
放牧和氮添加对半干旱沙质草地优势种糙隐子草及群落功能性状的影响
2021-04-01 11:58孙一梅郭爱霞张森溪左小安
草地学报 2021年3期
孙一梅, 田 青*, 郭爱霞, 张森溪, 吕 朋, 左小安,4
(1. 甘肃农业大学林学院, 甘肃 兰州 730070; 2. 中国科学院西北生态环境资源研究院, 乌拉特荒漠草原研究站, 甘肃 兰州 730000;3. 中国科学院大学, 北京 100049; 4. 中国科学院西北生态环境资源研究院奈曼沙漠化研究站, 甘肃 兰州 730000)
草地作为陆地生态系统的重要组成部分,占我国国土面积的41.7%[1]。过去几十年,由于不合理的人类活动和剧烈的气候变化导致科尔沁沙地沙漠化进程加快,生态环境不断恶化,使其成为我国北方农牧交错带沙漠化最为严重的地区之一[2]。放牧作为草地利用方式之一,能直接或间接地影响植物群落组成、结构、功能性状及生产力[3-4],同时氮沉降作为全球变化的主要内容之一,对半干旱沙质草地植物光合能力和养分利用效率影响较大[5]。有学者发现放牧和氮添加交互作用可能对草地生态系统存在影响,如Hodgkinson等[6]发现食草动物通常会放大资源对植物的影响;Gao等[7]发现在低耐牧性和高养分环境下,放牧与养分间表现为拮抗作用;刘思洋等[8]则发现围封和低氮交互作用会促进植被恢复和提高植物多样性,但高氮会降低植物多样性和系统稳定性。因此,探究放牧和氮添加对半干旱沙质草地植物功能性状的影响能更好地为未来草地的合理利用提供理论依据。
放牧和氮添加条件下植物通过改变各功能性状来响应和适应环境的变化,如高度、叶面积、比叶面积、叶干物质含量及叶片氮含量等,这些性状容易测定且与植物光合能力及资源获取能力密切相关,因此可以很好地预测生态系统功能[9]。从不同研究水平看,种群水平的功能性状更多地受基因和表型可塑性的影响,而群落水平上的功能性状则更多地受环境影响[10]。另外,群落中优势种的功能性状对表征生境变化和群落演替方向具有一定的指示作用[11],反映出物种水平功能性状与群落水平基本行为和功能间的密切关系[12-13]。糙隐子草(Cleistogenessquarrosa)为多年生丛生小禾草,属C4植物,作为科尔沁沙质草地的优势种,其水分利用效率、耐牧性、抗旱能力及光合能力等优于与其共存的C3植物[14]。研究发现,放牧可以增加糙隐子草总叶面积和叶片数,降低其单叶面积和株高[15],氮添加可以影响其高度、比叶面积及根系深度等。以往相关研究多以植物个体或群落功能性状对放牧或气候变化单个响应机制[16-18]为主,很少有研究基于种群和群落水平来探究功能性状对放牧和氮添加交互作用的响应。因此,本研究以科尔沁沙地优势种糙隐子草及其所在群落为研究对象,通过野外控制试验,探究半干旱沙质草地优势种糙隐子草和群落功能性状对放牧和短期氮添加的响应,进一步揭示优势种糙隐子草功能性状与其所在群落性状间的关系,从而增进氮沉降背景下草地生态系统如何响应放牧干扰的理解。
1 材料与方法
1.1 研究区概况
研究区(42°55′~42°57′ N,120°40′~120°43′ E)位于内蒙古通辽市奈曼旗境内,地处科尔沁沙地中南部,属温带大陆性半干旱季风气候,海拔约360 m,年均气温约6.4℃。全年≥10℃的有效积温在3 000℃以上,无霜期150 d,年均降水量约360 mm,其中70%~80%的降水集中在6—8月,年均蒸发量约1 972.8 mm。年均风速3.2 ~ 4.1 m·s-1,春冬两季盛行西北风,夏秋两季为西南风[19]。该研究区流动沙丘、半固定沙丘、固定沙丘、丘间低地等各种沙丘类型交错分布,土壤类型以风沙土、沼泽土、沙质栗钙土为主。试验区主要植物有:糙隐子草(Cleistogenessquarrosa)、猪毛蒿(Artemisiascoparia)、砂蓝刺头(Echinopsgmelini)、尖头叶藜(Chenopodiumacuminatum)、白草(Pennisetumcentrasiaticum)、狗尾草(Setariaviridis)等。
1.2 试验设计
在中国科学院西北生态环境资源研究院奈曼沙漠化研究站开展试验。于2019年4月底选取长期封育(F,1996年封育)和放牧(G,2014年开始放牧处理)草地,放牧草地为封育草地附近无围栏保护的公共自由放牧区。放牧强度为1头牛·hm-2,属于中度放牧[20]。放牧时间选择在非生长季(9月—来年4月),生长季(5—8月)禁牧。在围封和放牧区分别设置2个氮素水平:不施氮(N0)和添加氮(N+),小区面积为6 m×6 m,每个处理6次重复,共24个小区,试验小区在样地内随机布设,小区之间设置2 m的缓冲带。氮添加量参考国际上常用的氮素添加水平20 g·m-2·a-1[21],分两次施加,每次10 g·m-2·a-1。施肥时间选择5月和7月中旬,氮肥施用量根据小区面积和尿素含氮量(46%)计算。为减小氮素侧向干扰,在每个试验小区的四周挖1 m深,用包有塑料纸的金属隔板隔开。研究选取的样地及试验处理位于连续的同一地段,地势平坦,环境条件相对均质,有效地避免了空间异质性的干扰。
1.3 性状测定
于2019年8月生物量高峰期进行调查取样,在研究区样地的每个小区内随机设置3个1 m×1 m的样方,调查样方中植物群落物种组成、盖度和高度。采用齐地面刈割法分物种收获每种植物,装入信封袋带回实验室,60℃烘干至恒重后称重。采样时为了避免边缘效应,限制采样点离边界至少1 m。选择糙隐子草(种群水平)及其群落内其它共优种(群落水平)作为研究对象,每种随机选取5 ~ 10株植物,选择10 ~ 20片完全伸展且无病虫害的成熟叶片,用剪刀剪下,置于湿润的滤纸之间,迅速放入自封袋储藏于便携式车载冰箱内(内部温度< 5℃)。回实验室后将叶片浸入水中,处于黑暗环境下冷藏(5℃)12 h。取出叶片用纸迅速吸去叶表水分,在百万分之一的电子天平上称得采样叶片的总叶饱和鲜重。然后将叶片平铺在扫描仪上扫描,再用软件Image J计算叶面积(cm2)。扫描测定完的叶片装入信封袋,放入60℃烘箱内烘48 h至恒重,用电子天平称取叶片干重。将烘干的叶片磨成粉状,过0.25 mm筛,保存备用。分析前在65℃的干燥箱中烘干1 h,放入干燥器中保存待测。利用元素分析仪(Costech ECS 4010,意大利)测定叶片氮含量。
性状指标包括:植株高度、叶面积、比叶面积、叶干物质含量和叶氮含量。其中比叶面积和叶干物质含量的计算公式分别为:比叶面积(m2·kg-1)=叶面积/叶干重;叶干物质含量(g·g-1)=叶干重/叶饱和鲜重[22]。
重要值=(相对高度+相对盖度+相对生物量)/3,式中:相对高度为某一物种的高度占样方内全部物种高度之和的百分比;相对盖度为某一物种的盖度占样方内全部物种盖度之和的百分比;相对生物量为某一物种的生物量占样方内全部物种生物量之和的百分比。
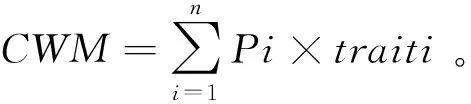
式中Pi是物种i在群落内的相对生物量,即物种i的生物量与群落中全部物种生物量和的比值;traiti是物种i的性状值。
1.4 数据分析
双因素方差分析比较放牧与氮添加及二者交互作用对糙隐子草和群落功能性状的影响,单因素方差分析比较糙隐子草和群落水平功能性状在不同处理间的差异,多重比较采用最小显著差异法(LSD)。采用回归分析法探究糙隐子草功能性状与群落功能性状之间的关系。数据统计分析与制图分别利用SPSS 20.0与Sigmaplot 10.0软件进行。文中数据均为平均值±标准误。
2 结果与分析
2.1 放牧与短期氮添加处理下植物重要值变化
调查发现,试验小区以一年生草本为主,多年生植物所占比例较少。另外,放牧与氮添加处理对植物重要值有明显影响。围封草地在不施氮的情况下优势种为糙隐子草、砂蓝刺头和猪毛蒿,其重要值分别为22.44%,21.26%和20.06%;氮添加处理下优势种为糙隐子草、白草和猪毛蒿,其重要值分别为25.37%,23.02%和19.45%。放牧草地在不施氮的情况下优势种为砂蓝刺头、糙隐子草和白草,其重要值分别为23.19%,22.48%和14.36%;氮添加处理下优势种为糙隐子草、猪毛蒿和白草,其重要值分别为26.25%,17.20%和15.99%(表1)。可以看出,糙隐子草在不同处理下都具有较高的重要值,其中,同一氮水平下放牧增加了糙隐子草重要值,氮添加增加了围封和放牧草地糙隐子草的重要值。同时,不论是围封区还是放牧区,氮添加使共优种一年生草本砂蓝刺头的重要值降低,使多年生植物白草重要值有所增加。
2.2 放牧与短期氮添加对糙隐子草功能性状的影响
方差分析表明,放牧对糙隐子草叶面积有极显著影响(P<0.01),氮添加对其叶干物质含量和叶氮含量有显著影响(P<0.05),两者交互作用对其性状均无显著影响(表2)。具体来看,氮添加使糙隐子草高度有不同程度的增加,放牧使其高度降低,但未达到显著水平;FN+处理下叶面积显著高于GN0和GN+处理,而FN+处理下的叶干物质含量显著低于FN0处理(P<0.05);就叶氮含量而言,不论是围封区还是放牧区,氮添加均使其显著增加(P<0.05)(图1)。说明糙隐子草主要通过改变叶面积来响应放牧干扰,通过改变叶干物质含量和叶氮含量来响应氮沉降。
2.3 放牧与短期氮添加对群落功能性状的影响
方差分析表明,放牧对半干旱沙质草地植物群落高度有显著影响(P<0.05),氮添加对叶面积、比叶面积和叶氮含量有极显著影响(P<0.01),放牧和氮添加两者交互处理对叶干物质含量有显著影响(P<0.05)(表3)。具体来看,放牧(GN0和GN+)显著降低了植物群落高度(P<0.05),而氮添加对其影响不显著;FN+处理下叶面积显著高于只进行围封或放牧处理(P<0.05);比叶面积在氮添加(FN+和GN+)处理下显著高于只进行放牧处理(P<0.05),而氮添加使得放牧区植物群落叶干物质含量显著降低(P<0.05);就叶氮含量而言,放牧会使其降低,但未达到显著水平,而氮添加(FN+和GN+)处理显著增加了围封和放牧区植物群叶氮含量(P<0.05)(图2)。说明群落水平上植物通过改变高度来响应放牧干扰,通过叶面积、比叶面积及叶氮含量变化来响应氮沉降。

表1 不同处理下半干旱沙质草地植被重要值的变化Table 1 Changes of important values of vegetation in semi-arid sandy grassland under different treatment
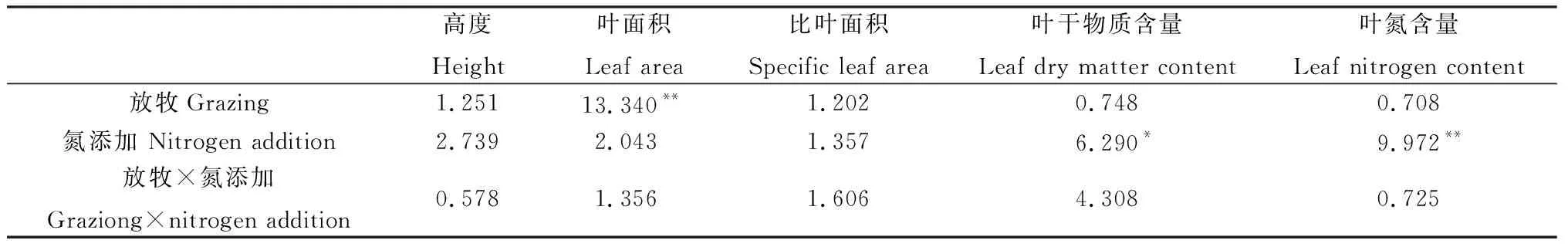
表2 放牧、氮添加及其交互作用对糙隐子草功能性状的影响(F值)Table 2 Effects of grazing,nitrogen addition and its interaction on functional traits of Cleistogenes squarrosa (F value)


图1 不同处理间糙隐子草功能性状的变化Fig.1 Changes of functional traits of Cleistogenes squarrosa under different treatments注:FN0:围封不施氮;GN0:放牧不施氮;FN+:围封并施氮;GN+:放牧并施氮。不同小写字母表示差异显著(P<0.05),下同Note:F:fenced grassland with no nitrogen;G:grazing grassland with no nitrogen;FN+:fenced and nitrogen addition;GN+:grazing and nitrogen addition. Different lowercase letters indicate significant differences at the 0.05 level,the same as below
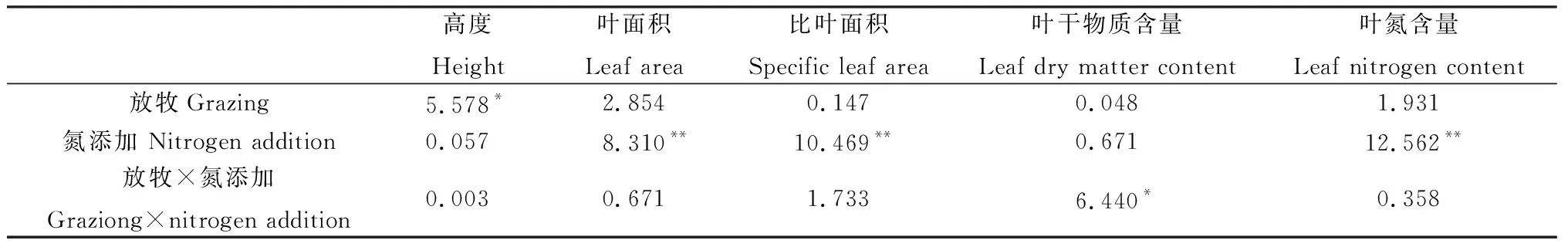
表3 放牧、氮添加及其交互作用对群落功能性状的影响(F值)Table 3 Effects of grazing,nitrogen addition and its interaction on community functional traits (F value)
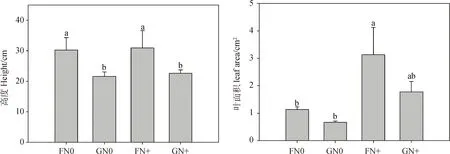

图2 不同处理间群落功能性状的变化Fig.2 Changes of community functional traits under different treatments
2.4 糙隐子草功能性状与植物群落功能性状间的关系
回归分析表明,糙隐子草高度、叶面积、比叶面积、叶干物质含量及叶氮含量与群落相对应的功能性状间存在不同程度的正相关关系(图3)。其中,叶面积和叶氮含量在两者间存在显著正相关关系(分别为:R2=0.39,P<0.01;R2=0.24,P<0.05),而高度、比叶面积和叶干物质含量在两者间相关性不显著。说明糙隐子草的叶面积和叶氮含量在一定程度上能解释群落水平叶面积和叶氮含量的变化。


图3 糙隐子草功能性状与群落功能性状间的关系Fig.3 Relationship between functional traits of Cleistogenes squarrosa and community functional traits
3 讨论
3.1 放牧和短期氮添加对植物优势度的影响
重要值反映了物种在群落中的优势地位,重要值越大说明物种在群落中的地位越高,反之则越低[24]。本研究中放牧增加了糙隐子草的重要值,氮添加增加了围封和放牧草地糙隐子草的重要值,说明放牧和氮添加在一定程度上可以促进糙隐子草的生长,进一步验证了糙隐子草的超补偿性生长[15]。另外,围封样地在不进行施氮处理下以糙隐子草、砂蓝刺头和猪毛蒿为主,添加氮素后砂蓝刺头的重要值降低,白草重要值明显增加,说明在排除放牧干扰后白草获取氮素的能力可能强于砂蓝刺头。因此,长期氮添加可能会影响植物群落优势种的地位。
3.2 优势种糙隐子草和植物群落功能性状对放牧和短期氮添加的响应
放牧对植物生长会产生一定的影响,而植物本身也会通过调节其外部形态及生理特征产生避牧或耐牧机制,从而适应环境变化[25]。有学者认为,植株高度是植物对放牧作出响应最直观的性状[16]。本研究发现,放牧显著降低了植物群落高度,符合前人提出的矮小化型变理论[26],而放牧对糙隐子草高度无明显影响,这可能是由于糙隐子草自身高度较矮,且木质素和纤维素成分相对较高,适口性较差,不利于反刍动物的消化代谢,进而减少了家畜对其进行采食。Zheng等[27]发现,位于群落下层的糙隐子草比上层的羊草(Leymuschinensis)具有更强的避牧性和耐牧性,本研究结果与其一致。氮添加可以促进养分受限环境中植株高度的增加,以增强对光资源的竞争能力[28],但本研究中短期氮添加对群落高度无显著影响,这可能与养分在植物不同构件中的分配差异有关[29]。增加植物高度有利于光资源的获取,但同时伴随着较高的成本,如投资和维持茎等支撑结构而产生成本,以及水分输送到较高部位而产生的成本[30],因此植物可能会优先将养分分配到叶片中,同样可以达到提高光合竞争能力的效果。
叶面积是可塑性最强、受载畜率影响缩减最为严重的性状,比叶面积和叶干物质含量与植物生长与生存对策有紧密联系,是植物比较生态学研究中的首选指标[31]。本研究结果表明,相同氮添加处理下放牧降低了糙隐子草叶面积,说明糙隐子草主要通过减小叶片大小来响应放牧干扰,而群落叶面积无显著变化,可能是由于群落植物间存在补偿效应[32]。此外,本研究发现只进行放牧处理对糙隐子草和植物群落比叶面积和叶干物质含量均无显著影响,张晶等[17]通过研究科尔沁沙地4种草地类型中的18种植物发现,比叶面积和叶干物质含量不受放牧干扰,本研究结果与此基本一致。氮添加显著增加了放牧区植物群落比叶面积,降低了放牧区群落叶干物质含量,说明适度放牧和短期氮添加交互作用可以通过改变与资源获取相关的性状来提高半干旱沙质草地植物快速获取资源的能力,促使植物可以在不利环境中快速完成资源的累积[9,33]。
氮素是限制陆地生态系统植物生长、维持和繁殖不可或缺的养分,且与植物光合作用密切相关[34-35]。在放牧草地,氮添加可以补偿因采食造成的光合面积下降,植物也可以通过调节资源分配策略将新产生的光合产物优先分配给光合组织,引起叶性状的相应变化,进而提高植物叶性状对家畜干扰后的恢复能力[36]。本研究中,氮添加使围封和放牧区糙隐子草和群落叶氮含量都显著增加,说明不论围封草地还是放牧草地,氮添加均能缓解土壤氮素对植物生长的限制,提高植物资源竞争能力及光合利用效率,同时较高的比叶面积也可以提高植物的氮素利用效率[37]。大部分研究发现,比叶面积和叶氮含量较高以及叶干物质含量较低的植物具有较高的养分周转率,通常表现出快速投资-收益型策略,本研究中植物性状在群落水平表现出相似的变化,可能与本研究区以短命的一年生草本为主有关。
3.3 优势种糙隐子草功能性状与群落功能性状间的相关性
群落中的优势种可以很好地代表植物群落,也可以将观察到的个体属性很好地放大到群落水平甚至整个生态系统水平[36]。从糙隐子草与群落功能性状间的相关性来看,糙隐子草叶面积和叶氮含量对群落叶面积和叶氮含量有显著影响,高度、比叶面积和叶干物质含量对群落性状影响较小。岳喜元等[38]发现典型草原优势种羊草对群落水平植物的叶面积、叶干物质含量和叶氮含量有明显影响,本研究结果与此结果具有一定的差异,可能是由于研究对象自身的形态与光合途径差异造成的[39]。羊草为一年生C3植物,处于所在群落的上层,对光资源有更高的竞争能力,而糙隐子草为多年生C4禾草,处于群落下层,在一定程度上会受到上层植被的遮光,且糙隐子草通常采取保守型资源利用策略[27],因此与资源竞争能力密切相关的性状,如高度、比叶面积和叶干物质含量,对群落性状影响不显著。此外,半干旱沙质草地植物群落以一年生植物为主,而糙隐子草为多年生植物,因此猜测半干旱沙质草地植物群落中一年生植物的功能性状,尤其是与资源获取、利用和植物光合竞争能力相关的性状,对群落水平性状变化的解释度更高,但这需要今后进一步验证。
4 结论
放牧和短期氮添加导致群落内植物优势度发生变化,放牧增加了糙隐子草的重要值,氮添加使糙隐子草和白草重要值有所增加。种群水平上,放牧显著降低了糙隐子草叶面积,氮添加增加了叶氮含量,而降低了围封草地糙隐子草叶干物质含量。群落水平上,氮添加显著增加了围封区植物叶面积和叶片氮含量,而放牧样地中,植物群落主要通过增加比叶面积和叶片氮含量、降低叶干物质含量来响应氮沉降。糙隐子草叶面积和叶氮含量能较好地解释群落水平植物叶面积和叶氮含量的变化。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
幼儿100(2020年31期)2020-11-18
河南科学(2020年3期)2020-06-02
新疆农业科学(2020年1期)2020-02-14
疯狂英语·初中版(2019年4期)2019-09-10
小太阳画报(2018年6期)2018-05-14
中国麻业科学(2015年5期)2015-12-28
草业科学(2015年9期)2015-12-11
