
基于LIN28A/NF-κB信号通路研究狗肝菜多糖缓解肝纤维化的作用机制
2021-04-07 03:27郑董璇张可锋钟明利赵唐莲曹后康段小群
天然产物研究与开发 2021年3期
郑董璇,张可锋,晋 玲,钟明利,赵唐莲,曹后康,段小群*,高 雅*
1桂林医学院药学院,桂林 541004;2甘肃中医药大学药学院,兰州 730000
肝纤维化(hepatic fibrosis,HF)是由各种有害刺激,包括病毒感染、药物毒性、酒精和自身免疫性肝病的持续伤口愈合反应对肝脏损伤所致[1]。它是慢性肝病的重要病理标志,是肝硬化发生的关键事件,也是终末期肝细胞癌和晚期肝硬化的主要病因[2,3]。秋水仙素作为肝纤维化和肝硬化的临床用药,主要抑制肝星状细胞早期的活化与增殖,促进肝星状细胞凋亡,干扰胶原代谢,针对不同病因导致的HF秋水仙素治疗效果不一,并且秋水仙素会引起许多副作用,例如腹痛、恶心、食欲不振等,高剂量服用后还会导致肝肾功能衰竭[4],因此仍需研究并开发新的安全有效的治疗药物。狗肝菜(Diclipterachinensis(L.) Juss)作为中国传统中药材,味甘性凉,具有清热,凉血,利尿,解毒平肝等功效,对CCl4、D-氨基半乳糖盐酸和脂多糖等多种类型的肝损伤模型具有缓解作用[5];本课题组在先前研究中发现,狗肝菜多糖(DCP)对肝脏脂质代谢紊乱尚有良好的抑制作用,其机理主要与缓解炎症反应和抑制氧化应激有关[6]。LIN28A是一种高度保守的RNA结合蛋白,在诱导多能干细胞、调节发育、细胞生长和代谢等方面发挥着重要作用,在诸如食道癌[7]、卵巢癌[8]等多种癌细胞中,均发现存在LIN28A的过表达现象。有研究表明,肝脏特异性缺失LIN28A可缓解小鼠肝细胞癌并有效延长患癌小鼠寿命[9],这提示LIN28A可能成为治疗肝细胞癌或肝脏疾病的潜在靶点。但目前尚无文献报道其与HF的关联性,此文章旨在探索LIN28A与HF的关联性并研究DCP能否通过抑制LIN28A起到保护肝脏的作用。
1 材料与试剂
1.1 试验药物与试剂
狗肝菜多糖(由本课题组所在实验室提取制备[6]);分析纯CCl4(广东汕头市西陇化工厂);SDS-PAGE蛋白上样缓冲液(南通市碧云天生物技术研究所);秋水仙素(上海如吉生物科技发展有限公司,纯度>99%);PVDF膜(Bio-Rad公司,美国);LIN28A抗体、核因子-κB p65(NF-κBp65)抗体、转化生长子-β1(TGF-β1)、抗体基质金属蛋白酶-9(MMP9)抗体、α-平滑肌肌动蛋白(α-SMA)抗体、环氧化酶-2(COX-2)抗体和诱导型一氧化氮合酶(iNOS)抗体(Abcam,英国);GAPDH抗体(Abcam,英国);辣根标记的山羊抗兔二抗、辣根标记的山羊抗鼠二抗(Bioworld,美国);Super ECL Plus超敏发光液(上海雅酶生物科技有限公司);PCR检测试剂盒(EZbioscience,美国);透明质酸(HA)ELISA试剂盒(上海江莱生物科技有限公司);Ⅳ型胶原蛋白(Ⅳ-C)、Ⅲ型前胶原(PC Ⅲ)ELISA试剂盒(上海赛默生物科技发展有限公司);层粘连蛋白(LN)ELISA试剂盒(武汉伊莱瑞特生物科技有限公司);RIPA裂解液(上海碧云天生物技术有限公司);PMSF(北京索莱宝科技有限公司)、BCA蛋白浓度测定试剂盒(上海碧云天生物技术有限公司)。
1.2 动物
60只SPF级SD大鼠,雄性,体重180~220 g,购买于湖南斯莱克景达实验动物有限公司,许可证号SCXK(湘)2016-0002。
1.3 仪器
ZD-9550台式振荡器(海门市其林贝尔仪器制造有限公司);Centrifuge 5424R台式高速冷冻离心机(Eppendorf公司,德国);O1ympus BX51显微镜(奥林巴斯公司,日本);SpectraMax i3X酶标仪(美谷分子仪器(上海)有限公司);电泳基础电源(Bio-Rad公司,美国);7500 RT-PCR system(ThermoFisher公司,美国);CLINX化学发光成像系统(上海勤翔科学仪器有限公司)。
2 方法
2.1 造模与给药
60只雄性SD大鼠适应性喂食1周后,随机分成6组,即正常组、模型组、秋水仙素(0.12 mg/kg)组、DCP低剂量(DCP low dose,DCP-L;50 mg/kg)组、DCP中剂量(DCP medium dose,DCP-M;100 mg/kg)组和DCP高剂量(DCP high dose,DCP-H;200 mg/kg)组,每组10只。除正常组外,其余大鼠均每周腹腔注射2次40%的CCl4橄榄油溶液(1 mL/kg),建立HF大鼠模型,正常组大鼠以相同剂量腹腔注射纯橄榄油。每日根据体重,给予秋水仙素组和DCP低中高剂量组大鼠相应的药物治疗,正常组与模型组大鼠按照相同计算方式灌胃蒸馏水,持续6周。末次给药后,所有大鼠禁食不禁水16 h,乙醚麻醉后腹腔静脉取血,收集肝脏。
2.2 检测肝脏病理学
取各组大鼠相同位置1 cm正方大小的肝脏,浸泡于4%多聚甲醛组织固定液中48 h,进行包埋切片后,摊片烘片,脱蜡至水,苏木精-伊红(hematoxylin and eosin,H&E)染色和Masson染色后在O1ympus BX51光学显微镜下观察组织病理变化。
2.3 DCP对HF大鼠血清HA、LN、PC Ⅲ和Ⅳ-C的影响
大鼠血液室温静置2 h后,于4 ℃条件下,4 500 rpm离心10 min后取上层血清。采用酶联免疫吸附测定法(ELISA),严格按照检测试剂盒说明书测定血清中HA、LN、PC Ⅲ和Ⅳ-C含量。
2.4 检测肝脏炎症因子
采用酶联免疫吸附测定法(ELISA),严格按照检测试剂盒说明书(武汉伊莱瑞特生物科技有限公司)测定大鼠肝脏中IL-1β、IL-6和TNF-α含量。称取肝脏组织(80~100 mg),按质量体积比1∶9加入裂解液,冰水浴中电动匀浆机破碎,于4 ℃条件下,12 000 rpm离心15 min后取上清保存作为检测样品。严格按照试剂盒说明书进行肝脏组织中IL-1β、IL-6和TNF-α的含量测定。
2.5 测定mRNA相对含量
采用聚合酶链式反应(PCR)测定肝脏组织中α-SMA、TGF-β1、LIN28A、NF-κBp65、MMP9、COX-2和iNOS的相对含量。取各组大鼠肝脏组织约100 mg,准确加入1 mL Trizol(ThermoFisher公司,美国),冰上研磨。加入0.2 mL氯仿剧烈震荡后静置5 min,4 ℃下10 000 rpm离心15 min,取上层水相加入等体积的异丙醇,静置10 min后4 ℃下10 000 rpm离心10 min,弃上清并用预冷75%乙醇洗涤RNA沉淀,4 ℃下5 000 rpm离心5 min,弃上清。离心管干燥5 min后加入0.2 mL去酶水,置于55 ℃水浴锅中10 min,制备RNA样品。上机逆转录制备cDNA样品,严格按照试剂盒要求,以GAPDH作为内参,进行肝脏组织中α-SMA、TGF-β1、LIN28A、NF-κBp65、MMP9、COX-2和iNOS的相对含量测定。各基因的引物序列见表1。
2.6 测定大鼠肝脏蛋白表达
采用蛋白免疫印迹法(Western blot)测定肝组织中LIN28A、α-SMA、TGF-β1、NF-κBp65、MMP9、COX-2和iNOS蛋白的表达水平。取各组大鼠肝组织约60 mg,加入含1%PMSF的RIPA裂解液1 mL冰上研磨至无肉眼可见组织,4 ℃条件下12 000 rpm离心10 min后收集上清液。另取少量上清,按照试剂盒说明书通过BCA法测定蛋白含量。按相应比例加入Loading buffer,沸水浴10 min后备用。
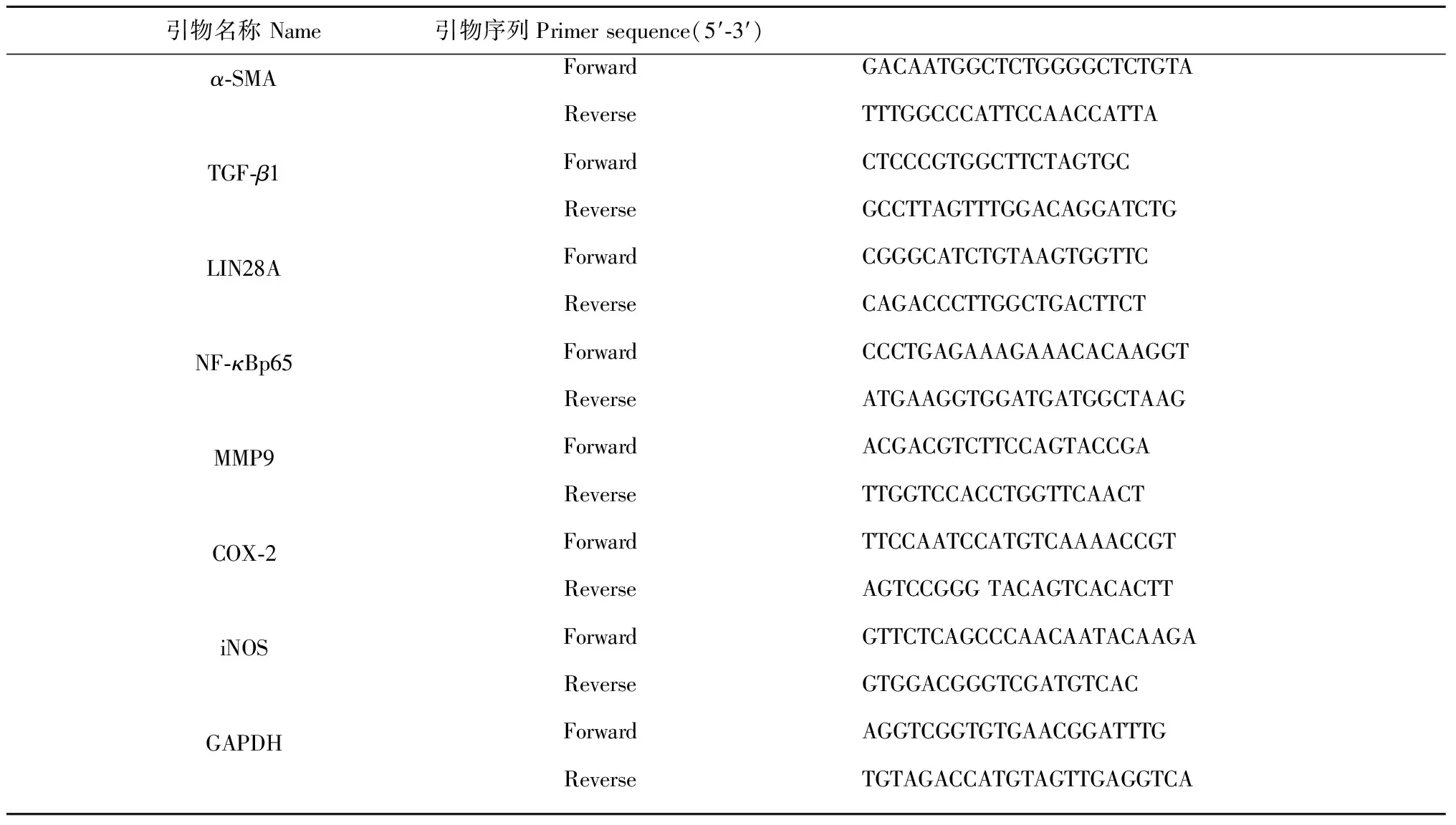
表1 引物序列
根据蛋白含量测定结果,调整蛋白样品上样量后进行10% SDS-PAGE凝胶电泳。于200 mA冰水浴转膜2 h,5%封闭液封闭2 h。加入相应一抗(稀释比例均为1∶1 000),4 ℃下摇床过夜;次日,TBST洗涤,条件同上,根据一抗基源动物加入山羊抗兔或山羊抗鼠二抗(稀释比例为1∶5 000),室温下孵育2 h,TBST洗涤3次,滴加显色液,使用CLINX化学发光成像系统显影,使用Quantity One对条带进行灰度分析并计算蛋白相对表达量。
2.7 数据统计学分析

3 结果与分析
3.1 DCP对HF大鼠肝脏形态的影响
H&E染色结果显示,正常组大鼠肝小叶结构完整,肝细胞排列整齐且饱满。模型组大鼠肝小叶结构受损,且伴大量炎性细胞浸润。相较于模型组,秋水仙素组和DCP低中高剂量大鼠肝小叶清晰,肝细胞变性现象有所改善,表明DCP可改善CCl4造成的大鼠肝脏损伤进而减缓纤维化进程(见图1)。
Masson染色结果显示,正常组大鼠肝脏肝细胞致密排列,无明显蓝色胶原沉积。模型组大鼠肝组织出现大量胶原沉积,有严重的纤维化现象。相较于模型组,秋水仙素组和DCP低中高剂量组肝脏病变有不同程度的减轻,且DCP高剂量组减轻程度明显(见图2)。

图1 大鼠肝脏切片的HE染色(×200)Fig.1 Photomicrograph of rats liver sections stained with H&E (×200)注:A:正常组;B:模型组;C:秋水仙素组;D:DCP-L;E:DCP-M;F:DCP-H;下同。Note:A:Normal group;B:Model group;C:Colchicine group;D:DCP-L;E:DCP-M;F:DCP-H;The same below.

图2 大鼠肝脏切片的Masson染色(×100)Fig.2 Photomicrograph of rats liver sections stained with Masson (×100)
3.2 DCP对HF大鼠血清HA、LN、PC Ⅲ和Ⅳ-C的影响
相较于正常组,模型组大鼠血清中HA、LN、PC Ⅲ和Ⅳ-C含量均有显著升高,差异均有统计学意义(P< 0.01),说明肝细胞通透性改变,提示造模成功。相较于模型组大鼠,DCP低中高剂量组大鼠血清中上述指标均有不同程度的降低,且DCP高剂量组各指标均有明显下降,差异具有统计学意义(P< 0.05),提示DCP可缓解由CCl4造成的大鼠肝细胞损伤(见表2)。

表2 DCP对血清中HA、LN、PC Ⅲ和Ⅳ-C的影响
3.3 DCP对HF大鼠肝脏中IL-1β、IL-6和TNF-α的影响
模型组大鼠肝脏中IL-1β、IL-6和TNF-α含量较正常组大鼠均有显著升高,差异均有统计学意义(P< 0.01)。DCP低中高剂量组大鼠肝脏中IL-1β、IL-6和TNF-α的含量较模型组大鼠均有不同程度的降低,且DCP高剂量组各指标均有明显下降,差异具有统计学意义(P< 0.05),此结果表示DCP可缓解CCl4引起的肝脏炎症反应和炎症介质的释放(见表3)。
表3 DCP对肝脏中IL-1β、IL-6和TNF-α的影响
3.4 PCR检测LIN28A、α-SMA、TGF-β1、NF-κBp65、MMP9、COX-2和iNOS
相较于正常组,模型组大鼠肝组织中α-SMA、TGF-β1、LIN28A、NF-κBp65、MMP9、COX-2和iNOS的mRNA相对含量均显著升高,差异均具有统计学意义(P< 0.01);相较于模型组大鼠,DCP低中高剂量组均可明显抑制上述蛋白的mRNA相对含量,差异均具有统计学意义(P< 0.01),此结果表明DCP抗肝纤维化可能与LIN28A/NF-κBp65信号通路有关(见表4、5)。
表4 DCP对肝脏中α-SMA和TGF-β1mRNA相对含量的影响
表5 DCP对肝脏中LIN28A、NF-κBp65、MMP9、COX-2和iNOSmRNA相对含量的影响
3.5 Western blot检测LIN28A、α-SMA、TGF-β1、NF-κBp65、MMP9、COX-2和iNOS
相较于正常组,模型组大鼠肝组织中LIN28A、α-SMA、TGF-β1、NF-κBp65、MMP9、COX-2和iNOS蛋白表达量均显著升高,差异均具有统计学意义(P< 0.01);DCP低中高剂量组较模型组大鼠均可不同程度抑制上述蛋白的表达,差异均具有统计学意义(P< 0.01),此结果再次验证DCP可能通过LIN28A/NF-κB信号通路起到抗肝纤维化作用(见图3)。

图3 DCP对TGF-β1、α-SMA、LIN28A、NF-κBp65、MMP9、COX-2和iNOS蛋白表达的影响Fig.3 Effect on the TGF-β1, α-SMA, LIN28A, NF-κBp65, MMP9, COX-2 and iNOS protein of DCP
4 讨论
肝纤维化是一种病理病变,以细胞外基质(ECM)在肝周隙的进行性积聚为特征,是慢性肝病的主要问题[10]。由于其最终会导致肝硬化,成为世界范围内发病和死亡的主要原因,据估计全球2%的人口会受到肝硬化的影响[11]。因此,研究DCP对肝脏的保护作用具有重要意义。
该实验采用CCl4诱导肝纤维化,其造成的病理改变与人体临床HF极为相似[12,13]。HA、LN、PC Ⅲ和Ⅳ-C是肝细胞外基质的主要成分,在肝纤维化中由于细胞外基质合成增多、降解不足,故其指标水平升高程度与肝纤维化程度呈现正相关[14]。本研究中,模型组大鼠HA、LN、PC Ⅲ和Ⅳ-C含量明显上升,H&E染色与Masson染色结果显示模型组大鼠肝脏组织脂肪变性和胶原沉积现象明显,提示造模成功。DCP给药后,大鼠血清中HA、LN、PC Ⅲ和Ⅳ-C含量显著下降,H&E染色病理改变有所缓解,显示DCP对肝脏以及肝细胞具有保护作用。
NF-κB是一种核转录因子,调控大量对细胞凋亡和炎症的调节有重要作用的基因[15]。当肝脏发生炎症反应时,NF-κB被激活并释放TNF-α、IL-6和IL-1β等炎症因子,导致肝组织炎症反应加剧[16]。在炎症部位诱导酶COX-2和iNOS表达也会提高,产生过度的炎症反应,也作为炎症标志物。实验结果显示,模型组大鼠肝组织NF-κBp65、COX-2和iNOS的mRNA相对含量和蛋白表达水平显著提高,并且TNF-α、IL-6和IL-1β的mRNA相对含量也有明显升高;相较于模型组,DCP低中高剂量各组大鼠肝组织NF-κBp65、COX-2和iNOS的mRNA相对含量和蛋白表达水平显著降低,炎症因子TNF-α、IL-6和IL-1β的mRNA相对含量也有不同程度下降。此结果表明DCP可通过抑制NF-κBp65蛋白表达,减少下游炎症因子的释放而发挥抗炎作用。
TGF-β1是一种多效细胞因子(促纤维化和炎症因子),参与免疫反应和肝纤维化的发展。TGF-β1可导致肝星状细胞(HSCs)的活化、增殖和纤维化过程[17]。α-SMA可反映HSC的活化情况,作为HSC激活的标志性蛋白,同时也可用于评估肝纤维化程度。基质金属蛋白酶(MMPs)是细胞外基质降解和重构的关键因素。MMP9是降解正常肝基质最相关的MMPs之一,可促进肝纤维化的发展[18]。Western Blot分析显示,模型组大鼠肝组织TGF-β1、α-SMA和MMP9的蛋白表达水平显著提高;相较于模型组,DCP低中高剂量各组大鼠肝组织TGF-β1、α-SMA和MMP9的蛋白表达水平显著降低。此结果表明DCP可通过减少CCl4引起的肝组织TGF-β1和MMP9蛋白表达,减缓肝脏内胶原沉积的速度,从而起到缓解HF的作用。
LIN28A基因可以通过其独特的冷休克结构域(CSD)和半胱氨酸半胱氨酸组氨酸半胱氨酸锌指节结构域的配对被识别,在序列特异性mRNA结合、miRNA结合、miRNA前处理以及miRNA调控基因沉默中起着重要的作用[19]。已有研究证明LIN28A通过NF-κBp65信号通路参与炎症反应[19],并且降低NF-κBp65的表达有助于缓解HF。在实验中,相较于正常组,模型组大鼠肝脏中LIN28A和NF-κBp65的蛋白表达水平和mRNA相对含量显著提高,且DCP低中高剂量各组大鼠肝组织中LIN28A和NF-κBp65的蛋白表达水平和mRNA相对含量显著降低且呈现出浓度依赖性。以上结果表明HF炎症产生的过程可能与LIN28A的激活有关,结合DCP可以抑制LIN28A和NF-κBp65的表达,并抑制NF-κBp65下游COX-2、iNOS、TNF-α、IL-6和IL-1β等炎症因子的表达,推测DCP抗炎和治疗HF的机制可能与通过抑制LIN28A/NF-κB信号通路有关。
综上所述,DCP对大鼠HF具有明显改善作用,其药效作用机制可能通过调控LIN28A/NF-κB信号通路,阻止炎症反应实现。
猜你喜欢
中老年保健(2022年4期)2022-11-25
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国现代医生(2022年21期)2022-08-22
传染病信息(2022年3期)2022-07-15
传染病信息(2022年3期)2022-07-15
肝博士(2022年3期)2022-06-30
中国药学药品知识仓库(2022年9期)2022-05-23
现代临床医学(2022年1期)2022-02-12
湖南畜牧兽医(2021年6期)2022-01-24
食品安全导刊(2021年21期)2021-08-30
