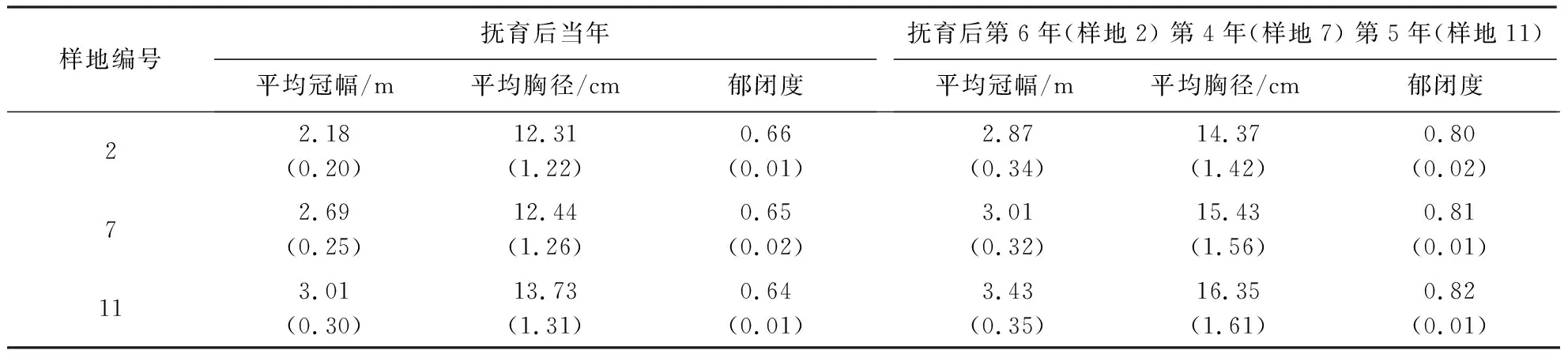
表7 不同林分株数密度木麻黄林分模拟抚育后效果1)Table 7 Effects of simulated tending on C.equisetifolia stands with different initial planting densities
2.6 初始林分株数密度与二次抚育年限的相关关系
由图9可知,目标试验林初始林分株数密度与模拟再次郁闭年限的相关关系用二次项函数模型拟合效果较好,即初次模拟抚育后,林分郁闭度再次达到0.8和0.9时总体趋势均表现为随着初始林分株树密度的增大,模拟再次郁闭年限呈现先降低后升高的趋势.
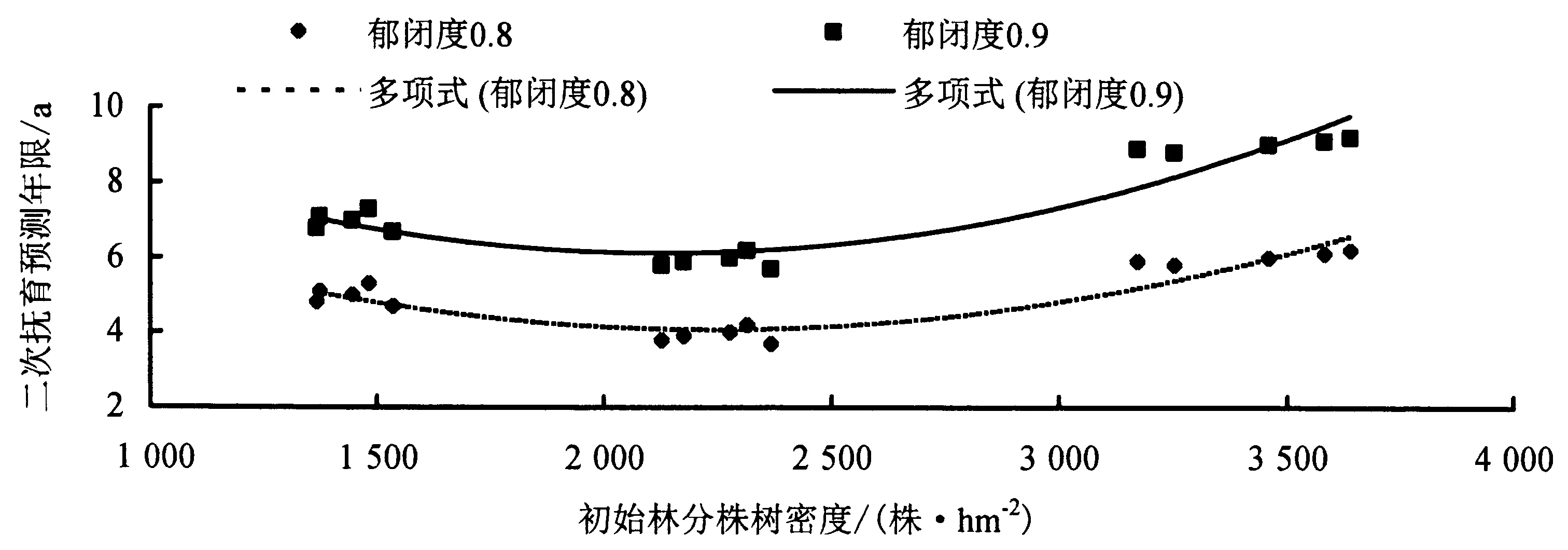
图9 木麻黄初始林分株数密度与模拟再次郁闭年限的相关关系Fig.9 Relationship between number density of initial C.equisetifolia stands and simulated re-closure years
3 讨论
本研究发现,对数函数模型能较好地表达目标试验林分冠幅—胸径间的生长关系,这与王媛等[37]对处于幼龄阶段的乔木树种构建的冠幅—胸径关系模型相似.模拟抚育显示,林分株数密度较高的样地2间伐后,林分平均冠幅增长率高于样地7和样地11,说明林分株数密度对木麻黄幼林树冠生长有显著的抑制作用,与前人对侧柏(Platycladusorientalis)[38]、山桃(Prunusdavidiana)[39]、南方红豆杉(Taxuschinensisvar.mairei)[40]、杉木(Cunninghamialanceolata)[41]等树种的研究结论一致.抚育间伐前,初植密度越高的木麻黄基干林,单木相互间生境资源竞争越激烈,生长受到抑制的程度越大.抚育间伐后,生长资源竞争矛盾得到有效缓解,抚育强度越高的林分,保留木能占据的生长空间越大,越有利于枝叶的伸展扩张[42].
平均胸径增长率表现为样地7较高,样地2和样地11较低,这与林分密度对林分胸径结构分布的影响有关[43-44]:初植密度较大的木麻黄幼林,抚育时虽伐除部分小径阶木,但保留木林分株数密度仍较大,且小径阶木仍占有较高比重,因此胸径生长速率较小;而初植密度较小的木麻黄幼林,抚育主要是伐除林分中容易传播病虫害的枯病腐木及部分小径阶木,对占比较大且生长速率相对缓慢的大径阶木影响较小;中等初植密度的木麻黄幼林,其林分胸径结构较趋近于正态分布,中径阶木占比较大,抚育间伐进一步提高了中径阶木的比重,且保留木林分株数密度较小,因此胸径增长率较高.
模拟预测结果显示,样地2、样地7和样地11的二次抚育时间随初始林分密度的增加呈现先降低后升高的趋势,这与前人的研究结果一致[34],可能与木麻黄的生物学特性和前沿基干林风因素的干扰有关:木麻黄是一种他感作用较强的植物,密植会影响木麻黄单木的正常生长[45],抚育间伐后,单木纤细,抗风害能力较低,与初植密度较小,单木较粗壮、受风害较小的林分相比,生长发育年限相对迟滞.而初植密度过低,林分透风系数过高,幼林生长发育伊始就易受严重风害,木麻黄幼嫩的顶芽和当年生小枝受沿海飞盐强风的胁迫,形成生理干旱,发育不良甚至枯死[46],也会对木麻黄林分的生长发育年限产生影响.因此,初始造林密度适中的木麻黄基干林,模拟再次郁闭年限最短.
4 结论
对平潭岛6 a生木麻黄基干林构建冠幅胸径关系模型,并采用Matlab GUI设计模拟抚育预测系统,结果表明基干林木麻黄幼林不同初始林分密度模拟再次郁闭年限与现有研究的实际抚育年限基本一致,因此,通过监测基干林木麻黄幼林林分冠幅,实现利用计算机预测抚育间伐年限,对沙质海岸基干林的可持续经营及现代林业的精准管理均具有重要的参考价值.同时,本研究也存在不足,如缺少树木生长过程的描述,林木内树枝形态变化描述不足等问题,还有待进一步深化与探索.
猜你喜欢
中学生博览(2020年23期)2020-12-10 10:56:15
临床检验杂志(电子版)(2020年1期)2020-04-03 04:00:16
花卉(2020年4期)2020-03-16 08:17:50
绿色科技(2019年13期)2019-08-31 02:44:12
防护林科技(2019年1期)2019-03-13 05:27:50
新农民(2019年9期)2019-02-19 14:19:13
天然产物研究与开发(2018年3期)2018-05-07 06:38:39
闽南风(2017年12期)2018-01-04 18:13:52
东南法学(2016年2期)2016-07-01 16:42:34
学苑创造·B版(2015年12期)2016-06-23 09:36:36
