
利用ssr标记浅析云南登革热重点地区埃及伊蚊种群遗传特征
2021-03-24 05:55郑宇婷姜进勇周红宁马雅军
中国人兽共患病学报 2021年2期
郑宇婷,姜进勇,周红宁,马雅军
登革热(Dengue Fever)是登革病毒(Dengue virus)经蚊媒传播引起的急性虫媒传染病,主要在热带和亚热带流行[1]。1779年首次发现登革热病例[2],全球约25亿人有感染登革病毒的危险,每年约有1亿新感染病例,其中50万重症病例需住院治疗[3]。我国最早在1873年厦门首次报道,1995年后呈局部暴发、输入性流行的特征[2],2004年以来广东、福建、浙江等省份多次暴发本地疫情是我国重点防控区[4-6]。与云南省接壤的越南、缅甸和老挝等国家为登革热常年流行区[7]。云南省最早发现登革热病例是在1975年河口县[8], 2004年以来有散在输入病例报告,2008年在德宏州芒市、临沧市镇康县首次报道云南省登革热本地病例[9-10], 2013年景洪市暴发登革热本地疫情[10],此后我省西双版纳州景洪市、勐腊县、德宏州瑞丽市、临沧市耿马县等多次暴发流行,且流行日趋严重,登革热已经成为我省边境地区重要的虫媒传染病,严重危害边境地区人群健康。
埃及伊蚊(Aedesaegypti)是登革热的重要传播媒介,其传播能力远高于白纹伊蚊。埃及伊蚊为云南省的入侵蚊虫,2002年首次在瑞丽市发现,其后10余年间分布范围不断扩大,德宏州的芒市、陇川和盈江,西双版纳的景洪、勐腊、勐海,临沧市的耿马等3个州(市)、8个边境县(市)均有埃及伊蚊的分布[11-12]。埃及伊蚊的入侵加剧了云南省登革热的暴发和流行风险。媒介控制是登革热防控的重要措施,研究埃及伊蚊的遗传特性,对登革热防控具有重要意义。
微卫星(Simple Sequence Repeat,SSR)一般是由2~6个碱基为核心组成首尾相连的串联重复序列。DNA的滑动复制导致微卫星DNA长度多态性的产生,在埃及伊蚊种群特征的研究中,利用对核苷酸重复单位数目差异的研究来分析遗传相关的各项指标。微卫星因共显性遗传、高多态性和重复性好等特点,是研究蚊虫群体遗传结构理想的分子标记之一[13],本研究采集云南省5个登革热重点县的埃及伊蚊,筛选12个多态微卫星位点,通过微卫星DNA标记分析云南省5个不同地理来源的埃及伊蚊种群的亲缘关系群遗传多样性和遗传结构特征,对有效防制埃及伊蚊、科学防控登革热提供科学依据。
1 材料与方法
1.1材料来源 依据埃及伊蚊入侵、定值和扩散的程度,选择云南省德宏州瑞丽市、西双版纳州景洪市、勐腊县、勐海县、临沧市耿马县作为调查地点,在埃及伊蚊蚊虫密度高峰季节,采集花瓶、轮胎、水桶、废弃瓶以及其他等容器中孳生的伊蚊幼虫,带回实验室根据不同的孳生容器分开饲养至羽化,形态学鉴定虫种[14],并将鉴定出的埃及伊蚊保存于冻存管分装,单管单只。每个点采集根据采集方位和孳生容器随机抽取29~30只[15]埃及伊蚊成蚊样本,确保采集样本覆盖每种孳生容器。低温冰箱保存待后续实验。
1.2实验方法 样本单管单只成蚊,采用DNA提取试剂盒[TIANamp Genomic DNA Kit(血液/细胞/组织基因组DNA提取试剂盒)(离心柱型)(含蛋白酶K)],按照试剂盒操作说明书提取蚊虫组DNA,并用超微量核酸分析仪(杭州奥盛仪器有限公司,Nano-200)检测浓度和纯度,通过查阅文献找出埃及伊蚊微卫星位点的侧翼序列[16]设计13对引物(表1)。
对每个样本进行13对引物的PCR扩增,反应体系共25 μL:10*Ex Taq buffer 2.5 μL,DNTP (2.5 mmol/L) 2 μL,MIX primer 1 μL(MIX primer 为正、反向引物等量混合),DNA模板1 μL,Ex Taq 0.2 μL,H2O 18.3 μL。反应条件为:96 ℃ 5 m, 96 ℃ 20 s, 53~58 ℃ 30 s,72 ℃ 30 s,10个循环;96 ℃ 20 s, 52 ℃ 30 s,72 ℃30 s,30个循环;72 ℃ 5 m。扩增产物毛细管电泳后得到原始数据(华大基因公司完成)。
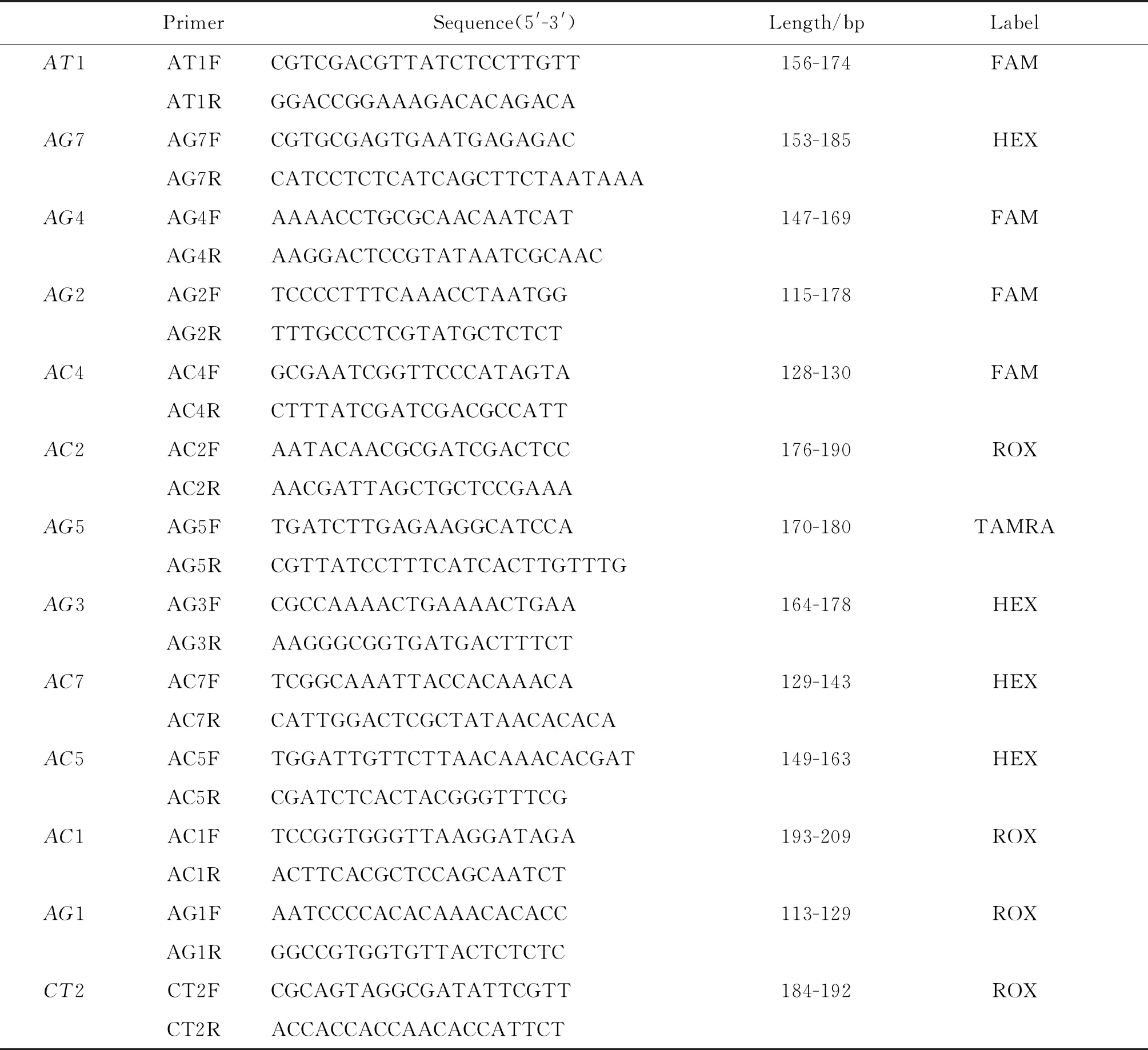
表1 埃及伊蚊13个微卫星位点引物的相关信息
1.3数据统计与分析 采用GeneMaker(version2.20)对原始数据进行分析处理,利用GenAIEx 6.503计算各基因座等位基因数(number of allele, Na)、有效等位基因数(effective number of allele, Ne)、观测杂合度(observed heterozygosity, Ho)、期望杂合度(expected heterozygosity, He)、群体内近交系数(population inbreeding coefficient, Fis)、遗传分化系数(genetic differentiation coefficient, Fst)、基因流(number of migrants per generation, Nm)以及Nei氏标准遗传距离(Nei’s standard genetic distance)、遗传相似系数(Nei’s standard genetic Identity)等指标,并根据遗传分化和地理距离进行mantel test分析。使用PIC-CALC软件计算多态信息含量(polymorphism information content,PIC)。利用Arlequin3.5.1计算种群间成对固定指数Fst。
2 结 果
2.1蚊虫采集结果 2017年6月1日-24日分别在瑞丽、勐腊、景洪、勐海、耿马采集伊蚊幼虫,并在显微镜下初步判定,收集埃及伊蚊幼虫并饲养至成蚊,共收集234个有伊蚊幼虫的容器,经鉴定饲养收集的埃及伊蚊有603只(表2)。
不同孳生容器中随机选取1~10只,总149只开展实验(表3)。

表2 埃及伊蚊采样信息表

表3 选取埃及伊蚊实验样本信息表
2.2毛细管电泳扩增的结果 样本DNA提取经超微量核酸提取仪检测后浓度在16.51~77.88 ng/μL,经过毛细管电泳扩增以后共得到1 750个有效片段用于埃及伊蚊的遗传特征分析,其中AG7、AC4、AC2引物扩增效果最理想,全部扩增出有效片段;AC1引物扩增效果最差。扩增产物大小为91~197 bp之间。用GeneMaker(version2.20)对原始数据进行分析处理,检测数据和结果较多,分别选取2个位点的2个样本的系列图峰作为示例描述。图1分别是样本MH066位点AG1上的杂合子、样本RLCQ23位点AC4上的纯合子。

(MH066AG1为杂合子,大小为107和114;RLCQ23AC4为纯合子,大小为115和115)
2.3微卫星遗传多样性 CT2位点未扩增出目的片段,其余12对微卫星标记对5个不同地理位置:景洪、勐腊、勐海、瑞丽、耿马的埃及伊蚊自然种群进行扩增,共检测到216个等位基因,每个位点的等位基因个数为10~31个,所有位点的平均等位基因数为18,有效等位基因为4.653个。其中,位点AG7和AG2的等位基因为31个,等位基因数量最多;位点AG3的等位基因为10个,数量最少。观测杂合度为0.548~0.910之间,平均值为0.711;期望杂合度为0.623~0.827,平均值为0.728。12个多态位点的多态信息含量范围为0.615~0.881,平均PIC 为0.778。其中AG1位点标记PIC指数最高为0.881,AC1位点的标记PIC指数最低为0.615(表4)。
从种群角度分析,平均每个种群的等位基因数在7.750~10.583之间,等位基因数最多的是耿马,最少的是勐腊(表5)。5个自然种群平均观测杂合度分布范围是0.661(JH)~0.741(ML),平均期望杂合度的分布范围在0.629(ML)~0.774(RL)之间。PIC指数最高是RL种群(0.750),最低是ML种群(0.588)(表5)。
2.4遗传分化系数及基因流 12个微卫星位点的群体内近交系数(Fis)有8个位点的杂合子过剩(Fis>0),4个位点的杂合子缺失(Fis<0),Fis范围在0.220~0.207之间,平均值为0.028,AC1和AG2位点的Fis和Fit值最高,分别为0.207、0.137和0.260、0.267。说明这2个位点群体内近交程度较其他位点高(表6)。
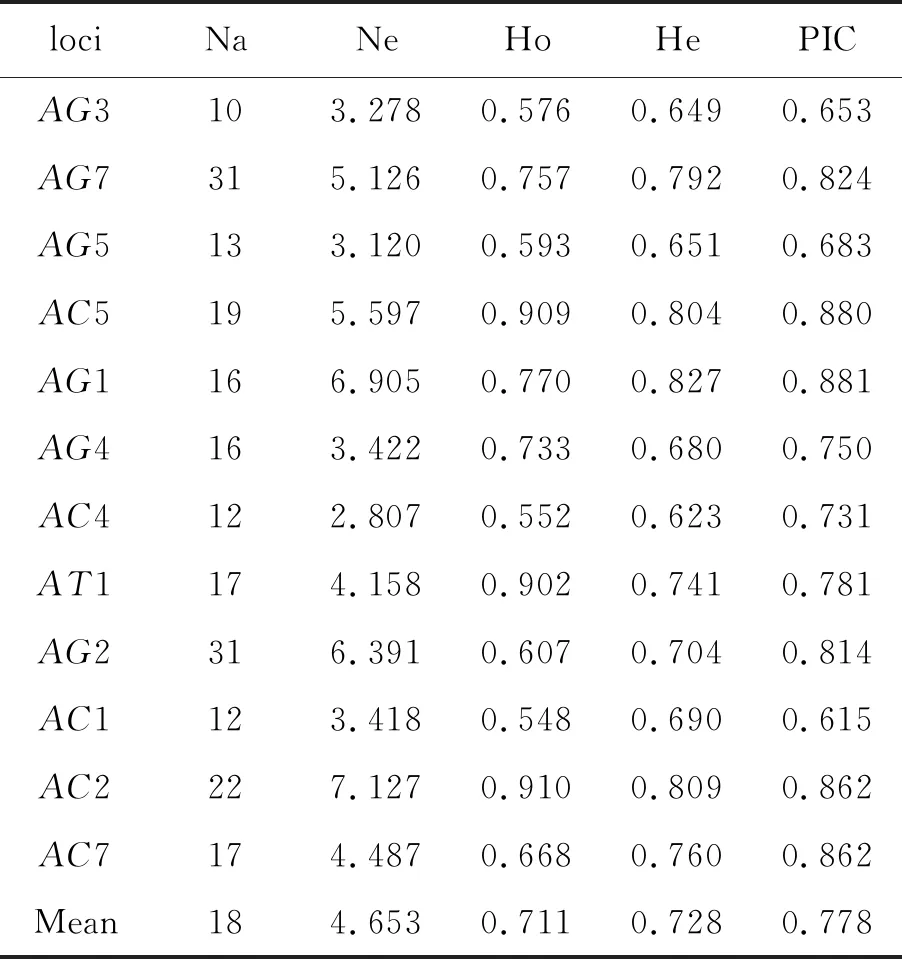
表4 12个微卫星位点的总遗传参数
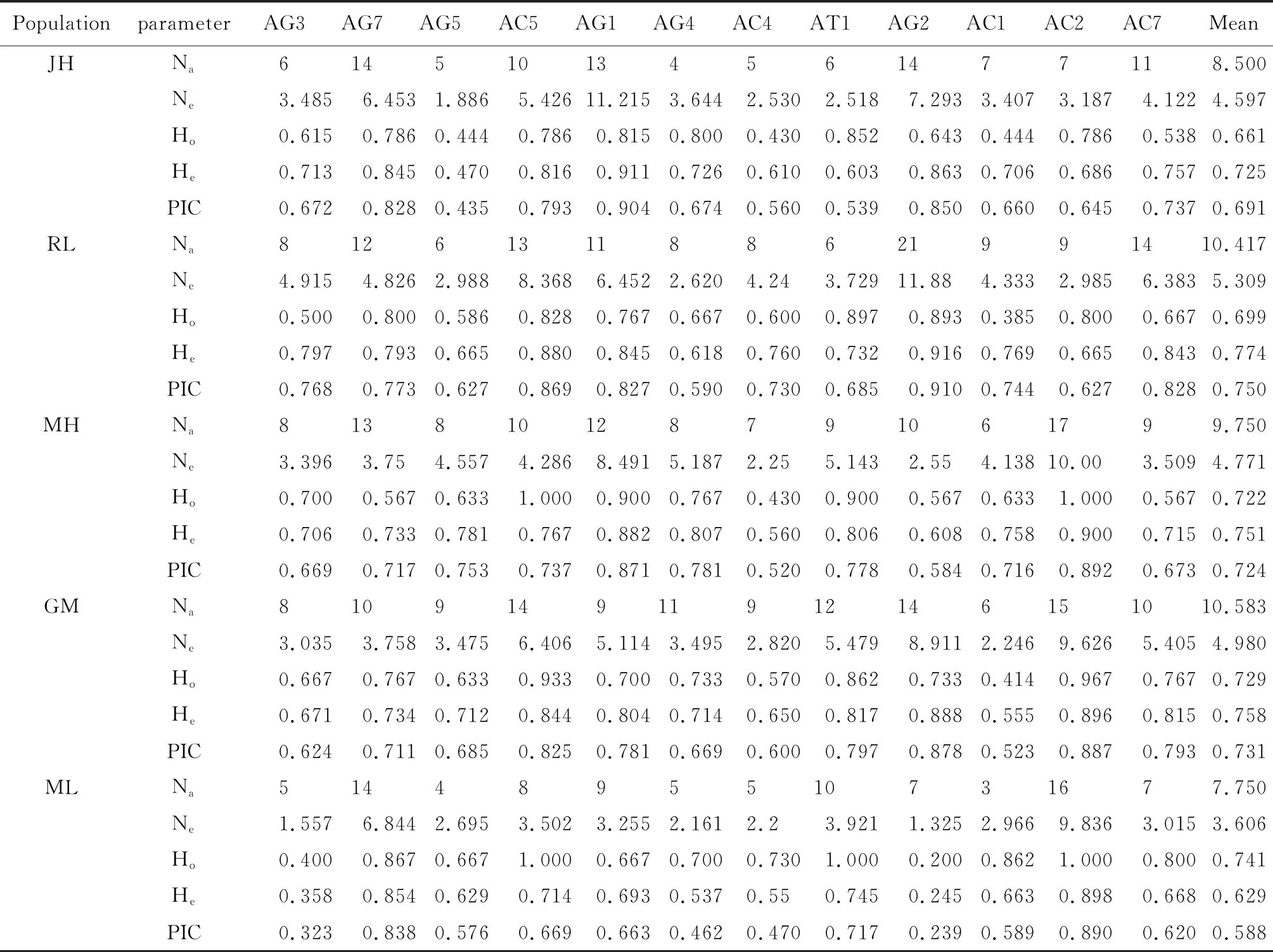
表5 云南省5个埃及伊蚊自然种群12个微卫星位点的遗传特征数值
按照Balloux等[17]的遗传分化的标准(低:0.15~0.25,中:0.05~0.15,高:0~0.05),本研究群体间近交系数(Fst)平均值为0.078,为中度遗传分化。
云南5个埃及伊蚊群体Nm均值为3.321,AG5位点上的基因流最大(5.210),AG2位点上的基因流最小(1.414)。所有位点的基因流均大于1(表6)。
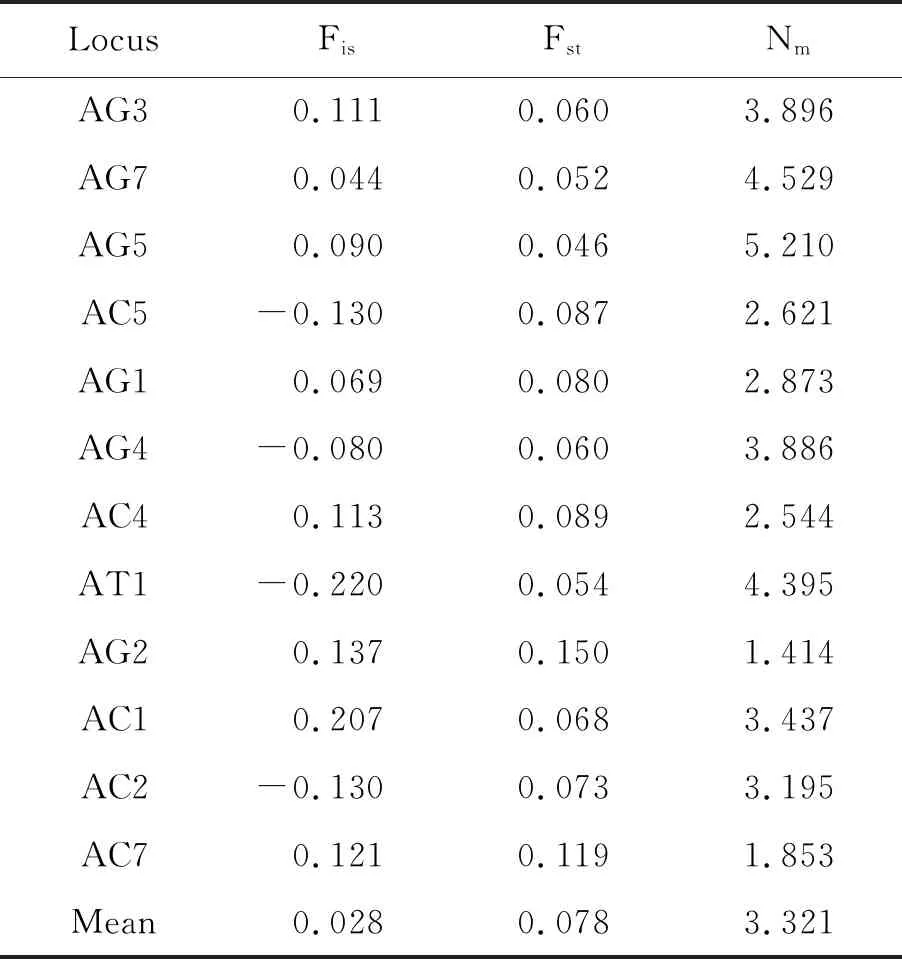
表6 云南5个埃及伊蚊种群12个微卫星位点上的遗传分化系数和基因流的情况
应用GenALEx软件中的Mantel Test检验埃及伊蚊种群间遗传分化和地理距离的相关性,地理距离(表7)由采样点经纬度计算得出,未发现相关性(y=4E-07x+0.251 1,R2=0.263 5,P=0.100)。

表7 云南5个埃及伊蚊自然种群间地理距离(对角线之上)与遗传分化(对角线之下)矩阵表
2.5云南5个自然种群间遗传相似性指数和遗传距离 Nei’s标准遗传距离在0.162~0.527之间,对应的遗传相似度在0.590~0.850之间(表8)。

表8 5个埃及伊蚊种群Nei’s标准遗传距离(对角线之下)与遗传相似系数(对角线之上)
勐腊与勐海群体的遗传相似度最高为0.850,遗传距离为0.162;勐腊与瑞丽群体的遗传相似度最低为0.590,遗传距离为0.527。
3 讨 论
5个埃及伊蚊自然种群的遗传多样性、生物多样性研究中遗传多样性的研究尤为重要,等位基因数、观测杂合度、期望杂合度和多态信息量等参数用于反映遗传多样性水平的重要指标,其值越大,说明基因丰富度越高、生物对环境的适应能力越强[18]。
本研究利用12对微卫星分子标记对5个埃及伊蚊自然种群进行遗传多样性分析,12个位点平均Ho为0.711,平均He为0.728,平均PIC为0.778,平均每个种群的等位基因数在7.75~10.583之间,表明各种群遗传多样性较高,与石清明、刘蓬勃等的研究结果项目一致[19-20]。景洪、瑞丽、勐海、耿马、勐腊的平均Ho分别为0.661、0.699、0.722、0.729、0.741;平均He分别为0.725、0.774、0.751、0.758、0.629;平均PIC分别为0.691、0.750、0.724、0.731、0.588,杂合度代表基因多样度,在不考虑样本数量影响的情况下,反应群体在多个基因位点的遗传变异程度。本研究中景洪、瑞丽、耿马、勐海种群平均期望杂合度高于平均观测杂合度,意味着种群内可能存在近交;勐腊种群平均观测杂合度高于平均期望杂合度,意味着种群可能有大量外来个体的补充,或者可能经历了瓶颈效应或奠基者效应[19]。参照Botstein等提出的衡量基因变异程度高低的PIC指标,本研究5个种群PIC都大于0.5(平均为0.697),为高度多态。5个群体中瑞丽的PIC最高,表明呈现高度多态。
5个埃及伊蚊自然种群的遗传和亲缘关系群体内近交系数,其值越接近0,基因的分布越接近平衡,值为负时提示杂合子缺失,值为正时存在杂合子过剩[21]。基因在群体中的流动称为基因流,Nm越大说明群体越均匀[22],大于1则能够抑制群体间的分化[23]。
各种群的近交系数中勐腊Fis值为负,表示与其他种群近交程度很低,其他4个种群为正,说明这些种群中存在着不同程度的近交。5个自然种群的样本成对固定指数Fst均在0.057~0.148之间,表示种群间中度分化。基因流Nm越大,群体间的相似性越大。5个自然种群12个微卫星位点的平均Nm为3.321,种群基因流较高,暗示5个群体间遗传交流水平高,能够抵制遗传漂变作用和防止种群分化的发生。与刘蓬勃[20]、石清明等[19]的研究结果一致。
遗传距离和相似度都是用来分析群体之间的亲缘关系,5个群体之间勐腊和瑞丽的Nei’s遗传距离最大,遗传相似度最低,表明这2个群体的亲缘关系最远;勐腊和勐海的Nei’s遗传距离最小,遗传相似度最高,说明这2个群体的亲缘关系最近。经计算5个自然种群的地理距离和遗传分化之间无显著相关性。
本研究采用微卫星标记技术对云南省5个埃及伊蚊自然种群进行遗传多样性分析,多样性指标PIC为0.588~0.750,呈现高度多态性,近交系数为0.028;遗传分化系数为0.078,基因流均值为3.321,表明群体间遗传交流水平高,存在一定程度的遗传分化。5个群体间的遗传距离为0.162~0.527之间,勐腊和勐海群体遗传距离最小,亲缘关系最近,勐腊和瑞丽的群体遗传距离最远,亲缘关系最远。景洪、瑞丽、勐海、耿马的埃及伊蚊种群存在着不同程度的近交,勐腊的埃及伊蚊种群可能经历过瓶颈效应或者有大量外来个体补充,还需要进一步确定。
利益冲突:无
猜你喜欢
烟台大学学报(自然科学与工程版)(2022年3期)2022-06-30
数学物理学报(2022年1期)2022-03-16
现代临床医学(2022年1期)2022-02-12
化工管理(2021年7期)2021-05-13
中国人兽共患病学报(2021年7期)2021-03-28
寄生虫与医学昆虫学报(2021年3期)2021-02-10
广东医科大学学报(2020年4期)2020-08-24
四川动物(2017年6期)2017-12-12
四川动物(2017年4期)2017-07-31
中南林业科技大学学报(2015年6期)2015-12-20
