
中国海洋浮游植物和赤潮物种的生物多样性研究进展(四): 长江口*
2021-03-30 02:28陈楠生崔宗梅
海洋与湖沼 2021年2期
陈楠生 崔宗梅, 4 徐 青, 5
中国海洋浮游植物和赤潮物种的生物多样性研究进展(四): 长江口*
陈楠生1, 2, 3崔宗梅1, 2, 3, 4徐 青1, 2, 3, 5
(1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室 青岛 266071; 2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室 青岛 266237; 3. 中国科学院海洋大科学研究中心 青岛 266071; 4. 中国科学院大学 北京 100049; 5. 华中农业大学生命科学技术学院 武汉 430070)
长江口海域是我国的一个典型水域, 具有很高的浮游植物物种多样性, 其中包括很多种赤潮物种, 引起赤潮频发。本文总结了过去40年间长江口海域历次航次调查的研究结果, 比较分析了52个航次调查鉴定的浮游植物组成, 重点探索了赤潮物种的组成, 并详细描述了长江口海域的6种典型赤潮物种: 中肋骨条藻、东海原甲藻、旋链角毛藻、夜光藻、米氏凯伦藻和尖刺拟菱形藻。在文献报道的657个浮游植物物种(包括571个物种和86个鉴定到属的未定种)中, 硅藻(368个物种和55个未定种)和甲藻(149个物种和15未定种)占绝大多数。这些浮游植物中赤潮物种占125个, 包括硅藻69种、甲藻49种、金藻2种、隐藻1种和蓝藻3种。本文为比较研究不同海域赤潮物种的生物多样性, 探讨生物多样性的起源与演化, 为解读宏基因组学及宏条形码分析结果提供了基础。
长江口; 生物多样性; 航次调查; 赤潮物种; 浮游植物
长江是我国最大的河流, 也是全世界长度排名第三的河流, 平均年径流量高达9.6×1011m3。长江口(Changjiang Estuary)位于东海北部, 黄海南部, 西靠大陆, 长江径流入海。长江巨量径流和泥沙入海形成显著的冲淡水舌, 为该海域带来浮游植物生长需要的营养盐(赵保仁, 1991)。长江口沿岸是我国经济发展最为活跃的区域之一, 人类活动频繁, 使得水体中营养盐变化较大。富营养化是引起赤潮暴发的具体原因之一。除此之外, 长江口邻近海域受复杂水系的影响, 包括受长江冲淡水(Changjiang diluted water)、台湾暖流(Taiwan warm current)的直接影响, 以及受到苏北沿岸流(Subei coastal current)、浙闽沿岸流(Zhejiang and Fujian coastal current)、黄海冷水团(Huanghai cold water mass)及黑潮(Kuroshio current)的间接影响, 加上各流系存在明显的季节变化, 使得长江口的理化条件较为复杂(图1)。这些随季节变化的复杂的水流系统也促进了长江口浮游植物的多样性及动态变化(周名江等, 2003; 李照, 2018)。另外, 台湾暖流沿浙闽近海终年北上, 与长江冲淡水交汇形成了重要的羽状峰及辐聚带, 为浮游植物的生长提供合适条件的同时, 也促进了赤潮的暴发(周名江等, 2003)。
由于不同浮游植物对营养盐的吸收利用策略不同, 富营养化过程也导致藻华原因种的多样性和优势类群的变化(于仁成等, 2020)。受陆源影响, 1960—2010年间长江口海域溶解无机氮的浓度和通量呈上升趋势, 磷酸盐的入海通量也在不断增加。虽然浓度变化范围不大, 但是这些变化仍然影响了该海域浮游植物的生长。比如, 有报道认为长期氮磷比的失衡导致甲藻类在浮游植物群落中的占比升高(李云等, 2007)。相比之下, 硅酸盐的浓度和通量自1960年以来呈逐渐下降的趋势。总体来讲, 营养盐的综合变化导致浮游生物大量繁殖, 指示浮游植物生物量及初级生产力的叶绿素(chlorophyll)的浓度也呈现出逐年升高的趋势(王江涛等, 2012)。
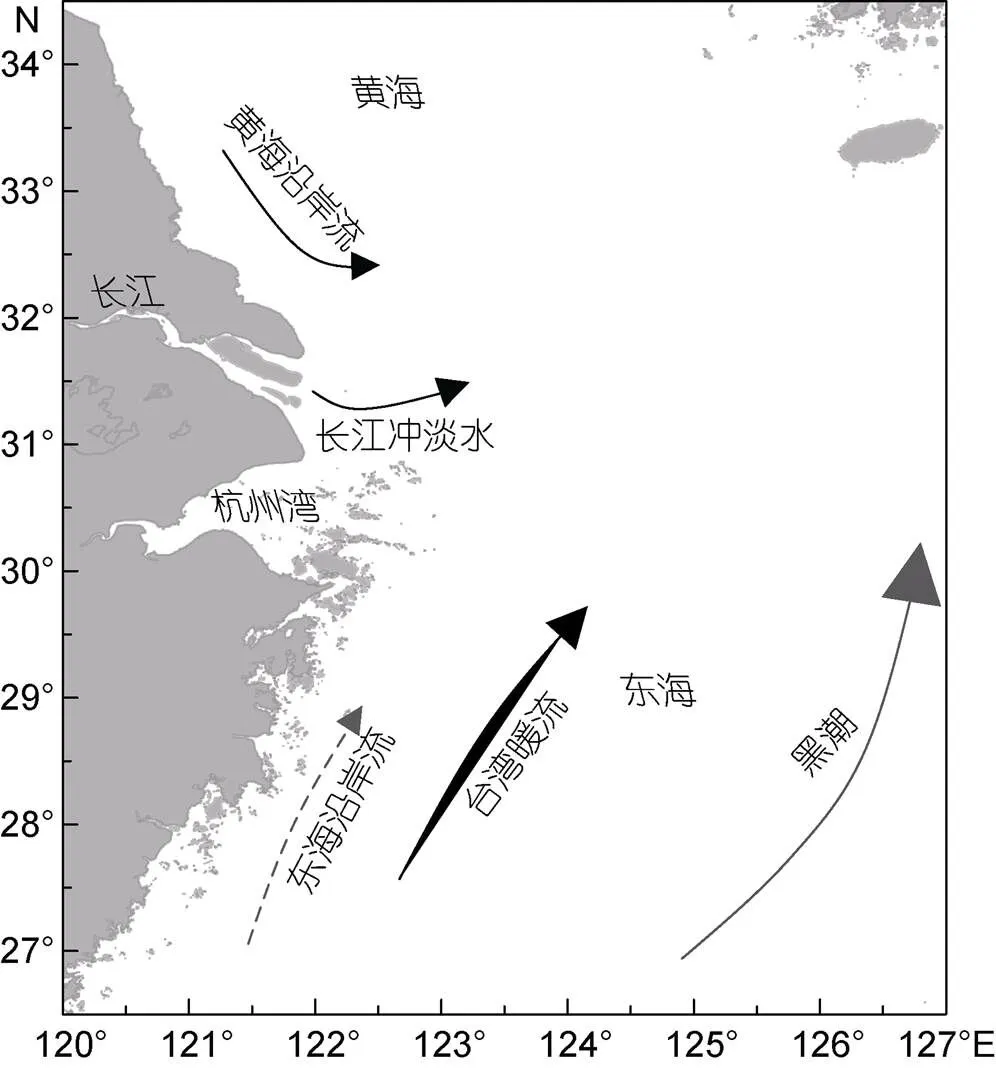
图1 长江口的地理位置以及夏季长江口海域的海流(修改自Jiang et al, 2014)
除了上述长期变化, 长江口邻近海域营养盐的浓度还存在明显的季节变化, 三种营养盐DIN、PO4-P和SiO3-Si的浓度均在每年的秋天和冬天达到高峰(Li, 2014)。叶绿素浓度在每年夏季最高, 其次是春季和秋季, 冬季最低(Yang, 2014)。
气候变化如全球变暖也会影响赤潮的发生规模、发生频率和分布情况(Hallegraeff, 2010)。在1980—2005年间, 长江口水温呈冷暖交替的趋势, 水温整体呈上升的趋势(章飞燕, 2009)。水温变化在一定程度上影响了浮游植物的演替。有研究表明甲藻的最适生长温度比硅藻高。水温的升高会导致海洋水体层化更加稳定, 更适宜具有垂直迁移能力的甲藻的生长, 这种情况对依赖于混合均匀水体的硅藻来说是不利的(李云等, 2007)。
近几十年以来, 针对长江口海域浮游植物及赤潮物种的研究快速发展, 但研究方法以形态学方法为主。本文对长江口海域赤潮发生情况和赤潮物种的生物多样性进行了综述, 为未来对长江口海域浮游植物特别是赤潮物种进行系统性的分子分析提供支持。
1 长江口海域的赤潮
有害藻华是由于海洋中微型浮游生物快速增殖、聚集, 导致生态系统结构与功能破坏的一种生态异常现象, 包括了各种藻华现象, 包括海水藻华和淡水藻华, 其中包括浮游和底栖的大型藻和微型藻(俞志明等, 2019)。长江口及其邻近海域是我国最重要的赤潮高发区之一(周名江等, 2003)。早在1933年费鸿年首次记录了发生在浙江海域的夜光藻和骨条藻赤潮(中国科学技术协会等, 2015)。随着近岸海域富营养化的加剧, 20世纪70年代以后记录的赤潮次数显著增加(于仁成等, 2017)。据统计1933—2011年间长江口及其邻近海域共发生赤潮491起, 其中硅藻赤潮111起, 甲藻赤潮258起, 其中暴发于1980s以后的赤潮记录占总数的99% (杨庶, 2013)。刘录三等(2011)统计了1972—2009年间发生在长江口及其邻近海域的面积>1000 km2的赤潮事件共25次。长江口海域的赤潮暴发具有明显的地区分布特点, 主要集中在长江口佘山附近海域、花鸟山-嵊山-枸杞山附近海域及舟山东部海域(Zhou, 2008; 刘录三等, 2011)。
长江口赤潮暴发呈现出明显的季节规律, 多发生在春夏两季, 并且5—6月份暴发频繁(Li, 2014)。长江口及邻近海域引发赤潮的生物中, 最具优势的赤潮藻类被认为是东海原甲藻(D. Lu, 2001)、中肋骨条藻()和夜光藻() (刘录三等, 2011)。在长江口赤潮中, 春季往往以硅藻(包括骨条藻、海链藻和拟菱形藻)暴发的赤潮为主, 随后以甲藻暴发的赤潮为主(于仁成等, 2017)。研究表明, 东海原甲藻还具有垂直迁移能力, 生长速度快而且能够适应长江口邻近海域独特的富营养化特征而更容易形成赤潮(李英等, 2005)。
2 长江口海域的航次调查
根据对文献的梳理发现, 1977—2016年间总共完成并报道了52个长江口航次调查。这些航次分析报道了长江口海域鉴定到的大量浮游植物, 并鉴定了长江口海域的赤潮物种(附表1)。这些航次调查时间跨度长达40年(1977—2016), 航次的采样站位覆盖面积比较大, 基本覆盖了赤潮高发区域(122°15′—123°15′E, 30°30′—32°30′N)(徐韧等, 1994), 对研究长江口海域浮游植物和赤潮物种的生物多样性具有很高的参考价值(图2)。航次调查采样的季节以夏季为主, 但是覆盖了所有四季, 因此这些航次调查对系统认识长江口海域浮游植物和赤潮物种组成和时空变化规律具有比较好的参考价值。
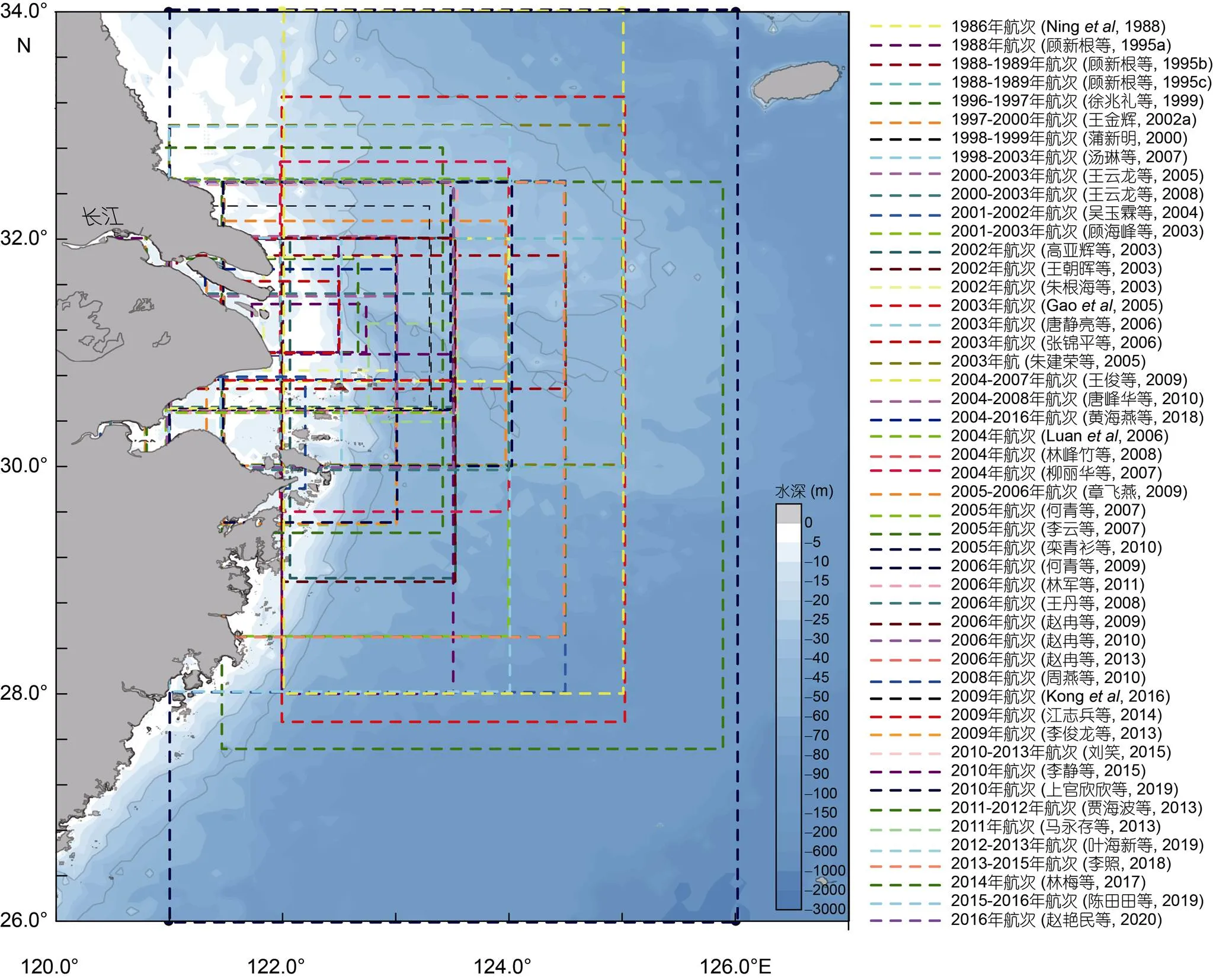
图2 长江口航次站位的分布范围
3 长江口海域的浮游植物
3.1 浮游植物的概况
综合1977—2016年间的52个长江口航次调查文献, 共鉴定并报道了571个浮游植物物种和86个鉴定到属的未定种, 包括硅藻门(Bacillariophyta)的368个物种和55个未定种, 甲藻门(Dinophyta)的149个物种和15个未定种, 绿藻门(Chlorophyta)的12个物种和5个未定种, 金藻门(Chrysophyta)的5个物种和1个未定种, 黄藻门(Xanthophyta)的2个物种和1个未定种, 隐藻门(Cryptophyta)的2个物种, 裸藻门(Euglenophyta)的2个未定种以及蓝藻门(Cyanophyta)的32个物种和7个未定种(附表2)。
值得关注的是, 在历次长江口航次调查报道的571个浮游植物物种中, 只有176个(30.8%)在NCBI数据库中有对应的全长18S rDNA序列(>1600 bp), 表明当前公共数据库中全长18S rDNA序列数据还很不全面, 长江口海域中的很多浮游植物尚未得到全面的分子鉴定和分析。针对这些物种的分子标记的解析将大大促进对长江口海域生物多样性及其动态变化规律的认识, 是我国海洋生态学者需要积极解决的问题。
利用数据库中查找到的全长18S rDNA序列构建系统进化树(图3a), 显示长江口海域的浮游植物聚类到8个浮游植物门, 与基于形态分类的结果相符。其中以硅藻门物种种类最多(112个), 其次是甲藻门(54个)。在长江口海域历次航次调查中出现频率最高的浮游植物包括中肋骨条藻、东海原甲藻、虹彩圆筛藻()、琼氏圆筛藻()、弓束圆筛藻()、洛氏角毛藻()和尖刺拟菱形藻() (图3b, 3c)。
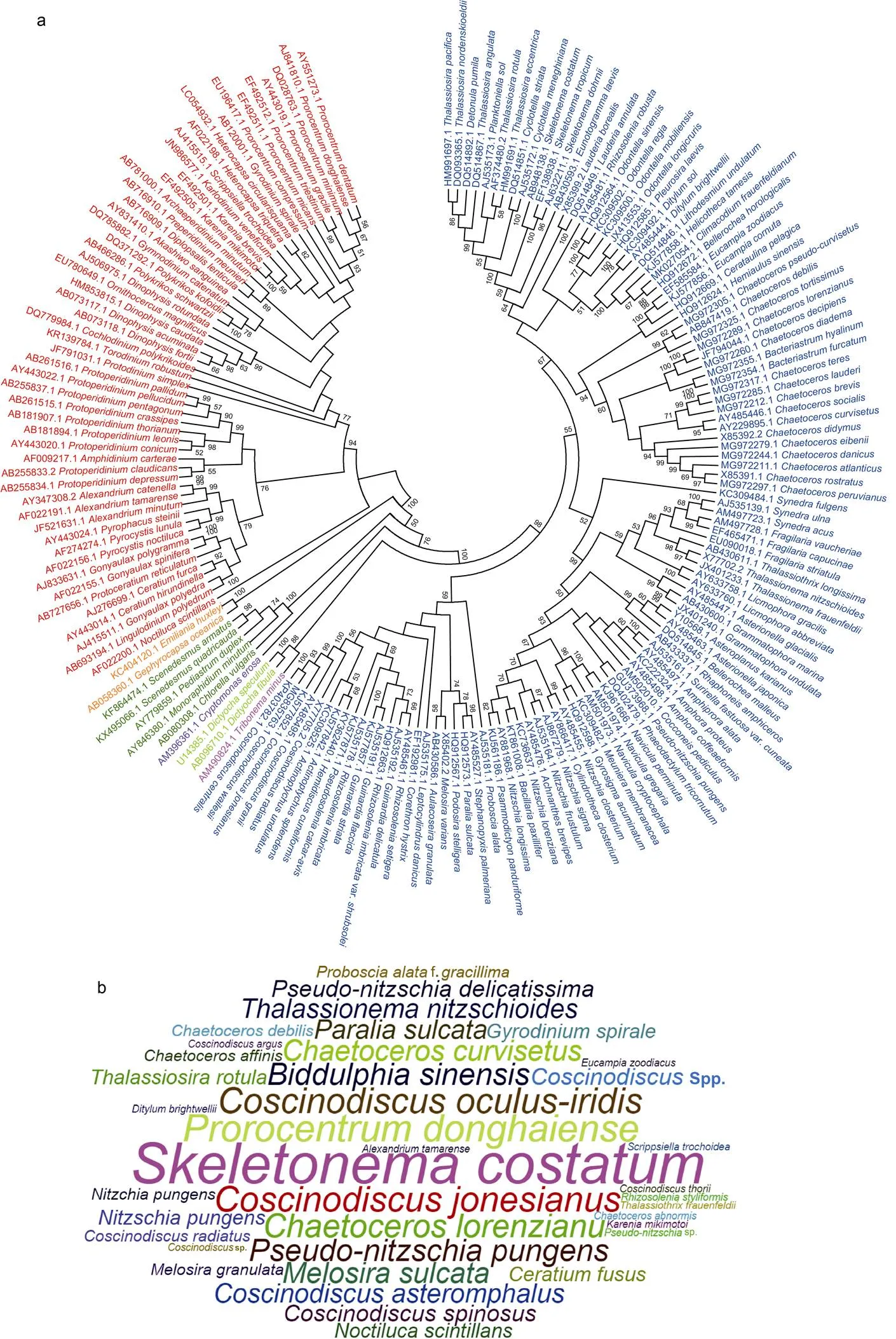
注: a. 长江口航次调查鉴定出的浮游植物的18S rDNA序列(>1600 bp)构建的系统进化树(ML), 其中蓝色代表硅藻, 红色代表甲藻; b. 长江口航次调查鉴定出的优势浮游植物(出现频次>1)词云图; c. 长江口历史文献中报道的优势浮游植物(出现频次>1)柱状图
3.2 浮游植物物种组成的比较分析
为了分析所有长江口历史航次中报道的浮游植物物种组成之间的相似性, 我们绘制了各航次浮游植物物种组成的Venn图(图4a)。比较分析可以看出, 不同航次报道的物种数目相差较大, 不同航次报道的浮游植物物种的相似性不高, 没有任何一个浮游植物物种在全部52个航次中都得到鉴别。这个统计结果表明航次调查结果受很多因素的影响, 包括采样季节、采样站位位置、采样站位数以及调查分析人员对浮游植物物种的鉴定经验(图2)。
有关长江口浮游植物类别组成变化, 多个报道认为在近几十年里, 长江口的硅藻组成逐步下降, 同时甲藻组成则逐步上升(王金辉等, 2004; 李云等, 2007; 林峰竹等, 2008; Zhou, 2008; 章飞燕, 2009; Jiang, 2010, 2014; Li, 2010, 2014; 王江涛等, 2012; Yang, 2014; 林梅等, 2017; 黄海燕等, 2018)。其中Li等(2014)整理了1959—2010年国家海洋局的历史数据, 发现1959—2000年硅藻细胞丰度在80%左右, 甲藻则低于30%, 而2000年后由于长江口氮输入的增加, 硅藻细胞丰度逐渐降低, 甲藻则逐渐升高。黄海燕等(2018)基于2004—2016年夏季长江口生态监控区13年的生态环境调查数据, 也发现在各年份物种组成上硅藻占绝对优势, 但是在占浮游植物种类数的百分比上呈下降趋势, 而甲藻则呈上升趋势。
为了验证上述假设, 评估长江口浮游植物种类的历史变迁规律, 我们系统分析了所有52个长江口调查航次中报道的甲藻和硅藻数目。比较分析表明, 硅藻组成并没有显示显著的下降趋势, 甲藻组成没有显示显著的上升趋势, 浮游植物组成中硅藻和甲藻的相对组成也没有显示出显著的变化(图4b)。文献中报道的硅藻组成逐步下降和甲藻组成逐步上升可能反映了诸多因素的影响, 比如季节、采样方法、采样位点、物种的分类标准等。除此之外, 主观原因比如人为因素(比如不同研究人员的分类能力)也可能影响了对物种的鉴定和判断。随着基于分子标记(比较宏条形码分析)等定量分析的广泛使用(陈楠生, 2020), 长江口海域浮游植物物种组成应该能够得到更加准确的判断。
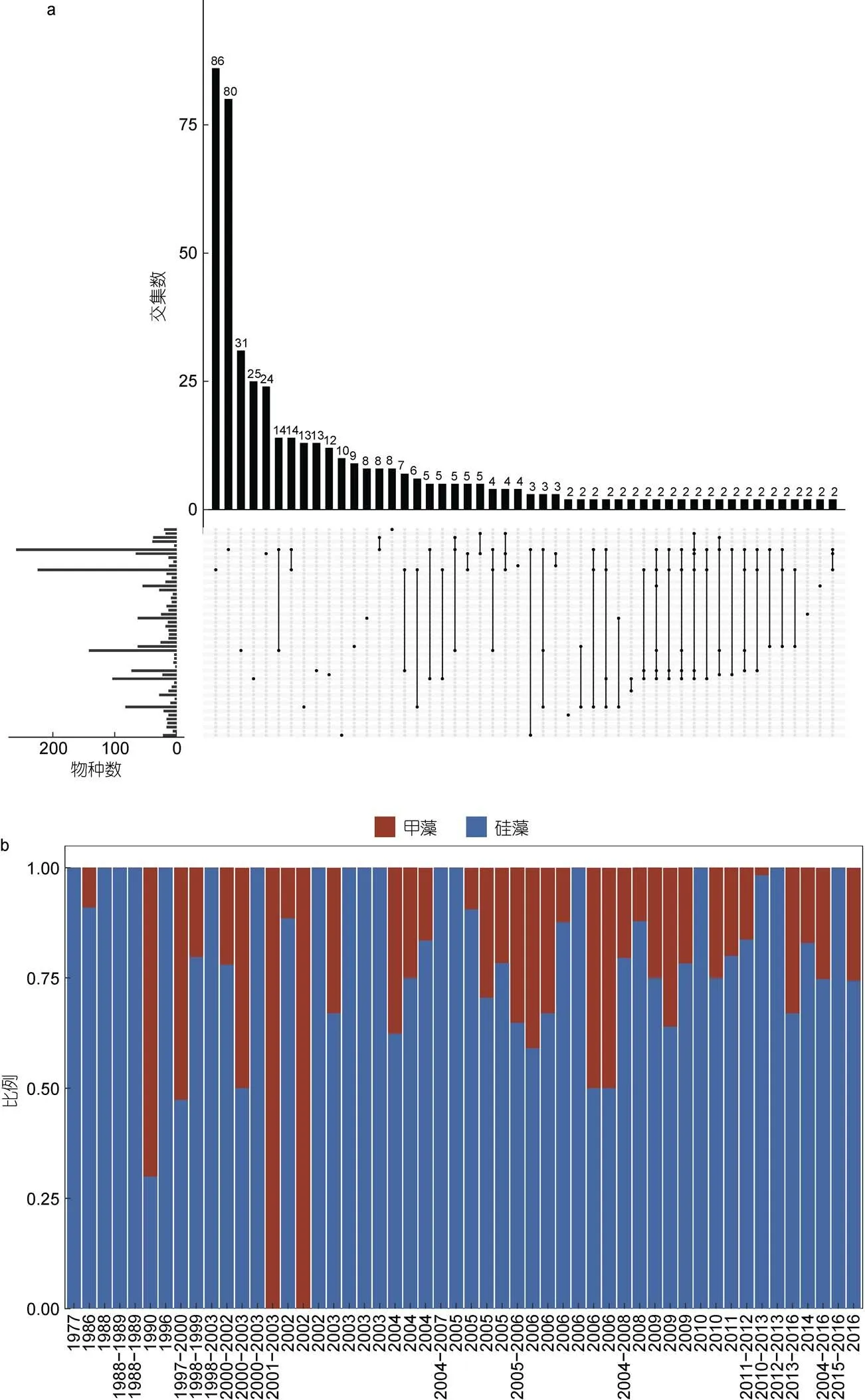
图4 长江口浮游植物组成的历史变迁
注: a. 各航次统计到的浮游植物物种, 上方的条形图显示在一个或多个航次中鉴定到的物种, 左方的条形图显示每一个航次鉴定到的所有浮游植物物种数; b. 各航次统计到的浮游植物中硅藻和甲藻的比例变化
4 长江口海域的赤潮物种
系统整理长江口及其邻近海域的52个航次的调查结果, 共发现了125个赤潮物种, 其中硅藻69种, 甲藻49种, 金藻2种, 隐藻1种, 蓝藻3种(附表3)。在52个航次中至少5个航次中检测到的赤潮物种为49个(表1)。不同航次报道的赤潮物种的种类和数目差异均较大, 并且没有一个赤潮物种在所有航次中都得到鉴定(图5a)。长江口海域中的优势赤潮物种包括中肋骨条藻、东海原甲藻、旋链角毛藻、夜光藻、米氏凯伦藻()和尖刺拟菱形藻(图5b, 5c)。
本文综合52个航次得到的125个赤潮物种数多于以前报道的数目。比如, 20世纪末长江口及其邻近海域报道了68个赤潮物种, 包括硅藻39种、甲藻26种、蓝藻2种和1种隐藻(王金辉, 2002b)。另据统计1981—2011年长江口邻近海域共有赤潮肇事种47种, 其中硅藻21种、甲藻22种(包括夜光藻)、针胞藻1种、金藻1种和定鞭藻1种(杨庶, 2013)。在过去40年中, 主要赤潮物种可能发生了变化。比如, 20世纪90年代之前该海域赤潮物种以中肋骨条藻和异养性甲藻夜光藻为主, 而在90年代后期由东海原甲藻、米氏凯伦藻、链状亚历山大藻()以及夜光藻等甲藻占优势。这种转变可能是由于高氮磷比以及长江输入氮增加导致磷酸盐限制引起的(Zhou, 2008)。
表1 长江口历史航次赤潮物种名录(出现次数前49)
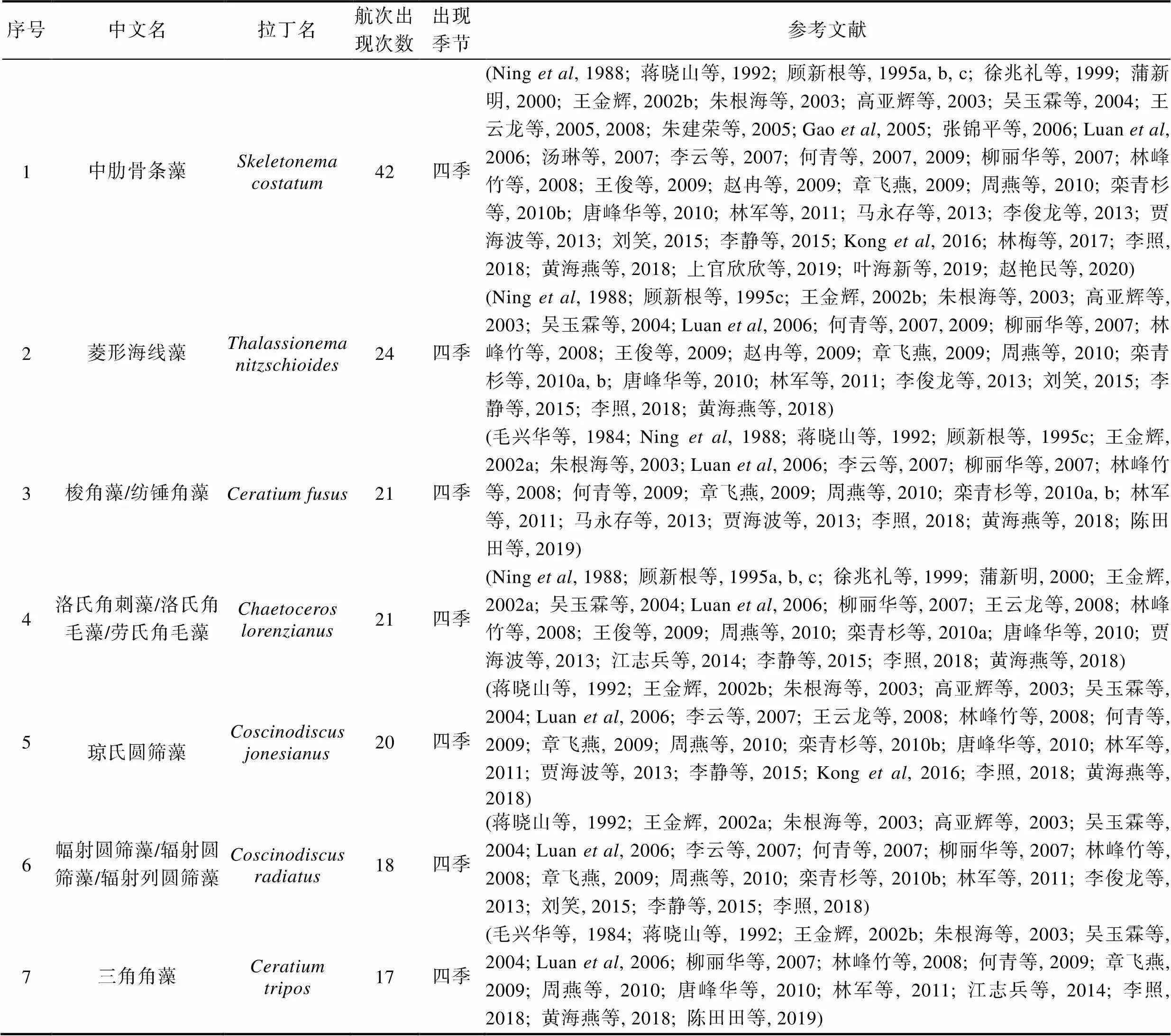
Tab.1 Red tide species identified in the expeditions over the last 40 years (Only the top 49 most abundant speciese were listed)
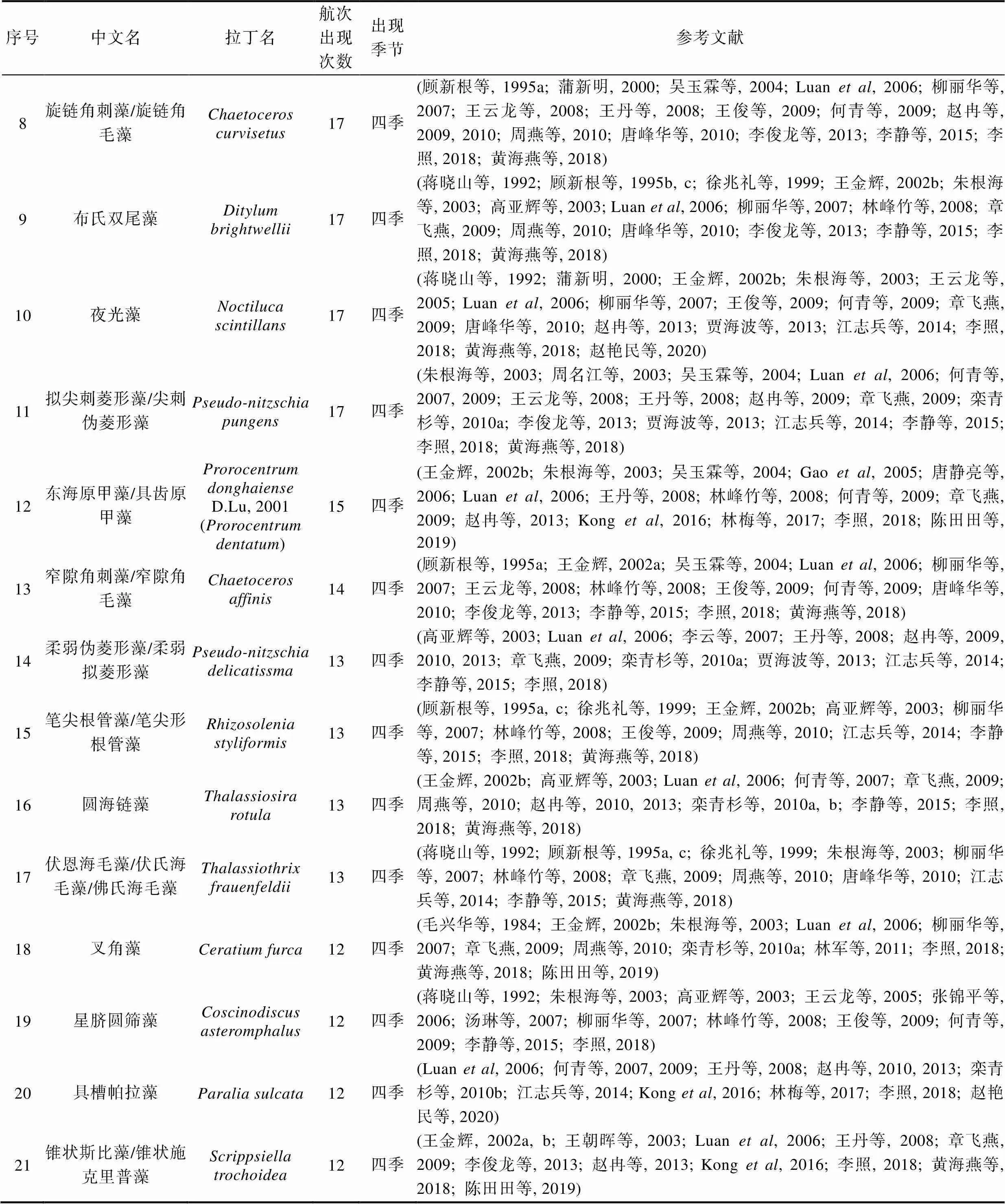
续表
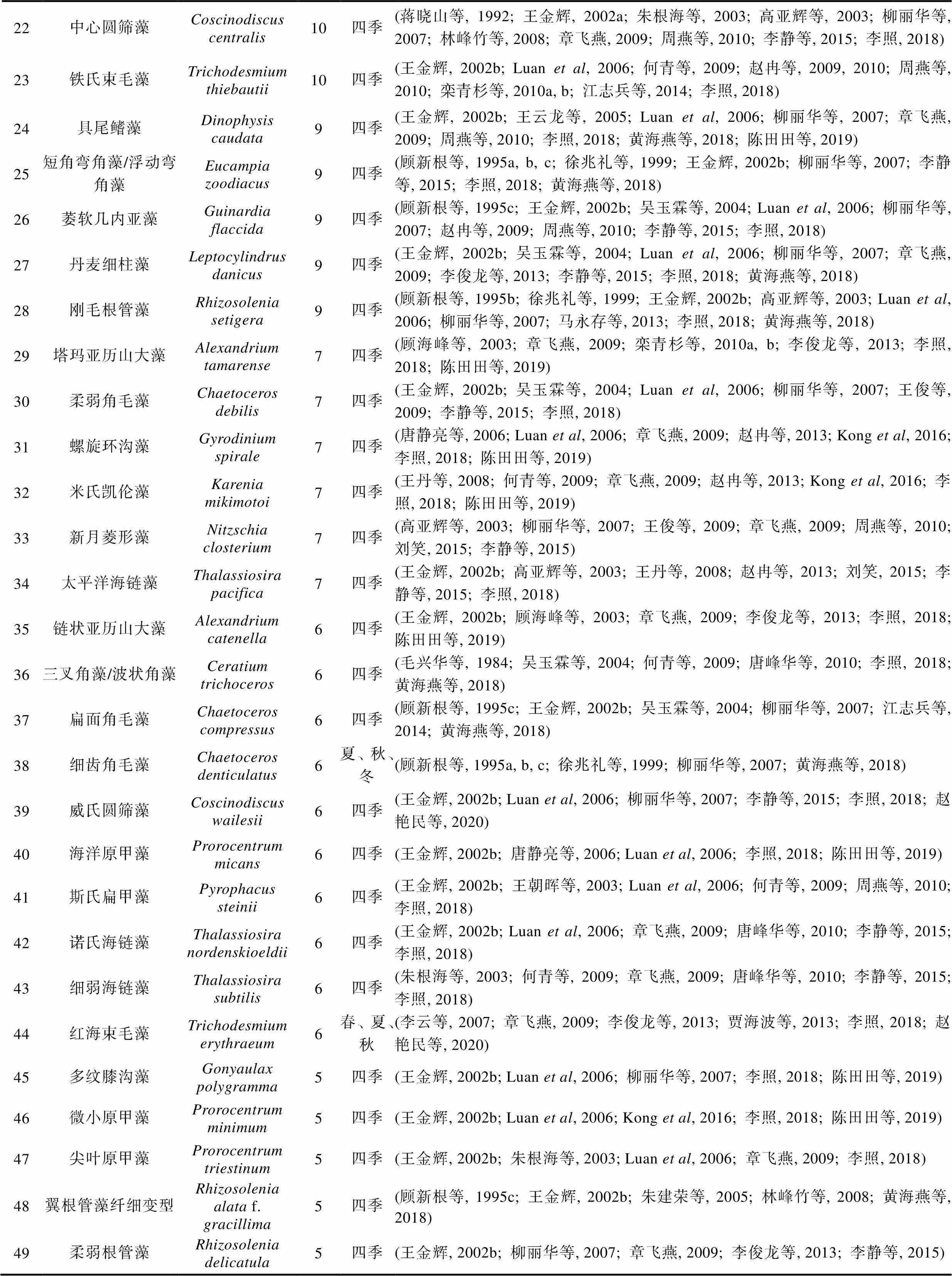
续表

注: a. 长江口航次赤潮物种的相对频次, 上方的条形图显示在一个或多个航次中鉴定到的物种, 左方的条形图显示每一个航次鉴定到的所有浮游植物物种数; b. 长江口历史文献中报道的优势赤潮物种(出现频次>1)词云图; c. 长江口历史文献中报道的优势赤潮物种(出现频次>1)柱状图
4.1 中肋骨条藻
中肋骨条藻是一种广温、广盐的近岸性硅藻, 细胞呈透镜形或圆柱形, 直径6—7 μm, 壳面圆而鼓起, 有一圈细长的刺, 与相邻细胞的对应刺相接组成长链(王金辉, 2002b)。细胞间隙长短不一, 通常大于细胞本身的长度。色素体数目1—10个, 通常有2个, 位于壳面, 各向一面弯曲(郭皓, 2004)。据报道, 中肋骨条藻是长江口及其邻近海域赤潮物种中个体密度最大的河口半咸水种, 最适水温25°C左右、盐度为14—20左右, 几乎一年四季都是长江口及其邻近海域的优势种(王金辉, 2002b)。中肋骨条藻无毒, 增殖速度很快, 其生长和生理状态受氮磷比的影响比较明显, 主要受氮的限制(刘东艳等, 2002)。据1986—1993年间长江口及邻近海域资料统计, 中肋骨条藻赤潮占该区赤潮发生总数的27%, 自1981—2011年间长江口及其邻近海域由中肋骨条藻引发的赤潮多达80起(徐韧等, 1994; 杨庶, 2013)。1991年8月该海域发生中肋骨条藻赤潮, 海水呈暗绿色均匀状态, 细胞密度达1.07×1010cells/m3, 发生过程和夜光藻交替成为优势种(洪君超等, 1993)。在52个航次调查中, 有42次鉴定到中肋骨条藻, 表明它的普遍性。中肋骨条藻引发的赤潮可导致大西洋大马哈鱼的鱼鳃受损死亡(Kent, 1995)。在经济海藻养殖区, 经常与海带、紫菜等争夺营养, 其大量增殖造成经济藻类变色, 甚至腐烂而失去食用和商业价值(陆斗定等, 2000)。2005年, Sarno等人发现骨条藻属除了中肋骨条藻以外, 还有很多其他骨条藻物种, 共鉴定出了8个骨条藻物种(Sarno, 2005)。随后, 我国学者对我国海域骨条藻的研究鉴定出了5个骨条藻物种(Cheng, 2008)。最近, 我们实验室结合宏条形码分析方法和单株系分子分析方法发现胶州湾的优势骨条藻物种并非以前文献中报道的中肋骨条藻, 而是玛氏骨条藻() (Liu, 2020)。因此, 长江口海域的骨条藻物种组成需要进一步分析。
4.2 东海原甲藻
东海原甲藻是大洋性或近岸浮游性甲藻, 单细胞或连接成链状群体。细胞小型至中等, 长度在16—22 μm左右, 宽9—14 μm, 厚7—9 μm, 细胞呈不对称梨形, 顶部稍微凹陷, 后部逐渐成锥状, 壳面观呈倒披针形, 前端平截, 无顶刺(Lu, 2001)。板状色素体2个, 黄褐色(郭皓, 2004)。东海原甲藻适温适盐范围分别为14—22°C、14—32, 最适温盐度分别为18—21°C、21—31。具有明显的昼夜垂直移动, 主要分布于表层水体, 在获得光能、营养盐方面更具优势。东海原甲藻分布于我国东南沿海, 5月份是繁殖繁盛期, 容易发生赤潮, 在河口低盐水域和高温季节不易发生赤潮。很多研究发现, 长江口邻近海域的赤潮优势类群已经由硅藻逐渐转变为甲藻, 据统计自1981—2011年, 长江口邻近海域由东海原甲藻引发的赤潮多达135起(杨庶, 2013)。2000—2002年5月连续三年在长江口舟山海域发生大规模的东海原甲藻赤潮, 最大范围达7000 km2, 持续时间长达1个月(王金辉, 2003)。在52个航次调查中, 有15次发现了东海原甲藻。研究发现东海原甲藻没有明显的急性毒性, 但是室内模拟实验和野外调查发现, 当东海原甲藻达到赤潮密度时, 会对黄海和东海的重要浮游动物中华哲水蚤()的产卵率产生显著的抑制效应, 影响其种群丰度, 有可能危及渔业资源(于仁成等, 2017)。不过, 学术界对引起东海长江口海域赤潮大面积暴发的原甲藻赤潮的原因种仍然有争执。存在两种不同的观点, 一种观点认为东海长江口赤潮暴发的原因种是陆斗定等人于2001年发表的新种东海原甲藻, 依据是通过从东海分离的原甲藻的显微和超微结构, 并与具齿原甲藻()模式种和 Schiller 鉴定的钝头原甲藻之间的形态结构和个体大小具有很大的差别(Lu, 2001; 齐雨藻等, 2003)。另外一种观点则认为东海原甲藻就是具齿原甲藻。吕颂辉等对东海赤潮原甲藻标本和美国国家海洋中心(CCMP)的具齿原甲藻藻株(CCM P1517)通过光镜鉴定和扫描电镜对其细胞表面结构进行观察和比较, 认为生长在东海水域的赤潮原甲藻是具齿原甲藻而非新种。罗立明等通过利用ITS和18S rDNA两种分子标记, 对东海原甲藻和美国国家海洋藻种保藏中心(CCM P)的具齿原甲藻进行分子鉴定, 结果也认为东海原甲藻和美国国家海洋藻种保藏中心的具齿原甲藻应为同一个种(吕颂辉等, 2003; 罗立明等, 2006)。因此, 东海长江口海域原甲藻赤潮原因种还需要进一步跟踪研究。
4.3 旋链角毛藻
旋链角毛藻()是广温性沿岸种。细胞借角毛基部交叉组成螺旋状的群体。宽壳环面为四方形, 宽7—30 μm。壳面椭圆形, 两边稍平, 角毛细而平滑, 自细胞角生出, 皆弯向链凸的一侧。色素体1个, 位于壳面中央(郭皓, 2004)。其生长适宜的盐度范围在25—28。自2006—2011年间旋链角毛藻在长江口海域引发的赤潮共计11起(杨庶, 2013)。2007年7月在长江口舟山海域暴发了角毛藻赤潮, 优势种为旋链角毛藻和扁面角毛藻(), 面积最大时达400 km2(费岳军等, 2008)。在52个航次调查中, 有17次发现了旋链角毛藻。
4.4 夜光藻
夜光藻藻体近圆球形, 游泳生活, 其细胞直径可达150—2000 μm, 肉眼可见。细胞壁透明, 由2层胶状物质组成, 表面有许多微孔。细胞中央有一个大液泡, 细胞质包着细胞核中央团, 并呈分散的细条分布于细胞表面(吕颂辉等, 1992)。口腔位于细胞前端, 并有1条长的触手, 触手基部有一条短小的鞭毛, 靠近触手的齿状突为横沟退化的痕迹, 纵沟在细胞的腹面中央。细胞背部有一杆状器, 使细胞作前后游动(周近明等, 1996)。夜光藻有眼点, 能产生生物荧光(郭皓, 2004)。夜光藻对环境的适应能力很强, 长江口夜光藻的温度和盐度的适应范围分别为12—27°C、25—30, 最适温度和盐度分别为25°C和27。20世纪80年代长江口邻近海域几乎每年都会发生夜光藻赤潮, 据统计1981—2011年间长江口由夜光藻引发的赤潮多达52起(王金辉, 2002b; 杨庶, 2013)。1988年在花鸟山海域发生面积达6600 km2的赤潮(徐韧等, 1994), 蒋晓山等(1992)跟踪观测了1990年6月发生在长江口及其邻近海域的夜光藻赤潮过程。夜光藻可以产生三甲胺, 通过食物链在鱼虾贝类体内聚集, 如被人食用有中毒危险。在52个航次调查中, 有17次发现了夜光藻。
4.5 米氏凯伦藻
米氏凯伦藻是世界性广布种, 细胞长15.6—31.2 μm, 宽13.2—24.0 μm, 营游泳生活。细胞背腹面观近圆形, 上锥部为半球形或宽圆锥形, 下锥部的底部中央有明显的凹陷, 右侧底端略长于左侧, 色素体10—16个(郭皓, 2004)。横沟位于细胞中央, 略靠上壳部, 始末位移为2倍横沟宽度。顶沟直, 始于横沟起点的右侧略上处, 经细胞顶部延伸至上壳部背部。纵沟入侵上壳部, 与顶沟形成一个明显的顶沟-纵沟连接结构。细胞核椭圆形, 位于下壳部左侧, 近细胞边缘(吕颂辉等, 2019)。米氏凯伦藻具有多种营养盐消化吸收途径, 以无机营养盐为主, 但是也能够吸收有机营养盐, 还具备吞噬营养的能力, 使其在种群竞争中处于优势, 经常在河口、上升流、锋面、层化水体等富营养的海域暴发赤潮。据统计, 我国沿海自1998年以来共发生米氏凯伦藻赤潮近90余次(Lu, 2014)。自1981—2011年间, 长江口及其邻近海域由米氏凯伦藻引发的赤潮有46起, 较大规模的有4次左右(杨庶, 2013; 吕颂辉等, 2019)。米氏凯伦藻能产生毒素, 通过食物链传递最终威胁人类健康, 并破坏生态环境。在52个航次调查中, 有7次发现了米氏凯伦藻。
4.6 尖刺拟菱形藻
尖刺拟菱形藻为广温性近岸种类。细胞细长, 梭形, 末端尖。细胞长80—134 μm, 宽3.7—9.0 μm。细胞末端相叠成链, 相连部分达细胞长度的1/4—1/3。每个细胞有两个色素体, 位于细胞核两侧(郭皓, 2004)。尖刺拟菱形藻通常不具毒性, 仅在新西兰和美国西海岸报道过有毒藻株。据统计自1981—2011年间, 长江口及其邻近海域由尖刺拟菱形藻引发的赤潮有5起(杨庶, 2013)。在52个航次调查中, 有17次发现了尖刺拟菱形藻。
5 主要结论与展望
我们通过对长江口历史文献资料的整理汇总, 主要得到如下几点结论:
(1) 长江口海域浮游植物的生物多样性很高, 以硅藻和甲藻为主, 硅藻物种最多, 甲藻次之。在长江口所有浮游植物中, 中肋骨条藻是航次调查报道中出现次数最多的物种。出现次数较高的物种还包括东海原甲藻和虹彩圆筛藻。值得强调的是, 虽然大多数报道认为在近几十年里, 长江口的硅藻组成在逐步下降, 而甲藻组成逐步上升, 我们对航次调查文献的比较分析并没有发现硅藻和甲藻组成的显著变化。
(2) 长江口及其邻近海域是我国近海海域赤潮高发区之一, 赤潮物种也具有较高的生物多样性。根据过去40年内完成的52个航次调查报告, 我们统计到125个赤潮物种, 包括硅藻69种、甲藻49种、金藻2种、隐藻1种和蓝藻3种。需要强调的是, 虽然这些赤潮物种在长江口海域的航次调查中被发现, 但是并非所有的赤潮物种都引发过赤潮。只有在合适的环境因子和营养状态等条件下, 赤潮物种才会引发赤潮。其中中肋骨条藻、东海原甲藻、旋链角毛藻、夜光藻、米氏凯伦藻、尖刺拟菱形藻等是长江口及其邻近海域的主要赤潮物种。
(3) 迄今针对长江口浮游植物的研究大多利用传统的形态学方法观察和鉴定物种, 这种方法要求研究人员具有熟练的经验准确分辨不同物种, 他们对不同浮游植物种内形态多样性的判断并不准确, 不能充分区分形态相似的隐存种, 对尺寸较小的浮游植物细胞更是无能为力。目前, 纯粹基于形态观察的浮游植物研究策略逐步被综合利用形态观察和分子分析的研究策略所代替(陈楠生, 2020)。近年来分子分析方法得到高速发展, 基于DNA分子标记扩增和测序的宏条形码分析方法(metabarcoding analysis)逐渐应用到野外样本的研究中, 并已经取得很多有益的结果。比如, Liu等(2017)结合形态学观察和基于18S rDNA V2—V3区的高通量测序分析, 研究了黄海和东海海域的原生生物, 发现寄生性甲藻虽然在高通量测序方法中占优势, 它们在形态学研究中却被忽略, 可见分子分析的方法能够发现传统形态学研究中被遗漏的物种。可以预期, 基于形态学方法的研究与基于分子生物学方法的研究的结合应用将全面准确揭示自然样本中物种的组成及丰度。
(4) 长江口海域浮游植物和赤潮物种都很丰富, 但是目前代表性分子标记(比如18S rDNA, 28S rDNA, ITS,,)数据库中代表长江口海域浮游植物和赤潮物种的数据有限, 严重限制了组学分析方法的充分利用。需要对长江口浮游植物特别是赤潮物种进行系统性的分子分析, 获得重要分子标记(比如18S rDNA)序列信息, 丰富数据库, 促进组学分析方法在研究长江口赤潮中的利用。
于仁成, 张清春, 孔凡洲等, 2017. 长江口及其邻近海域有害藻华的发生情况、危害效应与演变趋势. 海洋与湖沼, 48(6): 1178—1186
于仁成, 吕颂辉, 齐雨藻等, 2020. 中国近海有害藻华研究现状与展望. 海洋与湖沼, 51(4): 768—788
上官欣欣, 薛俊增, 吴惠仙, 2019. 长江口浮游植物群落结构的特征. 上海海洋大学学报, 28(5): 689—698
马永存, 徐 韧, 何培民等, 2013. 长江口低氧区及邻近海域浮游植物群落初步研究. 上海海洋大学学报, 22(6): 903—911
王 丹, 孙 军, 周 锋等, 2008. 2006年6月长江口低氧区及邻近水域浮游植物. 海洋与湖沼, 39(6): 619—627
王 俊, 陈瑞盛, 左 涛, 2009. 三峡工程截流后长江口邻近海域的网采浮游植物群落结构特征. 水生态学杂志, 2(2): 80—87
王云龙, 袁 骐, 沈新强, 2005. 长江口及邻近水域春季浮游植物的生态特征. 中国水产科学, 12(3): 300—306
王云龙, 袁 骐, 沈新强, 2008. 长江口及邻近海域夏季浮游植物分布现状与变化趋势. 海洋环境科学, 27(2): 169—172
王江涛, 曹 婧, 2012. 长江口海域近50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 31(3): 310—315
王金辉, 2002a. 长江口3个不同生态系的浮游植物群落. 青岛海洋大学学报, 32(3): 422—428
王金辉, 2002b. 长江口邻近水域的赤潮生物. 海洋环境科学, 21(2): 37—41
王金辉, 2003. 东海赤潮生物具齿原甲藻及其特征. 浙江海洋学院学报(自然科学版), 22(2): 128—157
王金辉, 黄秀清, 刘阿成等, 2004. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 23(1): 32—39
王朝晖, 齐雨藻, 2003. 甲藻孢囊在长江口海域表层沉积物中的分布. 应用生态学报, 14(7): 1039—1043
中国科学技术协会, 中国海洋学会, 2015. 中国海洋学学科史. 北京: 中国科学技术出版社, 297
毛兴华, 李瑞香, 1984. 东海北部陆架区浮游甲藻的分布及其生态特征. 海洋洋报, 6(5): 672—677
叶海新, 上官欣欣, 毕子昊等, 2019. 长江口北支贝类放流滩涂区潮下带水域浮游植物分析. 上海海洋大学学报, 28(5): 699—707
吕颂辉, 齐雨藻, 1992. 南海大鹏湾的主要赤潮生物. 暨南大学学报(自然科学), 13(3): 130—133
吕颂辉, 张玉宇, 陈菊芳, 2003. 东海具齿原甲藻的扫描电子显微结构. 应用生态学报, 14(7): 1070—1072
吕颂辉, 岑竞仪, 王建艳等, 2019. 我国近海米氏凯伦藻()藻华发生概况、危害及其生态学机制. 海洋与湖沼, 50(3): 487—494
朱建荣, 王金辉, 沈焕庭等, 2005. 2003年6月中下旬长江口外海区冲淡水和赤潮的观测及分析. 科学通报, 50(1): 59—65
朱根海, 许卫忆, 朱德第等, 2003. 长江口赤潮高发区浮游植物与水动力环境因子的分布特征. 应用生态学报, 14(7): 1135—1139
刘 笑, 2015. 长江口浮游植物与轮虫群落生态研究. 上海: 上海海洋大学硕士学位论文, 1—41
刘东艳, 孙 军, 陈宗涛等, 2002. 不同氮磷比对中肋骨条藻生长特性的影响. 海洋湖沼通报, (2): 39—44
刘录三, 李子成, 周 娟等, 2011. 长江口及其邻近海域赤潮时空分布研究. 环境科学, 32(9): 2497—2504
齐雨藻, 王 艳, 2003. 我国东海赤潮原甲藻应属哪种?. 应用生态学报, 14(7): 1188—1190
江志兵, 曾江宁, 李宏亮等, 2014. 长江口及其邻近陆架区夏季网采浮游植物及其影响因素. 海洋学报, 36(6): 112—123
汤 琳, 张锦平, 许兆礼等, 2007. 长江口邻近水域浮游植物群落动态变化及其环境因子的研究. 中国环境监测, 23(2): 97—101
李 云, 李道季, 唐静亮等, 2007. 长江口及毗邻海域浮游植物的分布与变化. 环境科学, 28(4): 719—729
李 英, 吕颂辉, 徐 宁等, 2005. 东海原甲藻对不同磷源的利用特征. 生态科学, 24(4): 314—317, 321
李 照, 2018. 长江口邻近海域浮游植物群落特征及其碳沉降研究. 青岛: 中国科学院大学(中国科学院海洋研究所)博士学位论文, 1—111
李 静, 陈长平, 梁君荣等, 2015. 2010年春秋季长江口南部硅藻种类组成和密度的时空变化. 应用海洋学学报, 34(3): 372—387
李俊龙, 郑丙辉, 刘录三等, 2013. 长江口浮游植物群落特征及其与环境的响应关系. 环境科学研究, 26(4): 403—409
杨 庶, 2013. 长江口及邻近海域浮游植物生长温度效应研究——长江口及邻近海域浮游植物生物量年际变化的温度控制效应. 青岛: 中国海洋大学博士学位论文, 1—73
吴玉霖, 傅月娜, 张永山等, 2004. 长江口海域浮游植物分布及其与径流的关系. 海洋与湖沼, 35(3): 246—251
何 青, 孙 军, 栾青杉等, 2007. 长江口及其邻近水域冬季浮游植物群集. 应用生态学报, 18(11): 2559—2566
何 青, 孙 军, 2009. 长江口及其邻近水域网采浮游植物群落. 生态学报, 29(7): 3928—3938
张锦平, 夏 静, 2006. 长江口浮游植物群落的聚类分析. 上海水产大学学报, 15(4): 456—460
陆斗定, Gobel J, 王春生等, 2000. 浙江海区赤潮生物监测与赤潮实时预测. 东海海洋, 18(2): 33—44
陈田田, 宋书群, 刘 云等, 2019. 2015—2016年长江口及其邻近海域甲藻群落与阿米巴藻感染的周年变化. 海洋与湖沼, 50(1): 139—148
陈楠生, 2020. 有害藻华的宏条形码分析: 机会与挑战. 海洋科学, 44(7): 116—134
林 军, 朱建荣, 张 经等, 2011. 长江口外海区浮游植物生物量分布及其与环境因子的关系. 水产学报, 35(1): 74—87
林 梅, 李 扬, 2017. 长江口海域春季浮游植物的年际变化. 海洋与湖沼, 48(2): 303—311
林峰竹, 吴玉霖, 于海成等, 2008. 2004年长江口浮游植物群落结构特征分析. 海洋与湖沼, 39(4): 401—410
罗立明, 胡鸿钧, 李夜光等, 2006. 东海原甲藻的分子鉴定. 海洋学报, 28(1): 127—131
周 燕, 赵聪蛟, 高元森等, 2010. 2005~2008年杭州湾生态监控区浮游植物分布特征及变化趋势. 海洋学研究, 28(2): 28—35
周名江, 颜 天, 邹景忠, 2003. 长江口邻近海域赤潮发生区基本特征初探. 应用生态学报, 14(7): 1031—1038
周近明, 朱明远, 1996. 大鹏湾的赤潮生物. 中国赤潮研究: SCOR-IOC赤潮工作组中国委员会第二次会议选. 青岛: 青岛出版社, 21—30
赵 冉, 白 洁, 孙 军等, 2009. 2006年夏季长江口及其邻近水域浮游植物群集. 海洋湖沼通报, (2): 88—96
赵 冉, 孙 军, 白 洁, 2010. 2006年秋季长江口及其邻近水域浮游植物群集. 海洋科学, 34(4): 32—39
赵 冉, 孙 军, 宋书群, 2013. 2006年春季长江口及其邻近水域浮游植物. 海洋通报, 32(4): 421—428
赵保仁, 1991. 长江冲淡水的转向机制问题. 海洋学报, 13(5): 600—610
赵艳民, 马迎群, 曹 伟等, 2020. 长江口2016年冬季浮游植物类群及其与环境因子的关系. 海洋环境科学, 39(2): 183—188
柳丽华, 左 涛, 陈瑞盛等, 2007. 2004年秋季长江口海域浮游植物的群落结构和多样性. 海洋水产研究, 28(3): 112—119
俞志明, 陈楠生, 2019. 国内外赤潮的发展趋势与研究热点. 海洋与湖沼, 50(3): 474—486
洪君超, 黄秀清, 蒋晓山等, 1993. 嵊山水域中肋骨条藻赤潮发生过程主导因子分析. 海洋学报, 15(6): 135—141
费岳军, 蒋 红, 2008. 舟山朱家尖海域角毛藻赤潮与环境因子关系的研究. 海洋环境科学, 27(S1): 38—41
贾海波, 胡颢琰, 邵君波等, 2013. 长江口及其邻近海域春秋季浮游植物群落及环境影响因子研究. 海洋环境科学, 32(6): 851—855
顾海峰, 蓝东兆, 方 琦等, 2003. 我国东南沿海亚历山大藻休眠孢囊的分布和萌发研究. 应用生态学报, 14(7): 1147—1150
顾新根, 袁 骐, 沈焕庭等, 1995a. 长江口最大浑浊带浮游植物的生态研究. 中国水产科学, 2(1): 16—27
顾新根, 袁 骐, 杨蕉文等, 1995b. 长江口羽状锋海区浮游植物的生态研究. 中国水产科学, 2(1): 1—15
顾新根, 袁 骐, 杨蕉文等, 1995c. 长江口外水域浮游植物垂直分布研究. 中国水产科学, 2(1): 28—38
徐 韧, 洪君超, 王桂兰等, 1994. 长江口及其邻近海域的赤潮现象. 海洋通报, 13(5): 25—29
徐兆礼, 白雪梅, 袁 骐等, 1999. 长江口浮游植物生态研究. 中国水产科学, 6(5): 52—54
栾青杉, 孙 军, 2010a. 2005年夏季长江口水域浮游植物群集特征及其与环境因子的关系. 生态学报, 30(18): 4967—4975
栾青杉, 孙 军, 2010b. 2005年秋季长江口及其邻近水域浮游植物群集. 长江流域资源与环境, 19(2): 202—208
高亚辉, 虞秋波, 齐雨藻等, 2003. 长江口附近海域春季浮游硅藻的种类组成和生态分布. 应用生态学报, 14(7): 1044—1048
郭 皓, 2004. 中国近海赤潮生物图谱. 北京: 海洋出版社, 34—93
唐峰华, 伍玉梅, 樊 伟等, 2010. 长江口浮游植物分布情况及与径流关系的初步探讨. 生态环境学报, 19(12): 2934—2940
唐静亮, 毛宏跃, 过美蓉, 2006. 浙江北部海域的具齿原甲藻赤潮分析. 海洋环境科学, 25(3): 63—66
黄海燕, 王秋璐, 许艳等, 2018. 2004—2016年夏季长江口网采浮游植物及其影响因素. 海洋与湖沼, 49(2): 319—330
章飞燕, 2009. 长江口及邻近海域浮游植物群落变化的历史对比及其环境因子研究. 上海: 华东师范大学硕士学位论文, 1—57
蒋晓山, 洪君超, 王桂兰等, 1992. 长江口赤潮多发区夜光藻()赤潮发生过程分析. 暨南大学学报(自然科学), 13(3): 134—139
蒲新明, 2000. 长江口区浮游植物营养限制因子的研究. 青岛: 中国科学院海洋研究所硕士学位论文, 1—59
Cheng J F, Li Y, Liang J R, 2008. Morphological variability and genetic diversity in five species of(Bacillariophyta). Progress in Natural Science, 18(11): 1345—1355
Gao X L, Song J M, 2005. Phytoplankton distributions and their relationship with the environment in the Changjiang Estuary, China. Marine Pollution Bulletin, 50(3): 327—335
Hallegraeff G M, 2010. Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge. Journal of Phycology, 46(2): 220—235
Jiang Z B, Liu J J, Chen J F, 2014. Responses of summer phytoplankton community to drastic environmental changes in the Changjiang (Yangtze River) estuary during the past 50 years. Water Research, 54: 1—11
Jiang T, Yu Z M, Song X X, 2010. Long-term ecological interactions between nutrient and phytoplankton community in the Changjiang estuary. Chinese Journal of Oceanology and Limnology, 28(4): 887—898
Kent M L, Whyte J N C, Latrace C, 1995. Gill lesions and mortality in seawater pen-reared Atlantic salmonassociated with a dense bloom ofandspecies. Diseases of Aquatic Organisms, 22(1): 77—81
Kong F Z, Xu Z J, Yu R C, 2016. Distribution patterns of phytoplankton in the Changjiang River estuary and adjacent waters in spring 2009. Chinese Journal of Oceanology and Limnology, 34(5): 902—914
Li Y, Li D J, Tang J L, 2010. Long-term changes in the Changjiang Estuary plankton community related to anthropogenic eutrophication. Aquatic Ecosystem Health & Management, 13(1): 66—72
Li H M, Tang H J, Shi X Y, 2014. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased Harmful Algal Blooms. Harmful Algae, 39: 92—101
Liu S Y, Gibson K, Cui Z M, 2020. Metabarcoding analysis of harmful algal species in Jiaozhou Bay. Harmful Algae, 92: 101772
Liu Y, Song S Q, Chen T T, 2017. The diversity and structure of marine protists in the coastal waters of China revealed by morphological observation and 454 pyrosequencing. Estuarine, Coastal and Shelf Science, 189: 143—155
Lu D D, Goebel J, 2001. Five red tide species in genusincluding the description ofLu SP. nov. from the East China Sea. Chinese Journal of Oceanology and Limnology, 19(4): 337—344
Lu D D, Qi Y Z, Gu H F, 2014. Causative species of harmful algal blooms in Chinese coastal waters. Algological Studies, 145—146(1): 145—168
Luan Q S, Jun S, Shen Z L, 2006. Phytoplankton assemblage of Yangtze River Estuary and the adjacent east China Sea in summer, 2004. Journal of Ocean University of China, 5(2): 123—131
Ning X R, Vaulot D, Liu Z S, 1988. Standing stock and production of phytoplankton in the estuary of the Changjiang (Yangtse River) and the adjacent East China Sea. Marine Ecology - Progress Series, 49(1—2): 141—150
Sarno D, Kooistra W H C F, Medlin L K, 2005. Diversity in the genus(Bacillariophyceae). Ⅱ. An assessment of the taxonomy of-like species with the description of four new species. Journal of Phycology, 41(1): 151—176
Yang S, Han X R, Zhang C S, 2014. Seasonal changes in phytoplankton biomass and dominant species in the Changjiang River Estuary and adjacent seas: General trends based on field survey data 1959—2009. Journal of Ocean University of China, 13(6): 926—934
Zhou M J, Shen Z L, Yu R C, 2008. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Continental Shelf Research, 28(12): 1483—1489
ADVANCES IN THE STUDY OF BIODIVERSITY OF PHYTOPLANKTON AND RED TIDE SPECIES IN CHINA (Ⅳ): THE CHANGJIANG ESTUARY
CHEN Nan-Sheng1, 2, 3, CUI Zong-Mei1, 2, 3, 4, XU Qing1, 2, 3, 5
(1. CAS Key Laboratory of Marine Ecology and Environmental Science, Institute of Oceanology,Chinese Academy of Sciences,Qingdao 266071,China; 2. Functional Laboratory for Marine Ecology and Environmental Science,Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao266237,China; 3. Center for Ocean Mega-Science, Chinese Academy of Sciences,Qingdao266071, China; 4. University of Chinese Academy of Sciences, Beijing 100049, China; 5.Huazhong Agricultural University, Wuhan 430070, China)
The Changjiang (Yangtze) River estuary is a special marine region in China. It has a high phytoplankton diversity, including a variety of red tide species, with frequent red-tide outbreaks in recent years. In this review, the compositions of phytoplankton identified in 52 reported expeditions in the Changjiang Estuary were examined and compared, with a focus on red tide species. Of the 657 phytoplankton described in these expeditions (including 571 species and 86 unidentified species), diatoms (368 species and 55 unidentified species), and dinoflagellates (149 species and 15 unidentified species) were the dominant phyla. Among these phytoplankton, 125 species were identified as red tide species in the Changjiang Estuary, including 69 diatoms, 49 dinoflagellates, 2 chrysophytes, 3 cyanobacteria, and 1 cryptophyta. The frequently-seen red-species in the estuary included,,,,and. This review shall provide a reference for studying biodiversity of phytoplankton and HAB (harmful algae bloom) species in different sea areas, discussing the origin and evolution of biodiversity, and facilitating the interpretation of metagenomics and macrobarcoding results.
Changjiang Estuary; biodiversity; expedition; red tide species; phytoplankton
附表1、附表2、附表3见http://dx.doi.org/10.11693/hyhz20200800236
* 中国科学院战略性先导科技专项(B类), XDB42000000号。陈楠生, 博士生导师, 研究员, E-mail: chenn@qdio.ac.cn
2020-08-22,
2020-10-20
X172; S944.3+49; S963.21+3
10.11693/hyhz20200800236
猜你喜欢
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
法医学杂志(2022年1期)2022-06-21
小天使·三年级语数英综合(2022年3期)2022-04-17
法人(2022年3期)2022-03-30
华东师范大学学报(自然科学版)(2019年2期)2019-06-11
水运管理(2017年10期)2017-11-20
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
棋艺(2001年11期)2001-05-21
