
血耳及其宿主菌的分离与系统发育分析
2021-04-01 07:58何贝贝陈启武
浙江农业学报 2021年3期
陈 莎,何贝贝,陈启武,李 利
(长江大学 生命科学学院,湖北 荆州 434025)
血耳,是湖北省西北部山区具有较长栽培历史的一种大型经济真菌,具有较重要的食疗价值和药用价值,可用于治疗各种妇科疾病、痢疾和急性肝炎[1]。1990年彭寅斌根据血耳的形态特征对其分类地位进行了研究,将其命名为TremellasanguineaY.B. Peng,归属于银耳纲(Tremellomycetes),银耳目(Tremellales),银耳科(Tremellaceae),银耳属(Tremella)[2],该分类地位一直沿用至今。
据《中国真菌志·第二卷·银耳目和花耳目》(1992)记载[3],银耳属在我国分布有32种,其中已进行人工栽培的有银耳(T.fuciformis)、金耳(T.aurantialba)和血耳,且三者均需要伴生菌才能实现子实体的发育[1,4]。研究表明,银耳和金耳与其伴生菌是一种寄生关系,银耳和金耳为寄生菌,其常见的伴生菌实质为宿主菌[4]。宿主菌一般其具有较高的纤维素酶、半纤维素酶和多酚氧化酶活性,能够分解木材,为寄生菌的生长发育提供营养和水分[4-6]。目前,银耳的宿主菌已被鉴定为阴环炭团菌(Annulohypoxylonstygium),金耳的宿主菌已被鉴定为粗毛硬革菌(Stereumhirsutum)[6-7],而血耳宿主菌的分类地位目前还未见报道。
银耳属的分类长时间内处于形态学研究阶段,包括子实体、担子、担孢子、菌丝等方面的形态特征。近年来,银耳纲的分子系统发育得到了深入研究[8-9],发现基于传统分类的银耳属在分子系统发育分析中是多源的(polyphyletic),如金耳(T.aurantialba)与Naemateliaceae科的Naematelia属聚为一类,而与银耳科,银耳属的银耳(T.fuciformis)遗传距离较远,金耳也因此被建议更名为Naemateliaaurantialba[9]。目前还未见血耳分子系统发育的研究报道。本研究利用担孢子弹射法和耳木组织培养法从椴木栽培的血耳中分离得到酵母状菌株和丝状真菌菌株,利用ITS序列对其进行了系统发育分析,旨在明确血耳及其宿主菌的分类地位,为后续血耳与其宿主菌的作用机理分析、新品种的选育提供参考。
1 材料与方法
1.1 材料
1.1.1 血耳子实体
血耳子实体采自湖北省保康县马良镇水田村,为新鲜的、成熟的椴木生血耳子实体,朵大,无病虫害。
1.1.2 培养基
马铃薯葡萄糖琼脂(PDA)培养基:马铃薯200 g,葡萄糖20 g,琼脂15 g,水1 L,pH自然。根据需要在使用前加入卡那霉素和氨苄青霉素,使其终浓度均为50 μg·mL-1。
1.1.3 仪器设备
奥林巴斯BX63正置显微镜、博迅SPX-150B-Z生化培养箱、精骐IS-RDS3恒温摇床、伯乐C1000 PCR仪、梅特勒ME204E电子分析天平、HVA-50灭菌器。
1.2 方法
1.2.1 菌株的分离纯化
采用担孢子弹射法和组织培养法。担孢子弹射法:用无菌水漂洗血耳子实体3次,再用无菌纱布把血耳表面的水分吸干,切一小块耳片,用无菌的不锈钢小钩钩住并悬挂于装有PDA(含50 μg·mL-1卡那霉素和50 μg·mL-1氨苄青霉素)的三角瓶内,塞上棉塞,25 ℃放置24 h,取出耳片,置28 ℃培养3 d后, 挑取白色的酵母状菌落在PDA培养基上划线分离得到纯培养。耳木组织培养法[10]:取耳根基部的耳木组织一小块,用无菌水漂洗3次,再用无菌纱布吸干表面水分,置于PDA(含50 μg·mL-1卡那霉素和50 μg·mL-1氨苄青霉素)平板中28 ℃培养7 d,挑取绒毛状菌落边缘的菌丝,转接至新的PDA平板,28 ℃培养,反复多次转接菌落边缘菌丝进行纯化。
1.2.2 菌落形态和显微形态观察
取新鲜的酵母状担孢子培养物在PDA平板中划线,28 ℃培养3 d;取新鲜的丝状真菌点接于PDA平板中央,28 ℃培养7~14 d,观察菌落颜色、形状、大小、边缘、表面等特征以及显微形态特征。
1.2.3 基因组DNA的提取
将分离到的菌株接种到50 mL 马铃薯葡萄糖液体培养基(PDB)中,28 ℃,180 r·min-1培养。酵母状菌株的培养时间为2 d,丝状真菌的培养时间为5 d。无菌滤纸过滤培养液收集菌体细胞,用无菌吸水纸压干水分,用CTAB法提取基因组DNA[11]。
1.2.4 ITS序列的扩增、测序与系统发育分析
ITS序列的扩增引物为通用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)[12-13]。20 μL的PCR反应体系含有2×pfu Master Mix 10 μL,ITS1F和ITS4各10 pmol·L-1,DNA模板50 ng。PCR反应程序为:94 ℃预变性5 min,94 ℃变性30 s,56 ℃退火30 s,72 ℃延伸1 min,30个循环;72 ℃后延伸10 min,4 ℃保存。所扩增的PCR产物经琼脂糖凝胶电泳检测后送往生工生物工程(上海)股份有限公司测序。测序得到的酵母状菌株(XE01)序列与16条来源于NCBI(https://www.ncbi.nih.gov)核酸数据库中的ITS序列构建系统发育树,包括 10条褐银耳属(Phaeotremella)菌株序列、5条银耳属(Tremella)菌株序列和1条胶珊瑚属(Holtermannia)菌株序列(表1),其中胶珊瑚属(Holtermannia)菌株作为外群(outgroup);测序得到的丝状真菌菌株(XE02)序列与8条来源于NCBI核酸数据库中的ITS序列构建系统发育树,包括5条韧革菌属(Stereum)菌株序列、1条阴环炭团菌(Annulohypoxylonstygium)菌株序列、1条香灰菌菌株序列和1条根霉属(Rhizopus)菌株序列(表1),其中根霉属菌株作为外群。采用Clustal X 2.1[14]软件进行序列比对;采用MEGA X软件[15]进行人工校正,删除两端未对齐的冗余序列后,以N-J(neighbor-joining)法构建系统发育树。
2 结果与分析
2.1 血耳子实体的形态特征
血耳子实体丛生,常生长成一大团,半球形,直径8~20 cm(图1)。耳片相互挤在一起,底部相连,上部分开呈瓣状;单个耳片呈波浪状卷曲,肉质薄,宽2~5 cm,厚0.5~1.0 mm。新鲜子实体柔软,硬胶质,富有弹性,颜色为暗赤褐色;干制后呈黑色。
2.2 两株分离菌株的形态特征
采用担孢子弹射法获得一株酵母状菌株,命名为XE01。菌株XE01在PDA培养基中28 ℃培养,形成白色圆形菌落,3 d时菌落直径约为1~2 mm,不透明,表面凸起,光滑,湿润(图2-a)。显微观察发现细胞为椭圆形,大小为(1~2)μm×(2~3)μm,部分细胞带有芽体(图2-b)。

表1 用于构建系统发育树的ITS序列信息

图1 血耳的新鲜子实体Fig.1 The fresh basidiocarps of Tremella sanguinea
采用耳木组织培养法获得一株丝状真菌,命名为XE02。菌株XE02在PDA培养基中28 ℃培养,形成白色绒毛状菌落,7 d时菌落直径约为6.3 cm,培养基中可见红褐色分泌物,随着培养时间延长,红褐色分泌物更明显(图3-a);显微观察发现菌丝分枝少,有隔,宽1~3 μm,菌丝顶端可见单生的球状孢子(图3-b),可见锁状联合(图3-c)。
2.3 两株分离菌株的系统发育分析
菌株XE01和XE02的ITS扩增片段经测序,分别得到长为551 bp和624 bp的DNA序列(GenBank 登录号分别为MN894289和MN894290)。BLAST分析发现,菌株XE01与褐银耳属的PhaeotremellaeugeniaeLE303429、P.foliaceaDSM 5493等菌株的ITS序列同源性较高,序列一致性分别为99.5%和99.1%;菌株XE02与韧革菌属的S.gausapatumCBS349.39、S.sanguinolentumCBS529.50等菌株的ITS序列同源性较高,序列一致性分别为95.4%和95.5%。系统发育分析表明,菌株XE01与褐银耳属(Phaeotremella)聚为一支,与P.eugeniae和P.neofoliacea的遗传距离最小,亲缘关系最近(图4);菌株XE02与韧革菌属(Stereum)聚为一支,与皱韧革菌(S.rugosum)的遗传距离最小,亲缘关系最近(图5)。
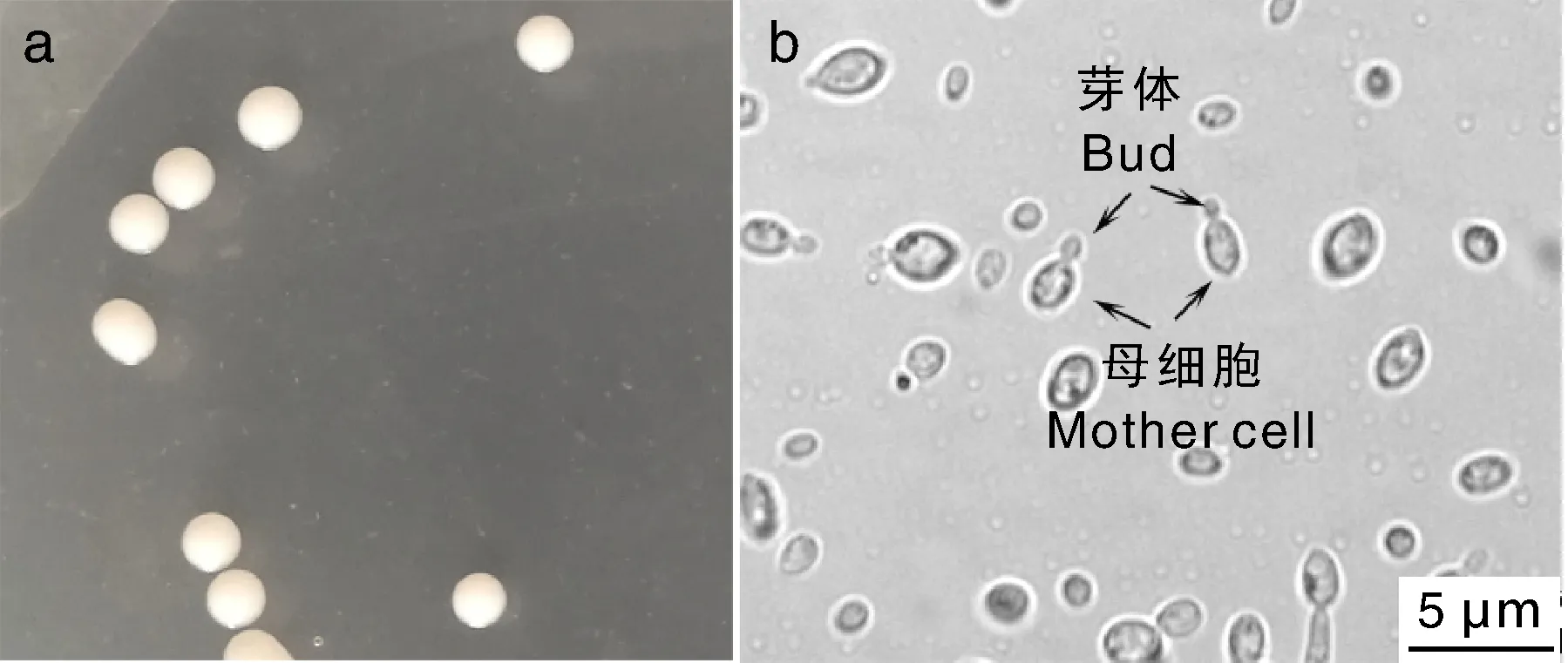
图2 XE01菌株菌落及显微形态Fig.2 Colony and microscopic morphology of the strain XE01
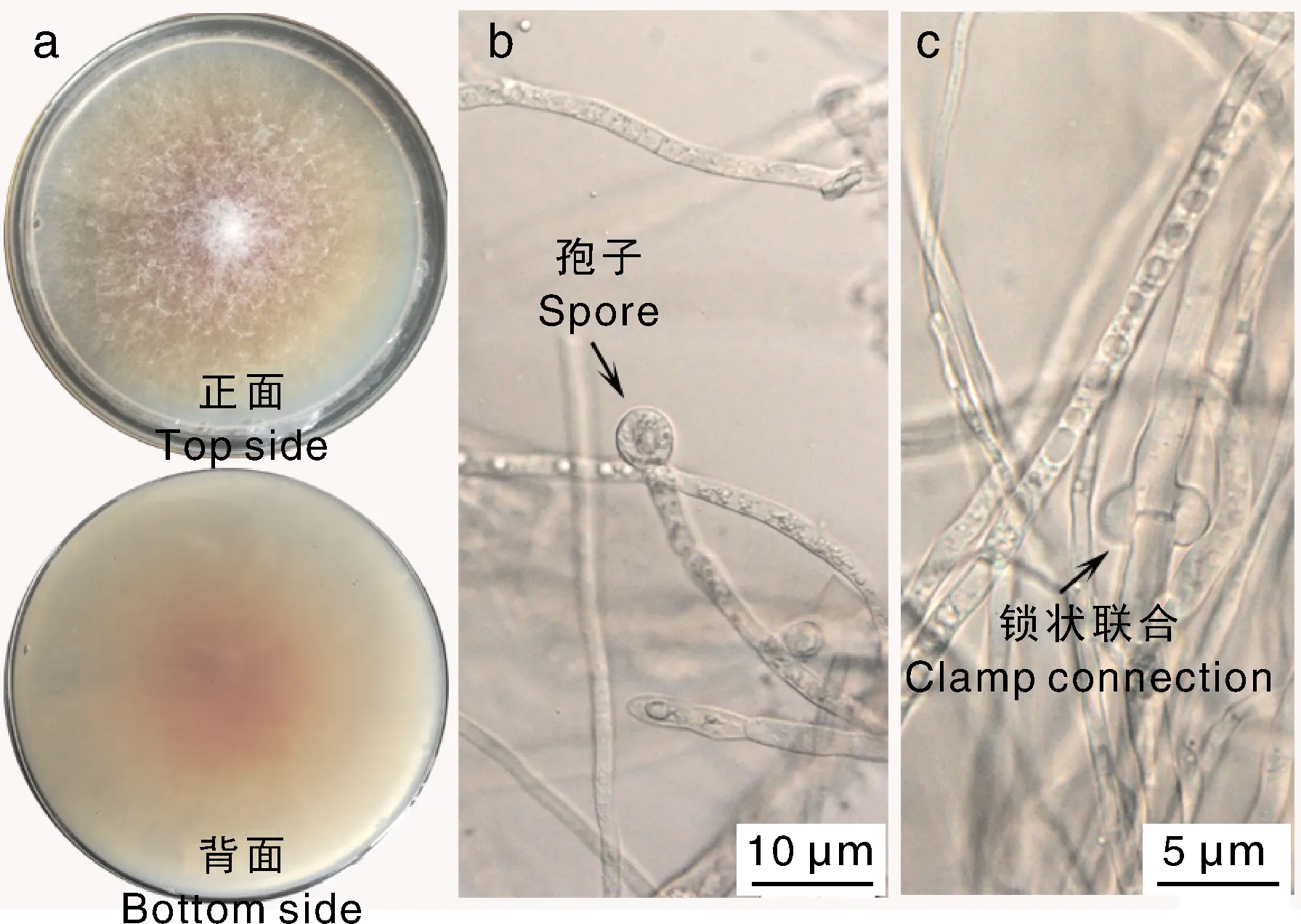
图3 XE02菌株菌落及显微形态Fig.3 Colony and microscopic morphology of the strain XE02
3 讨论
彭寅斌[2]根据血耳子实体、菌丝等方面的形态特征将其鉴定为银耳属的一个新种。传统分类中的银耳属为有性型(teleomorphic)担子菌,通常存在二相性(dimorphic),即生活周期中存在酵母型和菌丝型两种形态间发生互变[8-9]。随着银耳纲真菌的分子系统发育研究,人们发现银耳属担子菌具有明显的多源性(polyphyletic),分布于多个亲缘关系较远的支系中[8-9,24-25]。Liu等[9]根据7个基因的系统发育分析,将传统分类中银耳属重新归类,该属只保留了Tremellasensu stricto支系的11个种:T.brasiliensis、T.cinnabarina、T.coalescens、T.flava、T.fuciformis、T.globispora、T.laurisilvae、T.mesenterica、T.resupinata、T.taiwanensis、T.tropica,并且将Cryptococcusyokohamensis归为银耳属;同时,银耳属的其他种被归到银耳纲的其他类群中,其中银耳属的5个种归为银耳目(Tremellales),褐银耳科(Phaeotremellaceae)的褐银耳属(Phaeotremella),分别为P.foliacea、P.simplex、P.mycetophiloides、P.mycophaga、P.neofoliacea。褐银耳属担子菌的典型性状包括:子实体叶状,丛生,胶质,新鲜时棕褐色到肉桂色或浅棕色,干燥后深棕色到黑色;菌丝有锁状联合,偶尔可见吸器枝(haustorial branches);担子球形、近球形、卵球形或椭圆形,四孢,具纵向到斜向或横向隔膜;担孢子近球形到宽椭圆形,在培养过程中,可见芽体[9]。彭寅斌[2]描述的血耳典型性状及本研究的实验结果(图1和图2),符合Liu等[9]描述的褐银耳属特征,血耳分类地位的合理性因此而受到质疑。血耳的分子系统发育分析将为阐明血耳的分类地位提供分子证据。

图4 基于rDNA-ITS序列的菌株XE01系统发育树Fig.4 The phylogenetic tree of the strain XE01 inferred from rDNA-ITS
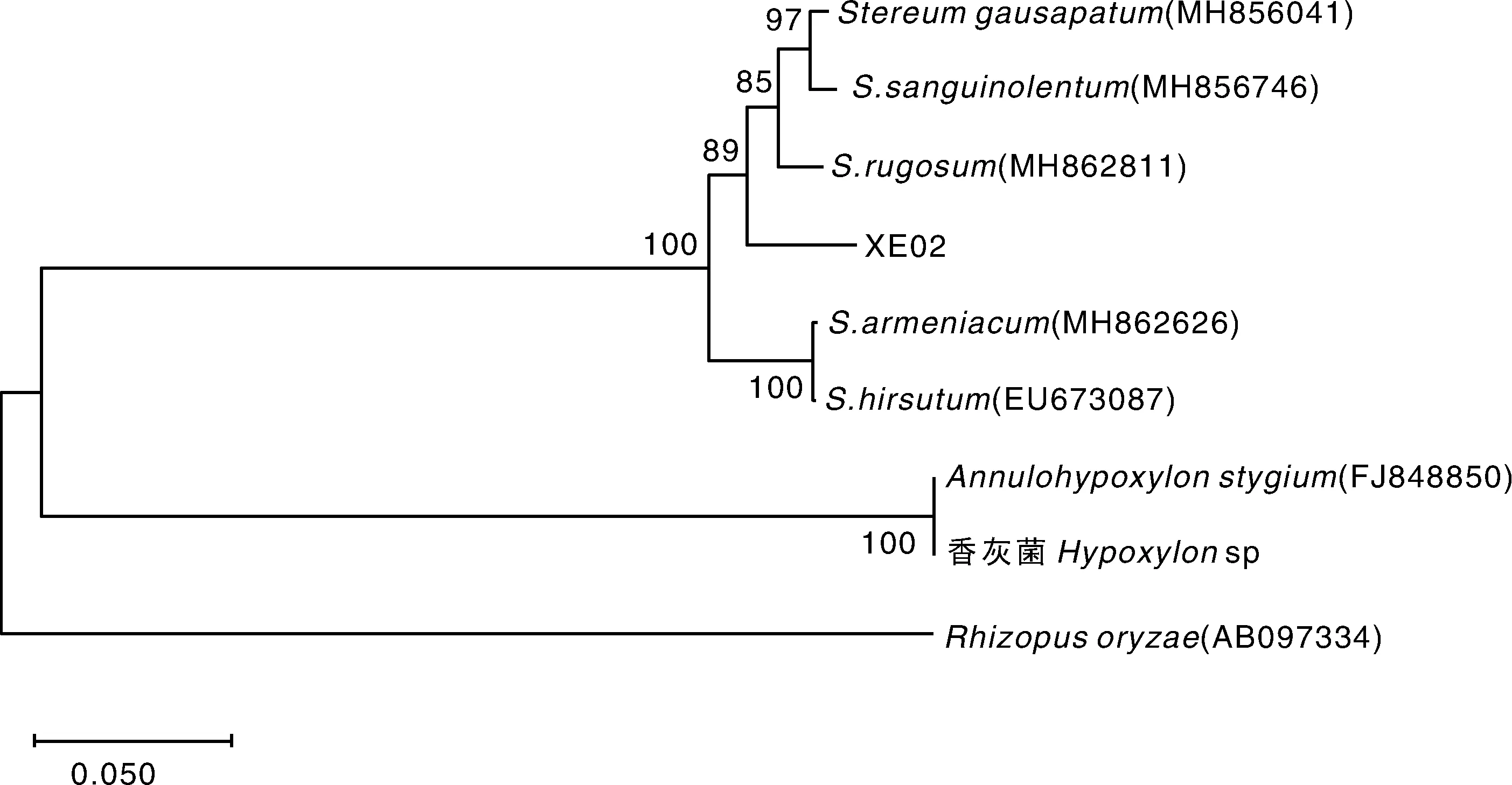
图5 基于rDNA-ITS序列的菌株XE02系统发育树Fig.5 The phylogenetic tree of the strain XE02 inferred from rDNA-ITS
本研究以新鲜的血耳子实体为材料,以担孢子弹射法和耳木组织培养法分别获得1株酵母状菌株XE01和1株丝状真菌菌株XE02。基于ITS的系统发育分析显示,酵母状菌株XE01跟褐银耳属亲缘关系最近,而与银耳属分布在两个不同的支系中(图4);丝状真菌菌株XE02与韧革菌属的亲缘关系最近,与已报道的银耳宿主菌香灰菌[21-22]分布于不同支系中(图5)。褐银耳属的担子菌P.foliacea、P.simplex、P.mycetophiloides、P.mycophaga、P.neofoliacea等均为二相性真菌,其担孢子的培养物为酵母状真菌,培养过程中可见芽体[9],结合担孢子弹射法所获得的酵母状菌株XE01的形态特征(图2)和系统发育分析结果(图4),推测酵母状菌株XE01为血耳担孢子的培养物,是血耳的酵母态型。韧革菌属典型的培养特征包括能形成有隔菌丝、锁装联合及厚垣孢子、能导致琼脂培养基背面发生褐色改变等[26],本研究从耳木中分离到的丝状真菌XE02的形态特征(图3)与报道的韧革菌属相符,系统发育分析结果(图5)也支持其归为韧革菌属,且与皱韧革菌(S.rugosum)亲缘关系最近。皱韧革菌(S.rugosum)是常见的褐银耳属担子菌的宿主菌[16],由此推测,丝状真菌XE02可能为血耳的宿主菌。据报道,银耳及金耳与其宿主菌的亲和性有较严格的专一性,不能随意搭配[4]。在人工栽培时,必须将银耳或金耳的菌丝与其宿主菌的菌丝按一定比例制成混合型菌种,才能形成银耳或金耳子实体。关于血耳XE01与韧革菌XE02寄生关系的进一步验证,有待在获得菌丝型血耳菌株后,进行混合制种及人工栽培试验。
Spirin等[16]基于形态、生态、地理分布和ITS、LSU和tef1序列,对广义Phaeotremellafoliacea类群中不同物种的分类进行了研究,发现该类群中存在同物异名(synonyms)的现象,认为P.neofoliacea、P.skinneri和P.foliacea为同一物种。本研究基于ITS序列的系统发育分析发现(图4),血耳XE01与P.eugeniaeLE303429、P.neofoliaceaCBS8481、P.foliaceaDSM5493、P.fimbriataNiemela 7897和P.skinneriCBS5029遗传距离非常小,是否与这些物种之间存在同物异名现象有待进一步研究。
猜你喜欢
动物医学进展(2022年6期)2022-11-26
南京师大学报(自然科学版)(2022年2期)2022-06-20
四川蚕业(2022年1期)2022-06-06
百姓生活(2021年5期)2021-07-08
海峡姐妹(2018年8期)2018-09-08
家庭医药·快乐养生(2017年6期)2017-06-16
家庭医药·快乐养生(2017年4期)2017-04-19
小星星·阅读100分(高年级)(2015年11期)2015-11-28
科学大众(中学)(2015年9期)2015-10-12
大自然探索(2015年11期)2015-09-10
