
醉鱼草内生细菌ZJ1的抑菌作用及其脂肽类抑菌代谢产物鉴定
2021-04-01 11:58刘晓峰周建波张宝俊赵晓军
草地学报 2021年3期
任 璐, 刘晓峰, 周建波, 殷 辉, 张宝俊, 赵晓军
(山西农业大学植物保护学院, 山西 太原 030031)
醉鱼草(BuddlejalindleyanaFortune)属马钱科醉鱼草属,为多年生小灌木,因其全株有毒,捣碎投入河中能使活鱼麻醉而得名[1]。醉鱼草属植物约有100种,中国分布50余种,其分布广泛、生长迅速,是天然的中草药,具有祛风杀虫和活血化瘀的功效。醉鱼草的主要成分有黄酮类、三萜类、环烯醚萜苷类等,具有抗菌杀虫、抗氧化等生物活性[2],其提取物对蝗虫、小菜蛾幼虫、菜青虫幼虫、朱砂叶螨及线虫均有一定杀虫活性[3]。
生物防治因其对人类和环境友好弥补了化学农药的不足,成为目前非常有潜力的研究方向。植物内生细菌作为生物农药的天然资源菌,其种类丰富且广泛存在于几乎所有高等植物中,通过与植物相互作用提高植物抗病性[4]。并且内生细菌在生存和繁殖的过程中产生多种次生代谢物,这些物质对于植物病害的防治起到积极作用。其中,芽胞杆菌Bacillus是一种革兰阳性细菌,其生长繁殖迅速,可以有效定殖在植物根际,能产生多种活性代谢物质,不仅具有较强的抗逆性,还对多种植物病害具有良好的防治效果[5-6]。
植物内生细菌的主要抗病机制之一是产生抗菌物质,抗菌物质种类多样为生物农药开发提供了丰富多样的先导化合物。其中,目前研究已经发现的抗菌物质主要包括脂溶性抗生素[7]、环脂肽类抗菌物质[8-9]、抗菌蛋白[10-12]、嗜铁素[13]、几丁质酶[14-15]、黄酮类物质[16]、多肽类物质以及有机酸类[17]等,其中脂肽类抗菌物质和抗菌蛋白的报道较多。但在抗菌物质种类鉴定和开发应用方面,植物内生细菌与其它微生物、植物活性成分以及生物医药开发等研究相比,还处于起步阶段。
目前,关于醉鱼草内生细菌的研究还鲜有报道,鉴于此,本研究以醉鱼草为研究对象,从醉鱼草茎中筛选出1株对多种植物病原真菌具有良好抑菌作用的内生细菌ZJ1,鉴定为贝莱斯芽孢杆菌(Bacillusvelezensis)。并在此基础上,探讨醉鱼草内生细菌ZJ1的抑菌特性、抗菌物质种类及其生防机制,旨在为醉鱼草内生细菌田间应用技术开发和后期市场化奠定基础。
1 材料与方法
1.1 内生细菌及植物病原真菌
内生细菌ZJ1从醉鱼草茎部分离获得,经鉴定为贝莱斯芽孢杆菌(Bacillusvelezensis);10种供试植物病原真菌,分别为番茄灰霉病菌(Bortytiscinerea),番茄早疫病菌(Alternariasolani),番茄枯萎病菌(Fusariumoxysporumf.sp.lycopersici),黄瓜枯萎病菌(F.oxysporumf.sp.cucumerinum),胡萝卜褐腐病菌(Leptosphaerialibanotis),核桃腐烂病菌(Cytosporajugandis),棉花立枯病菌(Rhizoctoniasolani),苹果腐烂病菌(Valsamali),桃褐腐病菌(Moniliniafructicola),西瓜枯萎病菌(F.oxysporumf.sp.niveum)均由山西农业大学植物保护学院植物化学保护实验室保存。
1.2 内生细菌ZJ1抑菌活性测定
采用平板对峙法测定ZJ1菌株对10种供试病原菌的抑菌率。将供试病原菌接种于PDA平板中央,在距其20 mm四侧对称接种ZJ1菌株,每种病原菌重复3次。26℃倒置培养5 d后,观察有无抑菌作用,测量病原菌菌落直径,计算抑菌率。
采用生长速率法测定ZJ1菌株发酵液的抑菌率。ZJ1菌株在160 r·min-1,28℃恒温条件下,在LB培养液中振荡培养72 h后,12 000 r·min-1,4℃离心12 min,去除菌体得到发酵液,分别进行121℃灭菌处理40 min和0.22 μm微孔滤膜过滤处理,然后1∶9(V∶V)分别与PDA混匀制备平板,接种病原菌,每试验重复3次。26℃倒置培养5 d,根据以下公式计算抑菌率。

1.3 内生细菌ZJ1代谢物物理性质测定
1.3.1排油能力测定 取9 mL蒸馏水水平放置于培养皿中央,取1 mL柴油缓慢滴加到液体表面形成一层薄油膜,取10 μL的ZJ1菌株发酵液轻落于油膜中心,观察是否有油膜被排挤向四周形成排油圈的现象。
1.3.2液滴坍塌测定 在水平放置的parafilm膜上面分别轻滴20 μL的ZJ1发酵液和蒸馏水,观察二者与parafilm膜的接触角,观察是否有液滴坍塌现象产生,如有则表明其具有一定的表面活性。
1.4 内生细菌ZJ1脂肽抗菌物质的提取
将ZJ1菌株26℃培养活化36 h后,挑取一环单菌落接种于LB培养液中,28℃,160 r·min-1振荡培养24 h得到内生细菌ZJ1种子液。取ZJ1种子液按1% (V/V)的接种量接入BPY培养液中,在160 r·min-1,28℃黑暗条件下振荡培养72 h,于12 000 r·min-1离心12 min,去除菌体得到上清液。
上清液中脂肽类代谢物的提取参照高振峰[18]的盐酸沉淀、甲醇抽提法进行,得到黄色晶体,将黄色晶体用无菌水溶解,0.22 μm滤膜过滤后得到脂肽粗提取物,并于—20℃保存备用。
1.5 脂肽粗提物抑菌活性检测
采用琼脂扩散法对脂肽粗提物抑菌活性进行测定:取100 μL供试病原菌孢子悬浮液(孢子浓度为1.6×106cfu·min-1)涂布于PDA平板表面,表面干燥后每皿用打孔器打2孔,分别加入80 μL脂肽粗提物和无菌水,每试验重复3次,于28℃恒温培养7 d,测定其抑菌圈直径。
1.6 脂肽粗提物的分离纯化及鉴定
分别采用制备型中压层析和制备型HPLC对脂肽粗提物进行分离纯化,采用琼脂扩散法测定各收集峰抑菌活性。筛选出活性峰后,采用液质联用仪(Thermo Fisher,型号:LTQ XL)对活性峰中脂肽类物质进行质谱鉴定。液相条件、洗脱程序及质谱条件参考高振峰[18]的方法。
上述抑菌活性的检测均以A.solani为靶标菌。
1.7 内生细菌ZJ1发酵液抑菌机制的初步研究
1.7.1对病菌菌丝生长及形态的影响 利用正置显微镜对1.2抑菌活性测定中抑制率较高的B.cinerea菌丝形态进行观察。
1.7.2对病菌细胞膜透性的影响 将B.cinerea接种于无菌的PD培养液中,160 r·min-1,28℃黑暗振荡培养7 d后,12 000 r·min-1离心后弃上清液,用无菌水清洗菌体3~5次,分别称取5 g置于浓度为2%,5%,10%,15%,20%,50%,100%(V/V)的ZJ1发酵液和无菌水混合液中,以无菌水为对照。160 r·min-1,28℃振荡培养,每1~2 h取样离心检测上清液电导率值。
1.8 数据统计分析
采用SPSS 18.0统计软件进行方差分析,应用Duncan氏新复极差法进行各处理间差异显著性检验。
2 结果与分析
2.1 内生细菌ZJ1抑菌活性测定
ZJ1菌株对10种供试病原菌均有不同程度抑制作用,如图1、表1数据所示,抑制率为52.63%~95.69%。其中ZJ1菌株对M.fructicola的抑制率最高,达到95.69%,其次是对B.cinerea和C.jugandis的抑制率较高,分别达到78.38%和77.39%,对F.oxysporumf.sp.niveum和F.oxysporumf.sp.cucumerinum的抑制率最低,为52.63%和54.27%,对于其他病原菌的抑菌率在60.55%~66.33%之间。

表1 ZJ1菌株对10种病原菌的抑制作用Table 1 The inhibition of strain ZJ1 on 10 kinds of pathogenic fungi
ZJ1菌株发酵液经过两种处理方式后对10种供试病原菌均有不同程度抑制作用。如表2所示,过滤后ZJ1代谢物对10种病原真菌的抑制作用有显著差异,抑菌率在32.39%~94.68%之间。其中对M.fructicola的抑制率最高,为94.68%;其次对F.oxysporumf.sp.niveum,L.libanotis和C.jugandis3种病原菌的抑制率较高,分别为81.10%,82.55%和88.52%;而对F.oxysporumf.sp.lycopersici的抑制率最低,仅为32.39%。经121℃高温处理后发酵液的抑菌率仅为13.55%~50.55%,其中仅对C.jugandis的抑菌率为50.55%,对V.mali,M.fructicola和L.libanotis的抑制率较低,在20%~40%之间,对其余病原菌的抑制率均在20%以下。
2.2 内生细菌ZJ1菌株代谢物物理性质测定
2.2.1排油能力测定 滴加10 μLZJ1菌株发酵滤液后油膜缓慢向四周扩散,油膜被挤向四周产生φ为45 mm的排油圈(图2左)。
2.2.2液滴坍塌测定 蒸馏水液滴(左)与parafilm膜接触面较小,成球状;而ZJ1菌株发酵滤液液滴(右)接触面较大,发生坍塌失去球状(图2右),说明ZJ1菌株发酵滤液具有一定的表面活性。
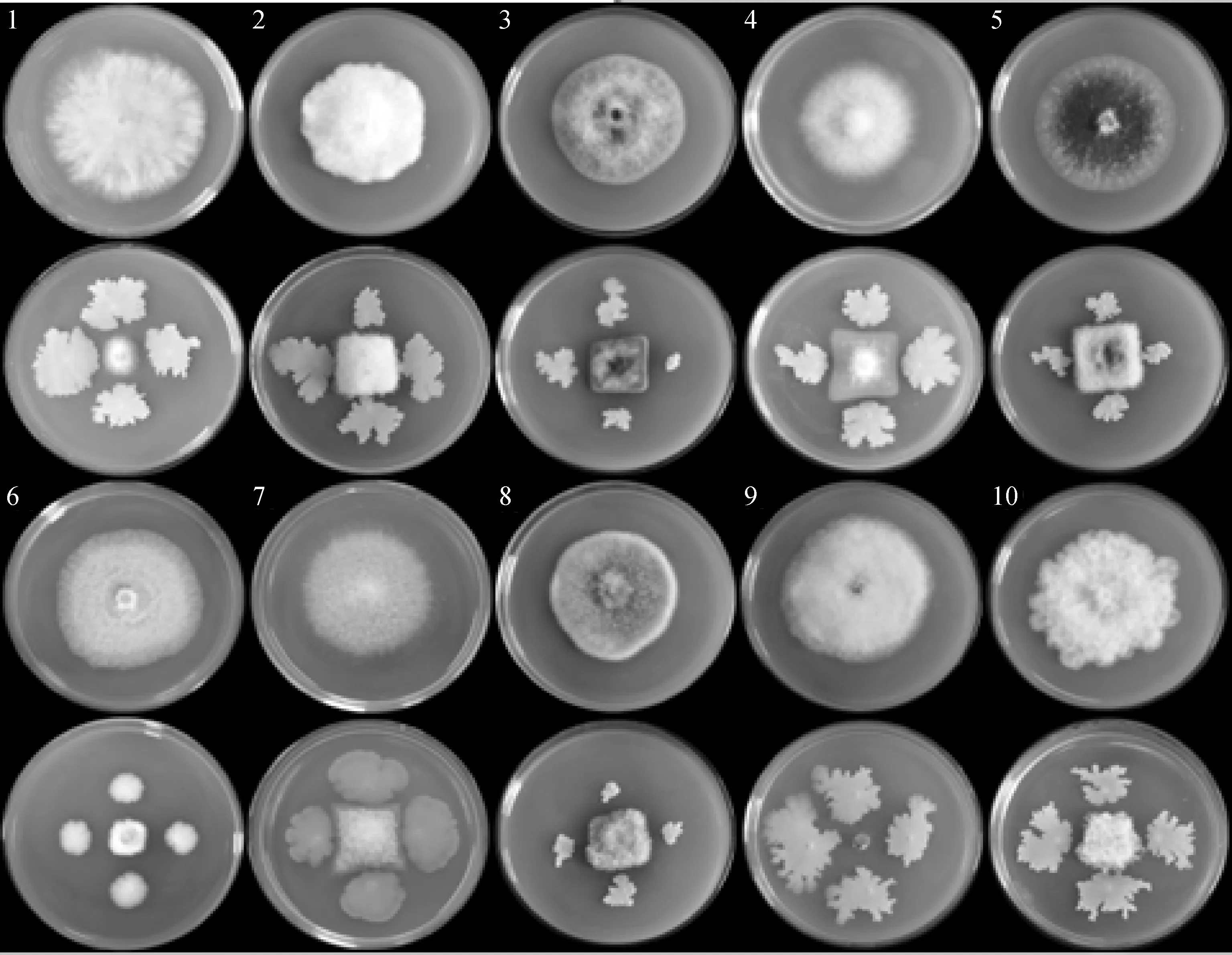
图1 ZJ1菌株对10种病原菌的抑制作用Fig.1 The inhibition of strain ZJ1 on 10 kinds of pathogenic fungi注:1~10分别为番茄灰霉病菌、番茄枯萎病菌、番茄早疫病菌、黄瓜枯萎病菌、胡萝卜褐腐病菌、核桃腐烂病菌、棉花立枯病菌、苹果腐烂病菌、桃褐腐病菌、西瓜枯萎病菌Note:1~10 represent B. cinerea,F. oxysporum f.sp. lycopersici,A. solani,F. oxysporum f.sp. cucumerinum,L. libanotis,C. jugandis,R. solani,V. mali,M. fructicola,F. oxysporum f.sp.niveum,respectively
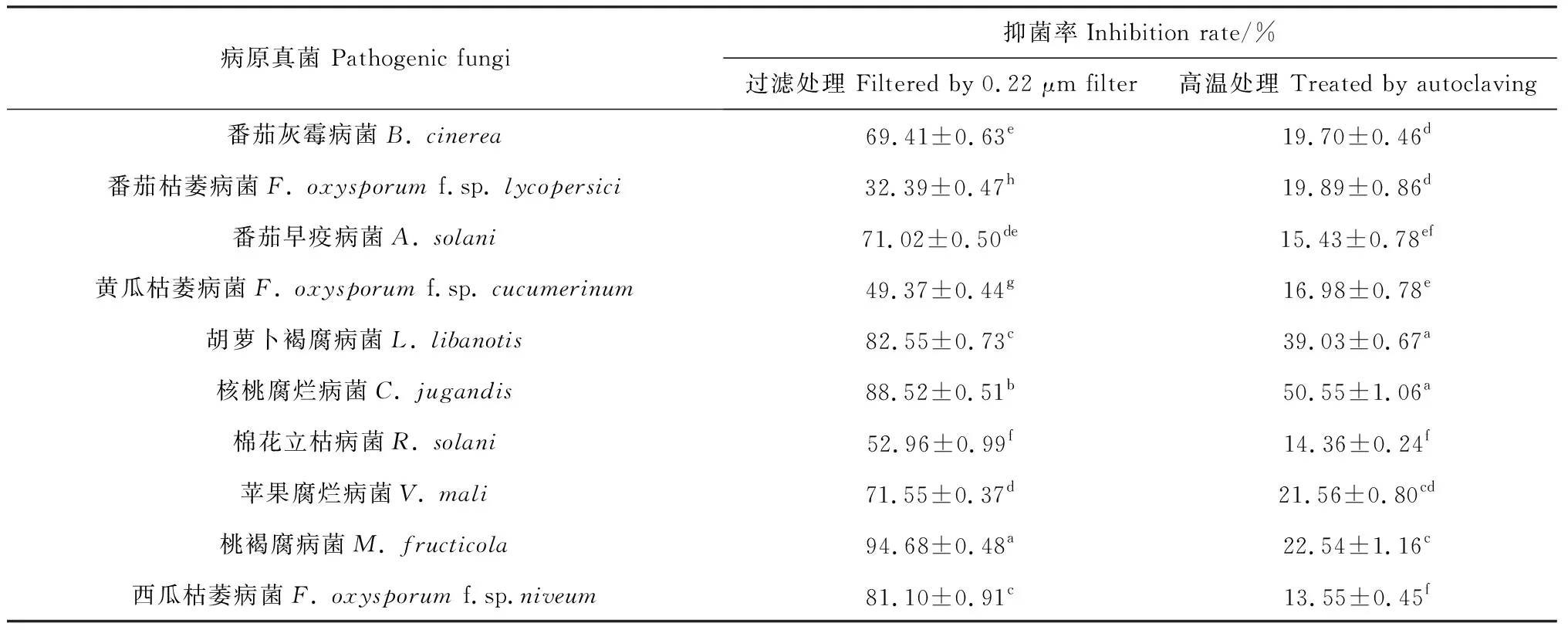
表2 ZJ1菌株发酵液对10种病原菌的抑制作用Table 2 The inhibition of fermentation broth of strain ZJ1 on 10 kinds of pathogenic fungi

图2 菌株ZJ1代谢物排油能力和液滴坍塌测定Fig.2 The oil discharge capacity and droplet collapse of ZJ1 metabolite
2.3 内生细菌ZJ1脂肽抗菌物质的分离、纯化及鉴定
2.3.1ZJ1菌株脂肽粗提物抑菌活性 以A.solani为靶标,如图3所示,ZJ1菌株脂肽粗提物具有较好抑菌活性,可形成明显抑菌圈,抑菌圈直径为24.8 mm。
2.3.2ZJ1菌株脂肽粗提物分离、纯化 如图4,5所示,在明确菌株ZJ1脂肽粗提物的抑菌活性后使用中压层析仪对其进行分离纯化,结果发现在检测波长为211 nm下,共检测出7个吸收峰;抑菌活性测定发现,仅峰7具有较好的抑菌活性,抑菌圈直径为21.6 mm,其余吸收峰均无抑菌活性。
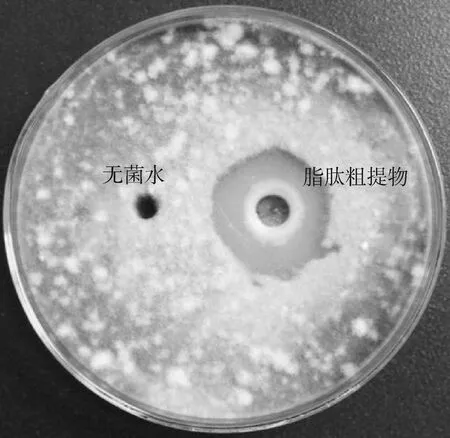
图3 ZJ1脂肽粗提物对番茄早疫病菌的抑菌效果Fig.3 Inhibitory effect of strain ZJ1 lipopeptide crude extracts on A. solani
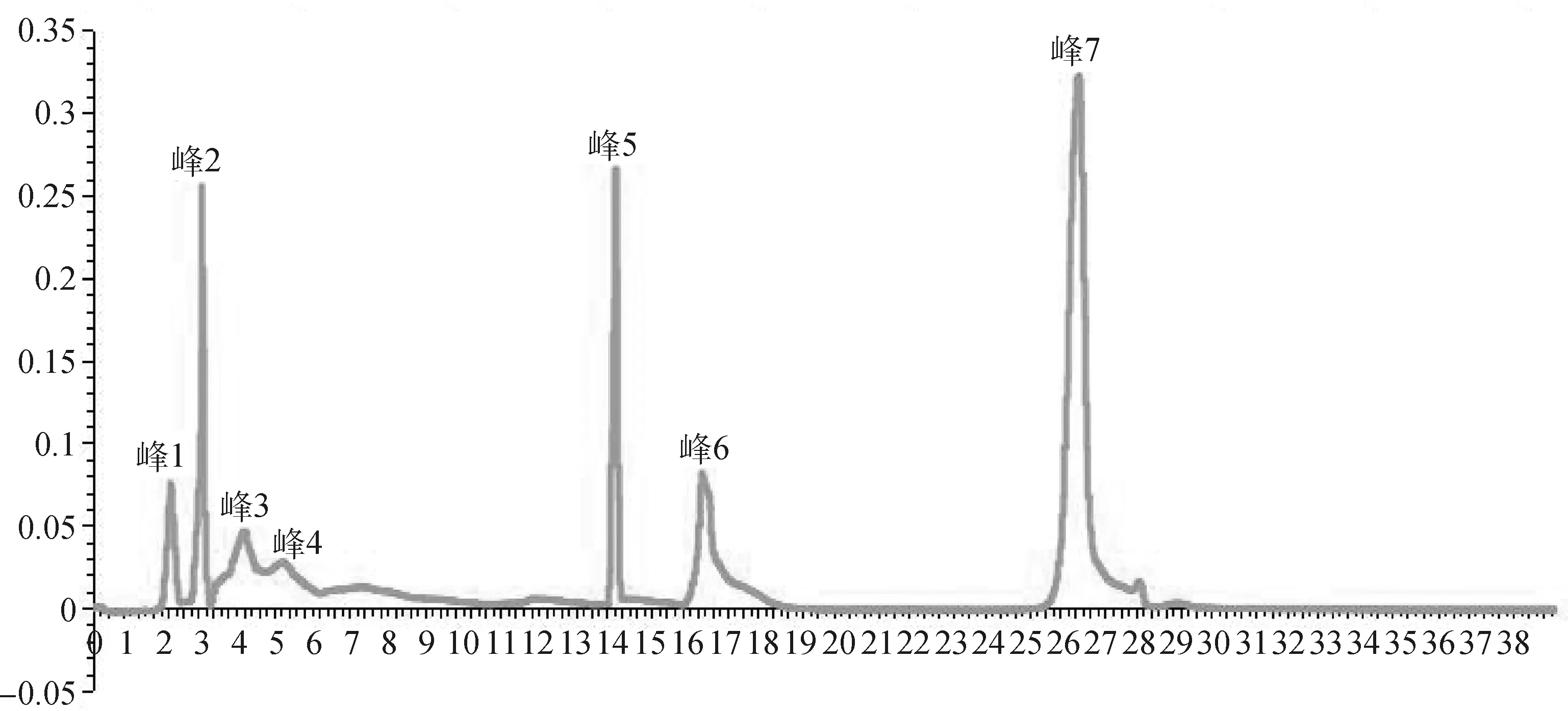
图4 ZJ1菌株脂肽物质中压层析活性峰HPLC纯化图谱Fig.4 The HPLC purification map of active peak purified by medium pressure chromatography of the lipopeptide of strain ZJ1
2.3.3ZJ1菌株脂肽物质的质谱鉴定 如图6,7所示,使用LC-MS对活性峰峰7进行质谱鉴定,结果发现活性峰峰7在保留时间3.90 min处具有较大吸收峰,且活性峰杂峰较少;对保留时间3.90 min下的质谱图进行查看,菌株ZJ1在m /z值为1 030.00,1 044.10 和1 058.01 处有离子峰(簇)出现,且分子量之间相差为14(CH2)出现的3个离子峰同表面活性素(Surfactin)家族相似;另外,还发现菌株ZJ1在m /z值为1 453.85,1 467.35,1 481.55和1 495.49处同样有离子峰(簇)出现,且分子量之间相差为14(CH2)出现的4个离子峰同泛革素(Fengycin)家族相似。
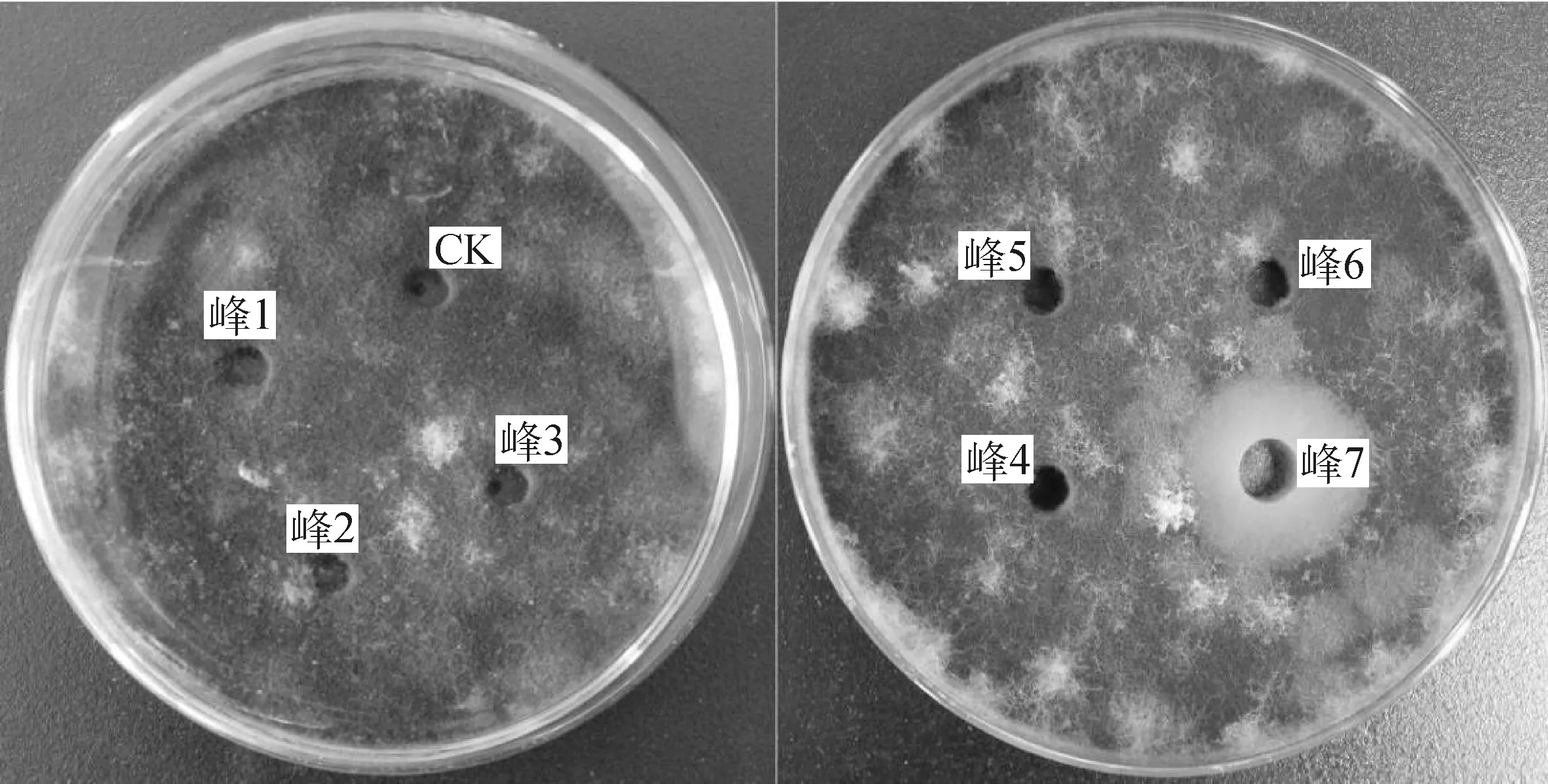
图5 ZJ1菌株脂肽粗提物中压层析各吸收峰对番茄早疫病菌抑菌效果Fig.5 The antifugnal activity of all absorption peak of lipopeptide purified by medium pressure chromatography of strain ZJ1 against A. solani
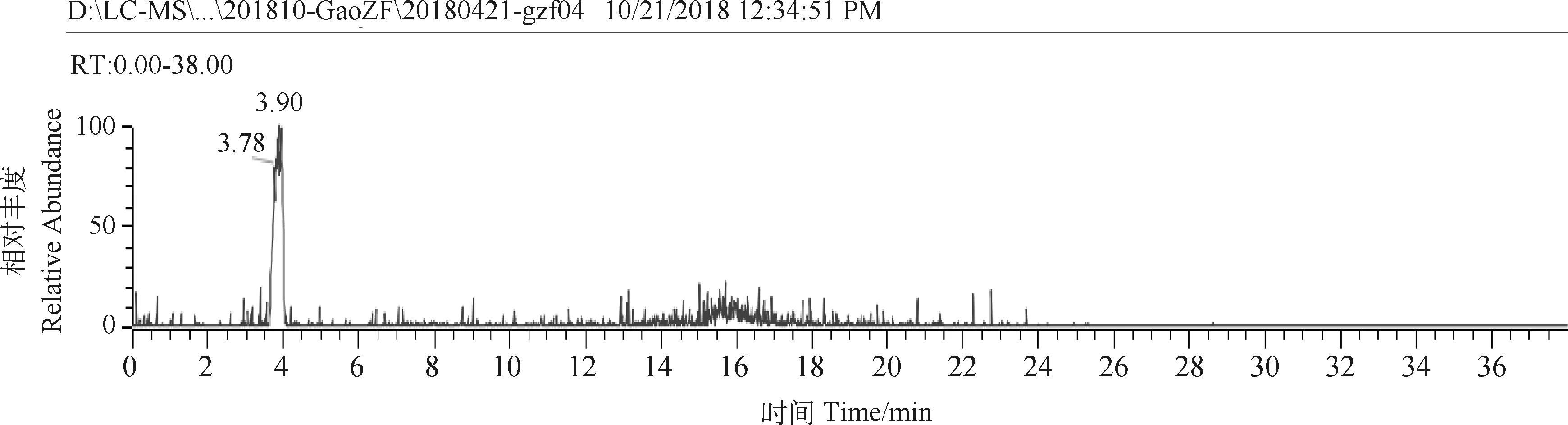
图6 ZJ1菌株脂肽抗菌物质LC检测图谱Fig.6 The LC detection map of antifugnal lipopeptide material of strain ZJ1

图7 ZJ1菌株脂肽抗菌物质保留时间3.90 min时MS图谱Fig.7 The MS map of of antifugnal lipopeptide material of strain ZJ1 at the time of 3.90 min
2.4 拮抗细菌ZJ1发酵液抑菌机制初步研究
2.4.1对病菌菌丝生长及形态的影响 ZJ1菌株代谢物对B.cinerea菌丝生长形态影响明显,处理后的B.cinerea产孢极少,菌丝生长异常,明显断裂且折叠分叉(图8)。
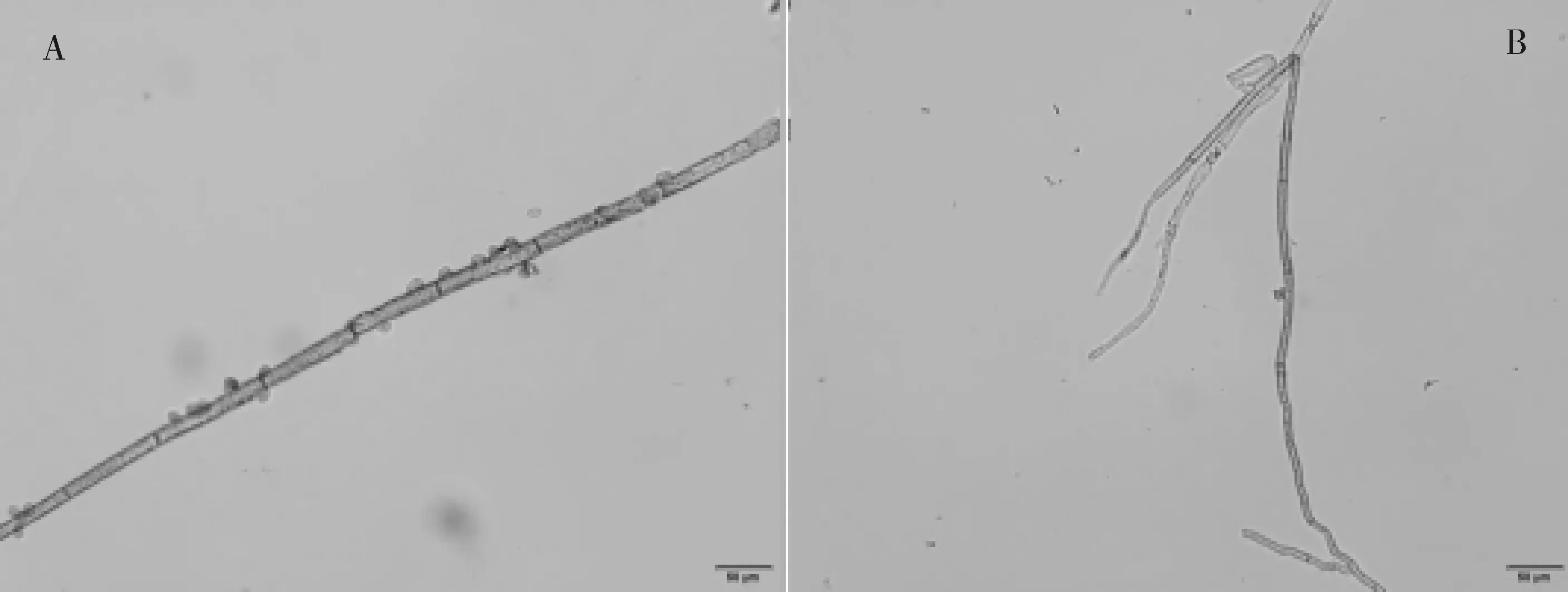
图8 ZJ1菌株发酵滤液对B. cinerea菌丝生长的影响Fig.8 Inhibiting effect of actinomycete strain ZJ1 on mycelium growth of B. cinerea注:图中A为处理前B. cinerea菌丝生长形态,B为处理后B. cinerea菌丝生长形态Note:A represent mycelium growth morphology of B. cinerea;B represent mycelium growth of B. cinerea after treatment
2.4.2对病菌细胞膜透性的影响 如图9所示,在不同时刻,对照组电导率(ms/cm)随时间的推移未出现较大变化。不同浓度发酵液处理后电导率值虽略有波动但总体均逐渐呈现小幅上升趋势,且上升幅度与发酵液的浓度呈正相关。在同一时刻,对照组与5%发酵液处理后菌体的电导率值较为相近,与其它浓度发酵液处理后的电导率有较大差异,且数值与浓度呈正相关。该结果表明ZJ1菌株代谢产物对B.cinerea的细胞膜透性会产生一定程度的影响,破坏程度会随着发酵液浓度和时间的增加而加剧。

图9 不同浓度的ZJ1菌株发酵滤液对B. cinerea细胞膜透性的影响Fig.9 The effect of different concentration of fermentation filtrate of ZJ1 on cell membrane permeability B. cinerea注:图中2,5,10,15,20,50,100表示ZJ1菌株发酵液的体积分数(V/V),PD表示PD培养基对照Note:2,5,10,15,20,50 and 100 in the figure represent the volume fraction of ZJ1 fermentation broth(V/V),PD represent PD medium control
3 讨论
大量研究发现,植物内生菌对植物病原真菌具有较强的抑菌作用。李乐溪等[19]从大蒜(AlliumsativumL.)蒜瓣中分离得到对5种病原菌有较好抑制作用的内生细菌和放线菌19株;李玲玲等[20]从银杏(Ginkgobiloba)茎叶中分离得到12株内生真菌、3株内生放线菌和4株内生细菌,均对金黄色葡萄球菌Staphylococcusaureus有较强抑菌活性。本研究从醉鱼草茎中分离并筛选出1株抑菌活性较强的内生细菌ZJ1,对M.fructicola、B.cinerea和C.jugandis的抑制率较高,达到75%以上。ZJ1具有抑菌广谱性和较强的抑菌活性,这与陈奕鹏[21]从香蕉(MusananaLour.)根部分离出的内生细菌,蒋晶晶[22]分离得到的葡萄(Vitisvinifera)内生细菌抑菌结果相似,均对多种植物病原真菌具有良好抑菌效果。对ZJ1菌株发酵液高温处理后,发现抑菌率明显降低,对M.fructicola,B.cinerea的抑菌率降到20%以下,说明高温可以降低内生细菌代谢物的活性。推测ZJ1菌株发酵液的抑菌成份中含有抗菌蛋白,但本研究未对抗菌蛋白进行分离和鉴定,还需进一步研究确定。
植物内生细菌的抑菌机理之一就是利用自身产生的抗菌物质来发挥抗病作用,微生物在适宜培养条件下,可分泌脂肽、磷酯类、糖脂类等多种化合物的次生代谢产物,使其具有一定表面活性。黄华毅等[23]从枣树根际土壤中分离获得1株枯草芽孢杆菌STO-12,杨柯[24]从海芦笋(SalicorniabigeloviiTon.)中分离得1株内生细菌YK-HLS-11,发现其次生代谢产物均为脂肽类化合物。大量研究表明,芽孢杆菌在生长后期产生的主要抗菌物质是脂肽类物质。本试验前期采用便捷、准确的排油圈法和液滴坍塌法对ZJ1菌株的抗菌物质进行初步鉴定,结果得到该菌株代谢物可产生较大排油圈,与水相比表面张力小,出现坍塌现象,说明该菌株含有表面活性成分,从而初步推测可含有大量脂肽类物质。
为进一步明确ZJ1菌株的抑菌物质,采用制备型HPLC与质谱分析对其进行分离纯化和结构鉴定,证实ZJ1菌株可产生表面活性素(Surfactin)和泛革素(Fengycin)。高振峰[18]从梓树(CatalpaovataG. Don.)中分离得到1株产脂肽类化合物的内生细菌ZSYB-1,鉴定其脂肽类代谢物主要为伊枯草菌素itruin A。目前发现的该类物质主要有表面活性素(Surfactin A,B,C)、伊枯草菌素(Iturin A,C和杆菌抗霉素L,D,F等)和泛革素(Fengycin A,B和制磷脂菌素)[25]。脂肽类物质具有较强的表面活性作用,植物体内的芽孢杆菌通过改善表面张力来改变生长环境,以便于形成菌群快速移动,方便抑菌物质的运输、储存和在植物表面定殖,并且使其具有或强或弱的的抗菌活性。脂肽抗菌物质与抗菌蛋白相比,具有较好的耐热、耐酸碱和耐紫外特性,应用潜力良好。本研究在ZJ1菌株抑菌物质成分的基础上,后续可对分子结构进行确定,以期推动开发环境友好型生物农药提供应用依据。
内生细菌的抑菌机理是在生长过程中所分泌的次生代谢物可以干扰、抑制另一种生物的生长发育而自身却不受影响,其作用于细胞,破坏菌体细胞的完整性,还会阻碍能量物质合成引起歧变,出现肿胀破裂造成原生质泄露[26]。周金伟等[27]从烟草(NicotianatabacumL.)中分离的短短芽胞杆菌011主要是导致A.solani菌丝生长异常,抑制孢子萌发。宁爽[28]从植物茎、叶部分离的假单胞菌BTa14、Bar25的发酵液可导致苹果斑点落叶病菌Alternariamali、苹果轮纹病菌Physalosporapiricola等病原菌菌丝膨大、扭曲、畸形从而影响病菌生长。张丽[29]从印楝(AzadirachtaindicaA.Juss.)叶片和果实中筛选出的内生放线菌YL-2的抑菌活性物质对稻瘟病菌Magnaportheoryzae菌丝生长、病原孢子萌发抑制率可达90%以上。本试验观察B.cinerea经ZJ1菌株发酵液处理后的生长形态,发现该菌株具有相似的抑菌机制。
细胞膜是病原菌的保护屏障,在遇到抑菌活性强的拮抗因子遭到破坏时,保护屏障便会打开,内部电解质外泄到培养液中,促使培养液中电导率上升[30]。陈忠军等[31]从发酵食品中分离出2株乳杆菌,发现其抑菌物质可破坏酵母细胞膜通透性。石玉星[32]从曼陀罗(DaturastramoniumLinn.)组织中分离出一株内生细菌MY1,发现其可破坏谷瘟病菌的细胞膜通透性,使电导率上升。本研究发现经ZJ1菌株处理后,对B.cinerea的细胞膜透性产生了较大影响,使大分子物质外渗到胞外,且这种破坏程度会随着发酵滤液浓度和时间的增加而加剧。在后续的研究中,将继续对其分泌抗菌物质的种类、分子组成等进行探索,以明确其抑菌机制。
4 结论
醉鱼草内生细菌ZJ1菌株及其发酵液对10种供试病原真菌均有良好的抑菌作用,对Moniliniafructicola的抑制率最高,达95.69%和94.68%。测定ZJ1菌株代谢物的物理特性发现其具有排油能力且出现液滴坍塌现象,推测ZJ1菌株代谢物中含有脂肽类化合物。对ZJ1菌株的脂肽类代谢产物进行分离纯化和鉴定,发现其含有表面活性素和泛革素。经ZJ1菌株代谢物处理后的Botrytiscinerea,产孢极少或不能正常产孢,菌丝生长异常,发生断裂。随着处理时间延长,B.cinerea细胞膜通透性增大,渗透物质流出,电导率上升。本研究结果为醉鱼草内生细菌ZJ1的开发和利用奠定了基础。
猜你喜欢
湖北农机化(2020年4期)2020-07-24
山东化工(2020年4期)2020-03-30
世界农药(2019年4期)2019-12-30
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
今日农业(2019年11期)2019-08-15
分析科学学报(2019年2期)2019-04-25
乡村地理(2018年2期)2018-09-19
天然产物研究与开发(2018年4期)2018-05-07
中成药(2018年1期)2018-02-02
物理化学学报(2015年7期)2015-12-30
