
不同浓度镉对番茄幼苗生理生化指标的影响
2021-04-07 06:53张雯
农村科学实验 2021年3期
张 雯
( 新疆环境保护科学研究院,新疆 乌鲁木齐 830011)
番茄是新疆普遍种植的一种农作物。加工番茄属于新疆的特色农产品之一,据世界番茄酱协会统计数据显示,2013 年中国加工番茄产量达450 × 104 t,其中 85%以上产自新疆,新疆已成为中国最大的番茄生产基地和番茄酱加工基地。赵多勇等学者通过研究新疆昌吉市、和硕县、焉耆县三个番茄产地的重金属污染情况发现,以上三地的土壤重金属均有超标现象,其中镉的超标情况最为严重。
Cd 是对植物危害较大的重金属元素之一,有关植物受Cd 毒害及植物体内缓解Cd 毒害的机制已有大量报道。金属硫蛋白(metallothionein,简称MT)为一类广泛地存在于生物界的低分子量、富含半胱氨酸的金属结合蛋白,尤其对Cd,Zn,Cu,Au,Ag 等有较强的亲和力。有很多实验证明MT 在提高植物重金属抗性、解除对Cu,Zn,Pb,Ag,Hg,Cd 等重金属毒性方面具有重要作用,多数MT 在C 端和N 端有两个富含Cys 的结构域,其中的巯基既可以与重金属螯合形成无毒或低毒的络合物。
因此本文主要研究不同浓度Cd 对番茄生长的影响,以及Cd 诱导番茄幼苗体内金属硫蛋白的情况,以阐明番茄幼苗体内Cd 毒害的初步机理。
1.材料与方法
1.1 试验材料
供试植物:番茄“新引98-1 号”。植物培养液为霍格兰溶液。试验仪器:人工气候箱,紫外/可见分光光度计;原子吸收光谱仪等。
霍格兰溶液配制:大量元素:微量元素:铁元素以100:10:5 配制。
取籽粒饱满、均匀的番茄种子,用1.2% NaClO 溶液消毒20 min,均匀摆在垫有滤纸的培养皿中,加入适量去离子水,放入人工气候箱中避光发芽5 天。选取发芽情况较好,平均长度在3 cm 左右的番茄芽于三角烧瓶,每个三角烧瓶中放同等根数,加入适量营养液,在人工气候箱中继续培养5d,过程中补充消耗的营养液。人工气候培养箱条件:光照16 h,温度28 ℃,湿度:75。暗室8h,温度20 ℃,湿度70%。
试验浓度的设定:处理中,配制Cd 浓度分别为0、1、10 mg/L,霍格兰溶液培养为对照(CK);采样时间:分别在0,5,10,15,20 d 取样,进行测定,每组处理均设3 个重复。
1.2 试验方法
1.2.1 番茄发芽率及根长、芽长的测定
在每个垫有滤纸的培养皿中均匀摆放25 粒未发芽的外形良好的番茄种子,后每个培养皿中分别加入上述步骤中所配的各种处理溶液10 ml 在人工气候箱中避光7 天。取出后,数出成功发芽的番茄种子个数,记下数据,算出发芽率。同时测量根长、芽长数据,比较不同处理下,番茄种子的萌发情况。
1.2.2 生物量的测定
将取出的水稻幼苗样品用自来水和去离水分别冲洗三遍,之后将地上部(后均用茎部代替)与根部剪开。用刻度尺量取根长和茎长并进行记录后,放入培养皿中。
将培养皿放入烘箱,在110 ℃下杀青30 min 后将温度调至90 ℃,继续烘干24 h。取出水稻幼苗根部和茎部分别称取两部分干重并进行记录;
1.2.3 叶绿素含量的测定
称取0.1 g 的叶片,置于10 ml 比色管中,加入10 mL丙酮和无水乙醇等量混合成的提取液,加塞置于4℃冰箱中下进行浸提24 h 后,用分光光度计测定在663 nm 和645 nm 处的吸光度,记下数据并计算。
1.2.4 番茄体内Cd 含量的测定
将样品的地下部分(根)和地上部分(茎叶)分开,各自保存后烘干,称重。分别测定地上部分和地下部分的Cd 含量。称取烘干磨碎后的样品0.01g ~0.08g 置于消化罐内,加入4 mL 硝酸和2 mL 双氧水,静置5 min,加盖后消化13 min。将消化液置于锥形瓶中赶酸,冷却后转移至10ml 容量瓶中,加入1%HNO3定容,静置30 min 同时做空白试验。消解后用原子吸收光谱法测出地上部分和地下部分的Cd 含量。
1.2.5 番茄体内金属硫蛋白(MT)含量的测定
金属硫蛋白(MT)含量的测定:将曝毒后的番茄从烧杯中取出,用剪刀将样品的地下部分(根)和地上部分(茎)分开,用吸水纸小心将整组番茄根、茎周围的溶液吸尽,用电子天平称取番茄根、茎鲜样组织l g,并置于研钵中,加入0.01 mol/L,pH8.6 的Tris-HCI 缓冲液10 mL,研磨后于4℃冰箱过夜抽提。抽提液于4℃,1×104 r/min 离心30min,收集上清液。上清于100℃水浴加热2-3 min,再于4℃下,1 ×104 r/min 离心20min。收集上清液,加入3倍体积-20℃预冷的无水乙醇,-20℃过夜沉淀后,4℃下,1.2×104 r/min 离心20 min。取沉淀加入0.01mol/L Tris-HCl 缓冲液5mL,溶解数小时后,在4℃下1.5 X 104 r/min离心20min,收集上清即为MT 提取液。取提取液1mL 用Ag 饱和法测定MT 含量。
2.结果与分析
2.1 对番茄种子萌发的影响
比较番茄的根长和芽长可知,镉浓度为10mg/L 较镉浓度为1mg/L 对番茄种子发芽的抑制作用更为明显,相较于对照组CK,低浓度Cd 对根长和芽长的抑制率分别为24.09%和17.73%,而高浓度Cd 对根长和芽长的抑制率分别为36.65%和28.02。比较对照组,在同一镉浓度和硫浓度下的根长芽长可发现镉对番茄幼苗各部分生长状况抑制能力大小不同,镉对根长的抑制作用明显大于对芽长的抑制。
2.2 对番茄幼苗生物量的影响

表3 番茄幼苗茎干重随时间的变化
表2 和3 反映了水稻幼苗根干重、茎干重在不同浓度Cd 下随时间的变化情况。由表中可知,番茄幼苗根干重和茎干重均随着时间的增加而增加。相对于对照组,在番茄幼苗暴露在不同浓度Cd 的情况下,均受到不同情况的抑制,并且由表中可知,高浓度Cd 对番茄幼苗根、茎干重的抑制更明显。

表4 番茄幼苗根长随时间的变化

表5 番茄幼苗茎长随时间的变化
表4 和5 反映了水稻幼苗根长、茎长在不同浓度Cd下随时间的变化情况。
由图可知,番茄幼苗根、茎长度随生长时间的增加而逐渐增长,同时随着Cd 浓度的增加而减少,说明了高浓度Cd 对水稻幼苗的毒性更大。
番茄幼苗茎干重的变化趋势与根干重变化趋势较类似,但由于茎干重数值与对照组相比,差别并不显著所以推测,Cd 对根部的毒性大于Cd 对茎部的毒性,主要是因为番茄由根部吸收Cd,而向茎部转移量较少,因此Cd 对番茄幼苗茎的毒性低于对根的毒性。由以上4 张表格可以看出,高浓度Cd 较低浓度Cd 对番茄的毒害更加明显。
镉对植物的正常生长有很强的胁迫作用。当小麦和大麦受到镉胁迫后,种子萌发率以及根系的生长速度都会下降,且随着镉处理浓度的增大和时间的延长会加剧毒性效应。
2.3 对番茄叶绿素含量的影响
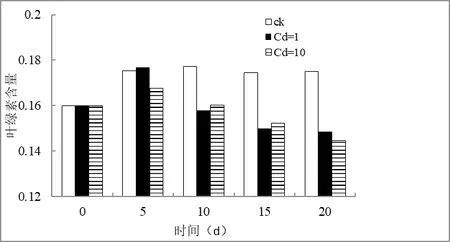
图1 番茄幼苗叶绿素含量随时间的变化
由图1 可知,随着时间增加到第5 天,不管是对照组还是添加了不同浓度的Cd,番茄体内叶绿素均有所上升,这是由于在番茄幼苗从种子萌发开始逐渐由暗室中未进行光合作用,而通过转移到培养模式后,叶绿素逐渐增多的缘故。
但随着时间的延长,番茄幼苗的叶绿素在对照组并无显著变化,而在添加不同浓度Cd 后,高浓度Cd 和低浓度Cd 均对番茄幼苗叶绿素具有明显的抑制作用。对比高浓度,在低浓度镉情况下,低浓度镉对番茄幼苗叶绿素的抑制相对高浓度不明显。
有研究表明镉可能破坏植物体内合成叶绿素所需要的酶而降低叶绿素的含量,或干扰了叶绿体中的电子载体配对。
2.4 对番茄体内镉含量的影响

图2 番茄幼苗茎中Cd含量随时间的变化

图3 番茄幼苗根中Cd含量随时间的变化
由图2、图3 可以看出,对照组由于未添加Cd,因此番茄幼苗根和茎均未发现Cd 累积。
而在经过Cd 暴露后,番茄幼苗茎部和根部镉累积随时间均有所增加,并且Cd 在根部的累积量以及累积速率均高于茎部,高浓度镉在番茄体内的含量明显高于低浓度镉在番茄体内的累积。
2.5 对番茄体内金属硫蛋白含量的影响

图4 番茄幼苗MT含量随时间的变化(低浓度镉)

图5 番茄幼苗MT含量随时间的变化(高浓度镉)
对比图4 和图5,对照组中的金属硫蛋白极低,而并且随着时间的增加也没有出现显著的变化由此推断只有在Cd 存在时,金属硫蛋白才可能被诱导。
而高浓度Cd 所诱导的金属硫蛋白要高于低浓度Cd诱导产生的金属硫蛋白,这是由于金属硫蛋白通过与重金属结合可以有效的减轻重金属对机体的毒害,重金属浓度越高,诱导产生的金属硫蛋白越多。从而络合Cd,达到缓解Cd 毒害的作用。
3.结论
添加一定浓度的Cd 的对番茄幼苗的生长有一定的毒害作用,高浓度Cd 相较于低浓度Cd 对番茄幼苗的危害程度更高。此外,从Cd 在番茄幼苗体内的累积情况来看,低浓度Cd 在番茄幼苗体内的含量比高浓度Cd 在番茄幼苗体内低,高浓度Cd 诱导产生的金属硫蛋白更高。
猜你喜欢
科学导报(2022年32期)2022-06-09
有色金属(矿山部分)(2021年4期)2021-08-30
阅读(科学探秘)(2020年8期)2020-11-06
绿色科技(2019年2期)2019-05-21
绿色科技(2017年8期)2017-05-22
中国科技纵横(2017年7期)2017-05-16
女性天地(2016年10期)2017-04-25
绿色科技(2017年2期)2017-03-23
江苏农业科学(2017年1期)2017-02-27
中国高新技术企业(2014年7期)2014-06-06
