
华北地区4个品种樱花温度抗逆性研究
2021-03-24 09:23侯世星刘东骏
中国农学通报 2021年3期
胡 娜,陈 思,侯世星,刘东骏
(1北京市玉渊潭公园管理处,北京100142;2北京电子科技职业学院,北京100176)
0 引言
樱花是著名的木本观赏花卉,隶属蔷薇科(Rosaceae)李亚科(Prunusoidea)樱属(Cerasus),原产北半球温带环喜马拉雅山地区,在世界各地有广泛分布[1]。不同品种樱花对于温度的抗逆性不同,因此适宜种植的地区也不尽相同。了解不同品种樱花的温度适应性,选择适宜的立地环境进行栽植能获得良好的园林景观效果。在有关研究中,叶超宏[2]对珠江三角地区适种的樱花进行了比较,筛选出了部分耐热性良好的樱花品种。马祥宾等[3]对鼋头渚10个樱属种(品种)进行了耐寒性生理指标测定,并进行了综合评价。郭学民等[4]对比了桃、杏和樱桃树的抗寒性,为果树抗寒品种的选育提供了参考。张生智等[5]对樱桃优良品种枝条的耐寒性进行了生理生化分析,不同品种抗寒性存在差异。
由原生种樱花寒绯樱(Cerasus campanulata)与原生种樱花大岛樱(Cerasus spesiosa)自然杂交或人工培育而得到的一系列品种樱花,如‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’等[6],因花期早、花色艳丽、着花量大,近年来愈发受到游客的喜爱。这类樱花在国内华东地区广泛栽植,并且适应性良好,而华北地区相关报道较少。为观测这类樱花在华北地区的温度抗逆性,笔者测定上述4个品种在高温胁迫下(25~45℃)叶片的叶绿素含量、脯氨酸含量,低温胁迫下(-20~0℃)枝条的超氧化物歧化酶含量、丙二醛含量,并结合田间观测进行综合分析,以期为其在华北地区的栽植提供参考。
1 材料与方法
1.1 试验地与试验材料
试验于2018年11月—2020年8月在北京市玉渊潭公园进行。玉渊潭公园地处北京市海淀区西三环内(东经116°18′、北纬39°55′),自1973年种植樱花以来已有40余年的历史,是国内北方地区著名的樱花园。属温带季风性气候。平均海拔32 m,年均日照2780 h,年均气温13℃,近10年极端最低气温-16℃,最高气温41℃。近10年年均降水量540.6 mm,汛期(6月1日—8月31日)平均降水量384.5 mm。试验材料名称、来源地、规格及数量见表1。

表1 试验材料名称及来源地
1.2 试验方法
1.2.1 耐热性试验 7月下旬剪取观测植株外围中部各方向、节间长度与直径相仿、无病虫害且生长势良好的1年生带叶枝条。每株观测植株取5枝,每个品种共计25枝。分为5组,每组包含每株观测植株各1枝,下部插入蒸馏水中。根据北京地区近年来最高气温记录,将样品放入不同人工培养箱中,光照强度3000 lx,分别升温至25、30、35、40、45℃,萎蔫14 h。处理后的每组枝条,分别取同一品种5枝枝条上的叶片,剪碎后混合均匀,等分为3份。对每份叶片利用植物叶绿素含量测试盒(苏州科铭生物技术有限公司)提取叶片中叶绿素,采用可见光分光光度计法测定含量[7]。同时对每份叶片利用脯氨酸含量测试盒(苏州科铭生物技术有限公司)提取叶片中脯氨酸,采用茚三酮法测定含量[7]。
1.2.2 耐寒性试验 12月下旬剪取观测植株外围中部各方向、节间长度与直径相仿、无病虫害且生长势良好的1年生枝条。每株观测植株取5枝,每个品种共计25枝。分为5组,每组包含每株观测植株各1枝,用保鲜膜包裹。根据北京地区近年来最低气温记录,将样品放入低温恒温槽中,分别降温至0、-5、-10、-15、-20℃,冷冻24 h。处理后的每组枝条,分别将同一品种5枝枝条剪碎后混合均匀,等分为3份。对每份枝条利用丙二醛含量测试盒(苏州科铭生物技术有限公司)提取枝条中丙二醛,采用硫代巴比妥酸(TBA)缩合法测定含量[8]。同时对每份枝条利用超氧化物歧化酶含量测试盒(苏州科铭生物技术有限公司)提取枝条中超氧化物歧化酶,采用抑制氮蓝四唑(NBT)光还原法测定含量[9]。
1.2.3 田间观测评定 于2019年6—8月、2020年6—8月进行热胁迫田间观测。参考邱勇波[10]的方法,每10天对所有引种的20株植株进行观测。分级标准为:0级,无热害伤害;Ⅰ级,1/4叶片出现热伤害;Ⅱ级,1/4~1/2叶片出现热伤害;Ⅲ级,1/2~3/4叶片出现热伤害;Ⅳ级,3/4以上叶片出现热伤害。观测期结束后,汇总观测数据,计算每个品种出现热伤害的叶片在该品种全部叶片中的占比,得到该品种的热害等级。
2018年12月—2019年2月、2019年12月—2020年2月进行冷胁迫田间观测。参考李心[11]的方法,每10天对所有引种的20株植株进行观测。分级标准为:0级,无冷害伤害;Ⅰ级,枝条部分褐变;Ⅱ级,枝条全部褐变;Ⅲ级,枝条部分萎蔫;Ⅳ级,枝条全部萎蔫。观测期结束后,汇总观测数据,计算每个品种出现冷伤害的枝条在该品种全部枝条中的占比,得到该品种的冷害等级。
1.3 数据处理
利用SPSS 21.0对数据进行单因素方差(ANOVA)、最小显著差数法(LSD)进行多重分析。
2 结果与分析
2.1 耐热性生理指标
由表2可知,4个品种樱花的叶绿素含量,随着温度的上升,均呈现先上升后下降的趋势。‘大寒樱’在35℃达到最大值,其余3个品种均在40℃达到最大值。‘河津樱’、‘大渔樱’的叶绿素含量在25~35℃之间没有显著差异。‘修善寺寒樱’的叶绿素含量在25~30℃之间没有显著差异。‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’叶绿素最大值比25℃时分别增长115.24%、225.17%、108.43%、75.16%。
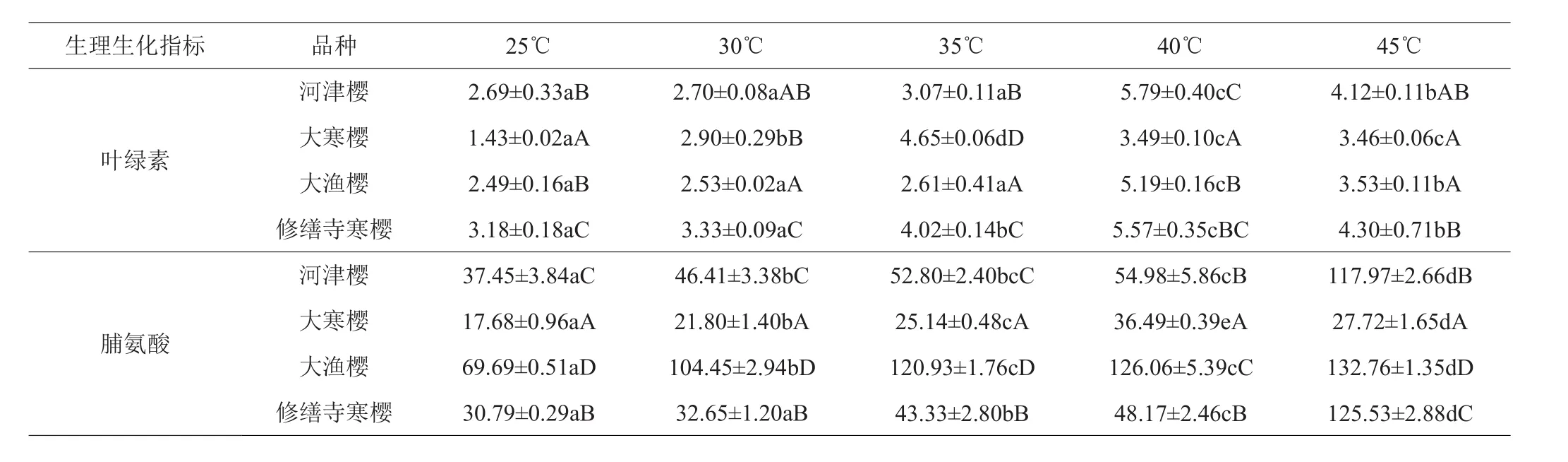
表2 4个品种樱花不同温度下叶绿素含量、脯氨酸含量 μg/(g·鲜重)
4个品种樱花的脯氨酸含量,除‘大寒樱’呈现先上升后下降的趋势,在40℃达到最大值外;其余3个品种均随温度升高持续上升,在45℃达到最大值。‘河津樱’与‘修善寺寒樱’在25~40℃脯氨酸增幅较缓,在45℃突然急剧增加。‘大渔樱’的脯氨酸含量始终处于较高水平,而‘大寒樱’的脯氨酸含量始终处于较低水平。‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’脯氨酸最大值比25℃时分别增长215.01%、106.39%、90.50%、307.70%。
2.2 耐寒性生理指标
由表3可知,4个品种樱花丙二醛含量均随着温度的下降而升高。‘修善寺寒樱’各温度间差异显著‘,河津樱’在-20~-10℃没有显著差异‘,大寒樱’‘、大渔樱’在-15~-10℃没有显著性差异。温度低于-5℃后‘,大寒樱’丙二醛含量较其他品种明显升高。-20℃时‘河津樱’‘、大寒樱’‘、大渔樱’‘、修善寺寒樱’的丙二醛含量比0℃时分别增长119.59%、406.27%、110.47%、291.78%。

表3 4个品种樱花不同温度下丙二醛含量、超氧化物歧化酶含量
4个品种樱花超氧化物歧化酶含量随着温度的下降,均呈现先上升后下降的趋势。其中‘河津樱’、‘大渔樱’在-15℃达到最大值,‘大寒樱’、‘修善寺寒樱’在-10℃达到最大值。‘河津樱’超氧化物歧化酶含量整体处于较高水平。‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’超氧化物歧化酶最大值比0℃时分别增长162.33%、227.79%、231.29%、131.92%。
2.3 田间观测结果
表4为栽植地中2018年冬至2020年夏4个品种樱花叶片热害分级情况及枝条冷害分级情况。在2年的观测中,‘河津樱’与‘大渔樱’越夏没有发生明显困难;但‘大寒樱’出现叶面发黄甚至早落叶的情况;而‘修善寺寒樱’在2019年进入夏季后约1/2的叶面持续出现发黄的情况,2020年越夏没有发生明显困难。越冬过程中,4个品种樱花均进行根系无纺布覆盖、树干缠干的越冬保护。‘河津樱’没有出现冷害;连续2年‘大寒樱’远端枝条出现萎蔫情况;‘大渔樱’在2018—2019年冬远端枝条出现萎蔫情况,2019—2020年冬没有出现冷害;‘修善寺寒樱’在2018—2019年冬远端枝出现部分褐变,2019—2020年冬没有出现冷害。

表4 2018年冬至2020年夏4个品种樱花热害、冷害分级
2.4 温度抗逆性指数
对25、30、35、40、45℃ 5个温度分别赋值1、2、3、4、5,定义各品种叶绿素含量、脯氨酸含量达到最大值时温度对应的数值,为该品种叶绿素含量、脯氨酸含量温度得分。对0、-5、-10、-15、-20℃分别赋值1、2、3、4、5,定义各品种超氧化物歧化酶含量达到最大值时温度对应的数值,为该品种超氧化物歧化酶含量温度得分。对各品种丙二醛最大值与0℃时的增长率赋值,400%~500%、300~400%、200~300%、100~200%、0~100%这5个区间分别赋1、2、3、4、5,得到各品种丙二醛含量涨幅得分。各品种生理生化指标得分情况见表5。
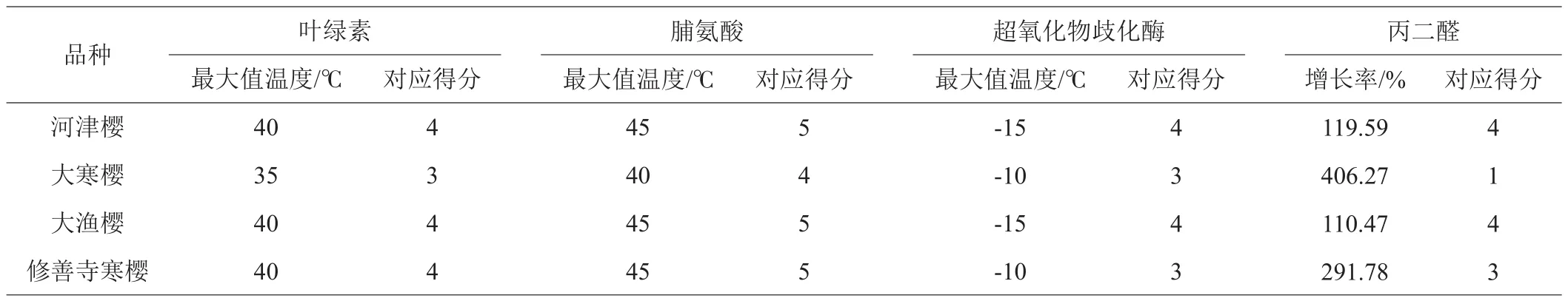
表5 4个品种樱花生理生化指标得分
为对比各品种之间生理生化指标的差异,定义表2中叶绿素含量、脯氨酸含量,表3中超氧化物歧化酶含量中的表示品种间显著性差异的A~D为温度抗逆性增强,赋值1~4[12]。定义表3中丙二醛含量中表示品种间显著性差异中A~C为温度抗逆性减弱,赋值3~1。对于表2、表3中同一数据中的不同显著性差异,以数值较小的进行计算。不同品种间,数值越高表示该品种温度抗逆性越强。各品种间显著性差异得分情况见表6。定义热害等级、冷害等级0~Ⅳ为温度抗逆性减弱,赋值5~1。各品种2年观测周期内观测得分见表7。

表6 品种间显著性差异得分

表7 2018年冬至2020年夏4个品种樱花热害、冷害分级得分
将生理生化指标得分与品种间显著性差异得分、热害、冷害分级得分进行加和,得到各品种温度抗逆指数。‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’的温度抗逆性指数分别为91、57、78、79。指数越大,温度抗逆性越强。结合实际田间观测,‘河津樱’在华北地区的温度抗逆性良好,‘大寒樱’的温度抗逆性弱。
3 结论
试验测定4种品种樱花在高温胁迫下叶片叶绿素含量、脯氨酸含量,低温胁迫下枝条丙二醛含量、超氧化物歧化酶含量等4个生理指标,并结合2年的田间观测进行综合评价,‘河津樱’在华北地区的温度抗逆性较强,‘大寒樱’较弱,‘大渔樱’和‘修善寺寒樱’居中。
在华北地区引种由原生种寒绯樱和原生种大岛樱杂交而得到的上述一系列品种樱花,应充分考虑不同品种的温度抗逆性差异,选择小环境相对良好的区域种植。夏季避免过度的高温暴晒,冬季进行适当的越冬保护,可提高引种成活率。由于该系列樱花具有花期早、花色艳丽的特点,如果能够在华北地区园林景观营造中适当应用,可以在初春形成亮丽的景观效果。
4 讨论
4.1 热胁迫对不同品种樱花叶绿素含量的影响
叶绿素是植物进行光合作用的重要色素,强光和高温的作用易使其分解[13]。在热胁迫下,耐热性好的品种,可维持较高水平的叶绿素含量[14]。过高的温度会导致叶绿素发生分解[15]。试验表明,‘河津樱’、‘大渔樱’在温度低于35℃时,叶绿素含量稳定。随着温度继续升高至40℃,2种樱花的叶绿素含量快速升高达到最大值,表明植物为应对高温逆境做出了相应的生理响应。但随着温度进一步升高到45℃,过高的温度导致叶绿素被破坏,叶绿素的含量出现下降。对比试验数据,‘大寒樱’在35℃叶绿素含量达到最大值,而其他3个品种在40℃时叶绿素含量达到最大值,表明‘大寒樱’的耐热性略弱。实际田间观测中,试验地夏季最高气温在40℃上下,‘河津樱’、‘大渔樱’生长正常,而‘大寒樱’连续2年越夏出现困难,与叶绿素含量测试得到的结果一致。郑书旗等[16]对不同苹果品种进行的耐热性研究表明,高温胁迫下叶片叶绿素含量下降。彭勇政等[12]对5个月季品种进行了耐热性研究表明,部分月季品种的叶绿素含量随温度升高呈现先升高后下降的趋势,与本研究结果相似。
对于华北地区而言,特别是城市中心,受热岛效应影响,夏季极值温度较容易超过35℃,这种环境下,种植‘大寒樱’易发生越夏困难。就北京地区而言,近10年的气象数据中,夏季出现过41℃的高温,此时其余3个品种的叶绿素抗逆机制也受到了挑战。为保障植株安全越夏,遇40℃左右的高温天气,应采取搭建遮阴网、增加喷淋设施等措施,以确保植株的越夏安全。
4.2 热胁迫对不同品种樱花脯氨酸含量的影响
脯氨酸与甜菜碱、甘氨酸等渗透物质的积累,是植物体内抵抗外界胁迫的适应性基质[17]。脯氨酸可以清除活性氧,减少脂质过氧化、清除NH4+及防止有毒氨基酸积累[18]。脯氨酸积累越高,植物耐热性越强[19]。试验结果显示,随着温度的升高,4个品种樱花的脯氨酸含量均有所增加。除‘大寒樱’外,其余3个品种均在45℃达到最大值。而‘大寒樱’在40℃达到最大值,随后有所下降,表明‘大寒樱’的耐热性略弱。这与田间观测中‘大寒樱’的越夏表现最弱一致。申惠翡等[20]对不同杜鹃花品种耐热性的研究表明,不同品种的脯氨酸含量对于热胁迫的反应不同。宋阳[21]对国内北方代表柳树品种进行了研究,发现脯氨酸含量随热胁迫而增加,部分品种脯氨酸含量在升高到一定温度后下降,这与本研究结果基本一致。
结合叶绿素试验数据可以看出,‘大寒樱’的耐热能力较其他3个品种有一定的差距。如果要在华北地区栽植该品种,可选择夏季小环境相对凉爽的区域。此外,也可通过在植株西南方向配植相对高大的乔木的方法以减少夏季西晒对植株的危害。
4.3 冷胁迫对不同品种樱花丙二醛含量的影响
低温会影响植物细胞内的自由基产生和清除,随着自由基累积,细胞膜逐渐受到伤害,造成膜脂过氧化,丙二醛含量上升影响植物体内代谢反应[22]。丙二醛具有细胞毒性,其含量越高表明植物细胞膜受伤程度越大[23]。本试验中,各品种的丙二醛含量均随温度降低而增大。‘河津樱’在-20~-10℃时丙二醛含量变化不大,显示其具有一定耐寒性。‘大寒樱’在冷胁迫过程中,丙二醛增幅超过400%,显示其细胞受到较大伤害,抗寒性较弱。实际田间观测中,在一定的越冬保护措施下,‘河津樱’可以正常越冬,‘大寒樱’的越冬表现最弱,这与本研究结果一致。张志伟[24]对5种棕榈苗木进行了抗寒性研究,发现不同品种对于冷胁迫下丙二醛含量增幅存在差异。李瑞雪等[25]对6种木兰科植物冷胁迫下的生理响应进行了研究,丙二醛含量均随温度降低呈现上升的趋势,与本试验结果基本一致。
华北地区冬季温度随地势变化差异较大,东南部平原地区冬季相对温暖,而西北部山区冬季寒冷。若选择栽植上述4个品种樱花,应充分考量冬季极端最低温度,最低气温频繁在-15℃上下浮动时,不宜种植。此外,种植时应选择背风向阳、冬季小环境相对良好的区域。由于这4个品种樱花由原产自中国福建山区的寒绯樱与日本的大岛樱杂交获得,其耐寒性受亲本寒绯樱的影响,相对较弱。因此,适当的越冬保护也是十分必要的。对根区进行保温覆盖,可以保障早春时节根系的正常萌动,保证植株地上部分的水分供应,防止抽条现象的发生。对植株进行缠干保护,也有助于植株抵抗华北地区冬季低温。
4.4 冷胁迫对不同品种樱花超氧化物歧化酶含量的影响
超氧化物歧化酶是非常重要的抗氧化酶,它能清除超氧根阴离子O2-,同时产生H2O2。超氧化物歧化酶含量越高说明植物耐寒性越强[26]。一定温度下的冷胁迫,会令植物体内的超氧化物歧化酶活性升高,有利于植株应对逆境。但若冷胁迫持续加强,抗氧化防御系统遭到破坏,超氧化物歧化酶的保护机制被打破,细胞受到损伤,酶活性下降[27]。4个樱花品种的超氧化物歧化酶含量均随着胁迫温度的降低呈现先上升后下降的趋势。‘大寒樱’在-10℃达到最高,而‘河津樱’、‘大渔樱’在-15℃达到最高,说明‘大寒樱’的抗寒性略弱。实际的田间观测中,试验地冬季最低气温在-15℃左右,在一定的越冬保护下‘河津樱’可以适应-15℃的温度,而‘大寒樱’则出现一定冻害,需要更进一步的越冬保护。孟诗原等[28]对5种卫矛属植物低温胁迫的生理响应研究表明,随着低温胁迫的加剧,超氧化物歧化酶含量呈现先上升后下降的趋势,不同品种达到峰值的温度不同。张迎辉研究了低温胁迫下福建山樱花的生理响应,发现随着温度的降低,植物体内的超氧化物歧化酶含量先上升后下降,与本研究得到的结果一致[29]。
对比丙二醛试验数据可以看出,‘大寒樱’的耐寒能力较其他3个品种有明显差异。结合田间观测情况,其对华北平原地区的冬季气候适应能力相对较弱,更不适宜在华北地区的山区栽植。冬季最低气温在-10℃以上的地区,在有效的越冬保护下可以种植该品种。
4.5 华北地区应用前景
本研究引种的‘河津樱’、‘大寒樱’、‘大渔樱’、‘修善寺寒樱’由寒绯樱与大岛樱杂交得到。由于上述4个品种中含有耐寒性不佳的寒绯樱基因,此前普遍认为不适宜种植于华北地区。本研究的开展弥补了华北地区引种上述品种的空白。通过研究表明,寒绯樱品系的樱花,因品种不同,温度抗逆性存在较大差异。通过筛选,部分品种可栽植于华北东南部平原地区。相较于夏季高温,华北地区的冬季低温更大程度地影响了上述品种樱花的生长表现,因此对于冬季低温的考量,应成为引种栽植时重点关注的方面。
上述4个品种樱花,花期早于目前华北地区常见的品种樱花,且花色呈玫粉色,深受游客喜爱。若能在园林造景中加以应用,可在早春呈现良好的景观效果。除上述4个品种外,寒绯樱与其他樱花杂交得到的品种,如‘椿寒樱’、‘飞寒樱’、‘红粉佳人’、‘オカメ’等,同样具有花期早、花色艳丽的特点,未来可进一步进行引种栽植试验。通过田间观测及生理生化指标测定,筛选出适宜华北地区栽植的品种。
猜你喜欢
今日农业(2022年1期)2022-11-16
中华诗词(2021年8期)2021-12-31
畅谈(2019年5期)2019-04-01
中华戏曲(2017年2期)2017-02-16
中国科技信息(2016年10期)2016-09-03
现代食品(2016年14期)2016-04-28
中国粮油学报(2014年7期)2014-02-06
浙江大学学报(医学版)(2013年6期)2013-12-09
