
亚热带城市环境下桑寄生植物的7年扩散动态
2021-03-17 06:43
中南林业科技大学学报 2021年2期
(四川大学 a.生命科学学院;b.教育部生物资源和生态环境重点实验室,四川 成都 610065)
桑寄生植物(Mistletoe)是一类能进行光合作用的半寄生植物,不能独立生长,一般寄生于木本植物的枝干上[1]。桑寄生植物主要包括桑寄生科Loranthaceae、槲寄生科Viscaceae、羽毛果科Misodendraceae 和檀香科Santalaceae 植物[2]。其生活史较为复杂,大部分种类依靠鸟类来传播种子[3-4],少数种类还可以通过小型哺乳动物传播种子[5]。桑寄生植物种子通过动物的传播到达其寄主植株上,在适宜的条件下种子萌发,之后与寄主木质部结合形成功能吸器,进而从寄主个体上获得水分和养分[6-7]。桑寄生植物的定居一般会对寄主植物造成不同程度的危害,它们往往具有较强的蒸腾作用,会给寄主植物造成强烈的干旱胁迫,导致寄主植物叶片气孔震荡关闭[8],降低寄主植物的光合速率[9-10],甚至会导致寄主植物死亡[11]。
特定区域内,被桑寄生植物所寄生的木本植物种类称之为寄主范围(Host range),寄主种类范围之外的植物种类被称为潜在寄主(Potential host)[12-13]。不同桑寄生植物的寄主各异,多数桑寄生植物可以寄生于多种寄主植物上,少数寄生于单属或单种寄主植物上[14]。寄生植物能否成功寄生,与其寄主植物的个体生物学特征密切相关[15-17]。比如,木本植物个体越高大,越容易被桑寄生植物所寄生[17],并且单株寄主树木上的桑寄生植物个体数量越多[18]。寄主所在群落的树种多样性也影响着桑寄生植物的定居和扩散[15,19-21],桑寄生植物多发生于乔木物种多样性低且受干扰的破碎化生境中[22-23]。另外有假说认为:潜在寄主物种的多度越多,越容易成为寄主植物[12,24-26]。目前,对于寄主特性和寄主群落特征的研究较为充分[6,9,13,21,25];对于桑寄生植物能否成功寄生在某种寄主植物上与该种寄主植物的多度有关的研究还较少,且大多建立在单次调查的基础上[12,25],无法从时间动态上来证实该假说的普遍性。潜在寄主植物多度是否是其成为桑寄生植物的寄主植物的限制性条件,需要从时间动态上来进行验证。
一般而言,桑寄生植物种群数量越大,寄主受到的危害越为严重[27]。桑寄生植物的种群大小受其聚集效应的影响[23],原因在于其种子传播者(鸟类或小型兽类)偏好于在寄生感染率高的生境中活动,导致该区域内的寄生感染率进一步增加[23,28-29]。桑寄生植物对其寄主造成的危害会因这种聚集效应而加剧,寄生植物的种群数量在局部可迅速增多,从而可对寄主植物造成严重危害,甚至死亡[11,27-28,30]。寄主个体死亡后,其上的桑寄生植物也会随之死亡。然而,对于特定区域内的桑寄生植物种群数量会因为寄主的死亡而减少还是因为其聚集效应而增加,尚需进一步的探索。
本研究以成都市区55.4 hm2内的桑寄生植物及其寄主为研究对象,分别于2012 和2019年对寄生植物与寄主植物的种类、数量及其空间分布进行了调查和对比分析,以期探明桑寄生植物的寄主种类、寄生感染率、寄生强度的动态变化,初步揭示桑寄生植物的种群扩散机制。另外,我们拟验证一个假说:桑寄生植物的寄主选择与其潜在寄主植物的多度呈正关联关系,即,桑寄生植物潜在寄主植物的多度越大,其越容易成为桑寄生植物的新寄主种类[12,15]。通过研究,可丰富桑寄生植物的种群生态学研究,为桑寄生植物的生态防控提供一定的理论支撑。
1 研究方法
1.1 研究区概况
四川省成都市位于四川盆地西部,以平原为主。成都市属于亚热带季风性湿润气候,年平均气温为16.2℃,气温最高月份为8月,平均气温为25.0℃;气温最低月份为1月,平均气温为5.6℃。降雨充沛,年均降水量约980 mm。研究区域位于四川省成都市四川大学望江校区内,该区域主要植被类型以草地、绿篱、行道树、小树林以及散落在房前屋后的自然植被。草地主要由早熟禾Poa annua和黑麦草Lolium perenne组成。乔木树种主要有榕树Ficus microcarpa、银杏Ginkgo biloba、银木Cinnamomum septentrionale、二球悬铃木Platanus acerifolia等。灌木种类主要有檵木Loropetalum chinense、小蜡Ligustrum sinense、蜡梅Chimonanthus praecox、多种苹果属Malusspp.植物、梅Armeniaca mume、紫荆Cercis chinensis、夹竹桃Nerium indicum等。校园内还种植着很多亚热带常见的常绿树种,如:黄葛树Ficus virens、阴香Cinnamomum japonicum、木犀Osmanthus fragrans、女贞Ligustrum lucidum等。校园内的大部分乔木种类为人工种植,只有少量种类是乡土树种,如构树Broussonetia papyrifera、桑Morus alba等。
依托大量种植的树木,研究区域内鸟类众多,分布着与桑寄生种子传播关系较为密切的鸟类,如:红胸啄花鸟Dicaeum ignipectus、白头鹎Pycnonotus sinensis、黑头蜡嘴雀Eophona personata、黄腹山雀Pardaliparus venustulus、淡黄腰柳莺Phylloscopus chloronotus、红胁绣眼鸟Zosterops erythropleurus和红头长尾山雀Aegithalos concinnus等[31]。该区域内桑寄生植物共2 种,分别为毛叶钝果寄生Taxillus nigrans和红花寄生Scurrula parasitica。
1.2 调查方法
2012年9—12月和2019年1—6月,分别对四川大学望江区55.4 hm2面积内的木本植物桑寄生状况进行了两次调查。以单株树木为单位进行调查,借助望远镜记录单株寄主树木上的桑寄生种类和数量(寄生强度)。记录寄主植物的种类和数量。
1.3 分析方法
以不同寄主个体上寄生植物的数量构建统计量。通过方差均值比法(A=S2/X)来判定桑寄生种群空间分布格局的类型[32-34]。当A=1 时,种群为随机分布;当A>1 时,种群为集群分布;当A<1时,种群为均匀分布。显著性水平α=0.05。
寄生感染率(Parasitic prevalence)是指研究区域内寄主植物株数占该种植物总株数的比例,计算公式为:寄生感染率=(寄主植物株数/该种植物总数量)×100%。采用Fisher 精确检验(0.95置信水平,P<0.05 为差异显著),对比2012年和2019年各寄主的寄生感染率之间的显著性。当P<0.05 时,表明不同年份间寄生感染率之间差异显著,否则差异不显著。采用独立样本t检验,比较2012年和2019年各寄主的寄生强度变化,显著性水平α=0.05,当P<0.05 时,表明同一寄主植物在2012年和2019年寄生强度差异显著,否则差异不显著。数据统计分析在SPSS 20.0 软件中进行,采用Origin 2017 软件作图。
2 结果与分析
2.1 不同年份寄主植物多样性
2012年研究区域内共调查到木本植物5 343株,隶属于41 科70 属86 种。其中991 株被桑寄生植物寄生,寄主植物共25 科37 属41 种。2019年共调查到木本植物5 490 株,隶属于42 科64 属78 种,其中1 294 株被桑寄生植物寄生,寄主植物共30 科42 属48 种。两次调查共记录寄主植物32 科47 属54 种,两次调查均记录桑寄生植物2 种,为毛叶钝果寄生和红花寄生(表1)。
相比2012年,2019年样地中新增寄主树种13个,分别为冬青卫矛、复羽叶栾树、黑壳楠、胡桃、毛泡桐、木芙蓉、枇杷、石榴、阴香、樟、梓、紫荆和檵木。在新增的13 种寄主植物中,毛泡桐和檵木未出现在2012年的调查记录中。调查区域内寄主树种减少了6 种,分别为李、榆树、八角枫、红花羊蹄甲和桃。桃和红花羊蹄甲的样本量在2019年小于20 株,榆树、李、乌柿和八角枫在2019年度调查中均未在调查区域中发现(表2)。
2012年,桑寄生植物的潜在寄主树种共45 种,其中研究区域内,个体数量≥20 株潜在寄主树种共10 种,<20 株的潜在寄主树种共35 种;寄主树种共41 种,个体数量≥20 株的寄主树种共31种,<20 株的寄主树种共10 种。个体数量≥20株的寄主种类显著多于个体数量<20 株寄主的种类(P<0.000 1),即寄主的树种主要集中于个体数量≥20 株的树种上(表3)。
2019年桑寄生植物的潜在寄主树种共30 种,其中研究区域内,个体数量≥20 株潜在寄主树种共7 种,<20 株的潜在寄主树种共23 种;寄主树种共48种,个体数量≥20株寄主树种共32种,<20 株的寄主树种共16 种。与2012年类似,个体数量≥20 株的寄主种类亦显著多于个体数量<20株寄主的种类(P=0.000 3),即寄主的树种主要集中于个体数量≥20 株的树种上(表3)。
2.2 桑寄生植物的扩散动态变化
2.2.1 桑寄生植物在寄主个体上的分布格局
2012年结果显示,桑寄生植物在单株寄主植物上的数量在1~64 之间;2019年的结果显示,桑寄生植物在单株寄主植物上的数量在1~63 之间,两次种群调查的分布均呈偏峰态分布。无论不论是2012年还是2019年,单株寄主上仅1 株桑寄生植物的寄主个体数量最多(2012年285 株,2019年327 株);单株寄主树木上小于3 株寄生植物的个体分别占所有被寄生个体的55.6%(2012)和50.9%(2019);单株寄主植物上的桑寄生植物数量的中位数为3;单株寄主树木上的桑寄生植物种群数量越大,这类寄主个体的数量越少(图1)。
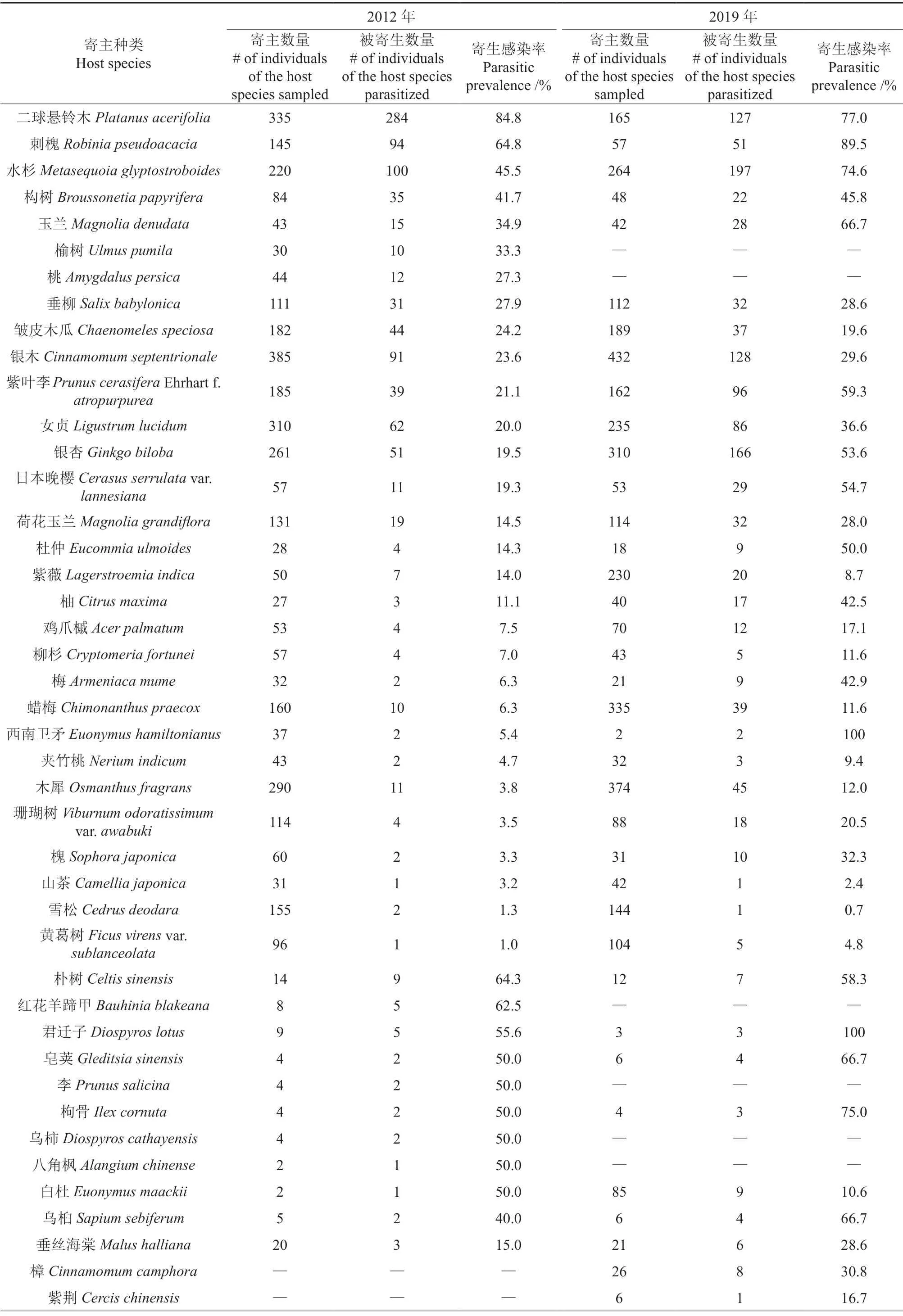
表1 不同年份寄主植物种类、样本量和寄生感染率调查记录†Table 1 Host species,sample size,and parasitic prevalence of the mistletoe parasitism in 2012 and 2019

续表1Continuation of table 1

表2 2012 和2019年寄主种类、数量和寄生感染率的变化Table 2 The changes of host species,individuals,and parasitic prevalence in 2012 and 2019

表3 2012 和2019年寄主和潜在寄主的样本Fisher 精确检验Table 3 Fisher’s exact test of the sample sizes on host species and potential host species in 2012 and 2019
2012年寄生强度在寄主个体上分布格局(A2012=2 156.4)显著大于1,2019年寄生强度在寄主个体上分布格局(A2019=2 455.1)明显大于1。在置信度为95%、自由度为45 和48 的t值为2.021时,2012年和2019年桑寄生植物的寄生强度在寄主个体上的分布格局均为显著的聚集分布(P<0.000 1)(表4)。
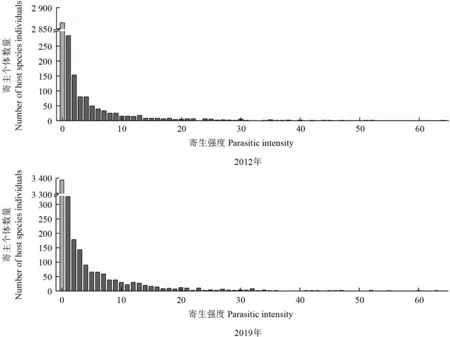
图1 2012 和2019年桑寄生寄主植物的寄生强度频数分布Fig.1 The distribution of the parasitic intensity for all host species individuals in 2012 and 2019

表4 2012 和2019年桑寄生植物寄生强度的空间分布格局Table 4 Distribution patterns of the mistletoe parasitic intensity in 2012 and 2019
2.2.2 不同寄主树种上的寄生感染率变化
对比2012年和2019年桑寄生感染率调查结果,桑寄生植物寄主的数量从991 株增加到1 294 株,寄主数量增加了303 株。2012年总体寄生感染率为18.1%,2019年总体寄生感染率为27.2%,2019年总体寄生感染率较2012年显著增加(P<0.05)。
2012年寄主和2019年寄主相同的树种共35种,对比不同年份35 种寄主植物的寄生感染率(图2)。结果显示:刺槐、水杉、玉兰、紫叶李、女贞、银杏、日本晚樱、荷花玉兰、杜仲、柚、梅、木犀、珊瑚树和槐和西南卫矛共15 种寄主树种的寄生感染率相较于2012年显著增加(P<0.05);构树、垂柳、银木、鸡爪槭、柳杉、蜡梅、夹竹桃、黄葛树、君迁子、皂荚、枸骨、乌桕和垂丝海棠共13 种寄主树种的寄生感染率较2012年趋于增加(P>0.05);二球悬铃木和皱皮木瓜的寄生感染率显著下降(P<0.05);紫薇、山茶、雪松、朴树和白杜共5 种寄主树种的寄生感染率较2012年趋于降低(P>0.05)。
2.2.3 不同寄主树种上的寄生强度变化
二球悬铃木、刺槐、银杏、水杉、荷花玉兰、女贞、君迁子、木犀、银木、日本晚樱和柚共11种寄主树种的寄生强度较2012年显著增加(P<0.05)。经过7 a 的扩散,二球悬铃木和刺槐依旧是平均寄生强度最高的树种,平均寄生强度分别为17.32 和15.14,寄生强度增加近一倍(图3A)。君迁子和日本晚樱的平均寄生强度增幅最大,寄生强度增加3 倍以上(图3A)。皂荚、乌桕、槐、梅、黄葛树、山茶、雪松和垂丝海棠共8 种寄主植物在研究区域内样本量较少,平均寄生强度相较于2012年均趋于上升(P>0.05)(图3C)。
紫薇和珊瑚树的平均寄生强度显著下降(P<0.05),杜仲、皱皮木瓜和西南卫矛的平均寄生强度也趋于下降(P>0.05)。此外,还有4 种小样本的寄主植物的平均寄生强度下降,分别是白杜、夹竹桃、枸骨和蜡梅。
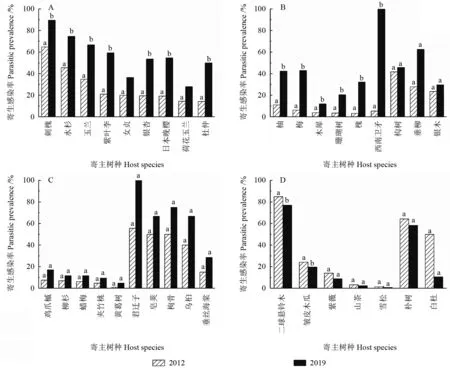
图2 2012 和2019年各寄主物种的寄生感染率比较Fig.2 Comparison of parasitic prevalence of each host species between 2012 and 2019
3 讨 论
3.1 桑寄生植物的寄主种类变化
研究表明,桑寄生植物对寄主植物的选择是动态变化的,这种动态性体现在时间和空间上是可变的,并取决于时间和空间上任何可利用的机会[25]。本研究中,七年间桑寄生植物的寄主种类增加了13 种,减少了6 种,寄主种类由41 种增加到了48 种。这说明增加或减少的19 种寄主植物能否被桑寄生植物成功定居在时间上是不稳定的,七年间部分寄主植物上的桑寄生消失了,成为了非寄主植物,而部分潜在寄主植物则成为了长有桑寄生的寄主植物。

图3 2012 和2019年寄主物种的寄生强度比较Fig.3 Comparison of parasitic intensity of each host species in 2012 and 2019
在2012年的研究结果中,桑寄生植物多寄生于个体多度较大的寄主树种上,这个结论与Roxburgh 等[35]在南非对桑寄生植物Erianthemum dregei的研究结论一致,桑寄生植物潜在寄主植物的多度越大,其越容易成为其新寄主种类[12,35]。研究区域内,桑寄生植物多定居在种群数量较大的木本植物上,这些木本植物相比较于种群数量较小的乔木或灌木有更多的机会接触到桑寄生植物的种子传播者[12]。随着种子传播者对种群数量较大的寄主植物的高频率造访,桑寄生植物的种子有更多的机会扩散这些种群数量较大的物种上,而后在适宜的环境下萌发,使其成为新的寄主物种。2019年的调查结果显示,新增的13 种寄主植物中,5 种寄主植物的样本数量大于20 株。同时,所有的寄主种类中,个体数量≥20 株种类共32种,<20 株的仅有16 种,该结果进一步说明了桑寄生植物偏好寄生于木本植物多度较高的寄主树木上。究其本质,这可能是由于多度较高的种类相较于多度较少的种类能够有更多的机会接触到桑寄生植物的种子[12,24-26]。例如:研究区域阴香和黑壳楠的种群数量均超过了200 株,这两种寄主植物在2012年并未发现有寄生植物成功定居。由于这两种寄主植物种群数量大,桑寄生植物的种子传播者在种子传播的过程中可能造访这两种植物的机会更多,使得桑寄生植物有更多的机会能在这两种寄主植物上萌发、生长并最终定居,所以在2019年的复查中记录到阴香和黑壳楠均由潜在寄主成为了寄主。
本研究结果再次证实了桑寄生植物潜在寄主的多度越大,就越有可能成为桑寄生植物新寄主的假说[12,18]。但也有例外,例如,研究区域内,榕树和银桦Grevillea robusta作为行道树被广泛栽植,但2012 和2019年均未发现这两种树木上有桑寄生植物定居,说明这两个潜在寄主种类很可能属于非寄主种类,即桑寄生植物无法成功定居的种类。关于桑寄生植物寄主选择的机制较为复杂[6]。对于本研究中的类似榕树和银桦的非寄主种类,我们认为桑寄生无法成功定居的原因可能为:1)时间跨度不够,在未来这些潜在寄主植物可能成为寄主植物;2)这些潜在寄主植物可能对桑寄生植物具有生理阻隔作用[15],桑寄生植物无法在其机体上成功定居。这种生理阻隔作用目前尚不清楚其机理[6],还需要进一步深入。
3.2 桑寄生植物种群动态
本研究中,桑寄生植物的寄主树木由2012年的991 株增加到了2019年的1 294 株,15 种寄主植物的寄生感染率显著增加。经过7 a 的繁殖和扩散,研究区域内的桑寄生植物种群数量进一步增多。11 种寄主树种的平均寄生强度相较于2012年显著增加,表明桑寄生植物在这11 种寄主树木上感染了更多的寄主个体。这种在城市中寄生植物感染率和寄生强度的增加可能有以下三方面原因:
1)不同年份的分布频度格局都表明桑寄生植物的寄生强度在不同寄主个体上呈现集群分布的分布模式。这种集群分布的模式体现在绝大多数寄主个体被少量的桑寄生植物寄生,仅有很少一部分寄主个体被严重寄生。该研究结果与巴西[15]和波兰[18]的研究类似,说明桑寄生植物在区域尺度上确实是集群分布的,跟时间动态无关。桑寄生植物的聚集分布模式会增大其寄主的寄生感染率。
2)在城市绿化环境中,树木一般种植得较为集中,空间分布和物种多样性与人类活动密切相关。当桑寄生植物克服寄主植物的抗寄生性成功定居后[36],其强大的果实生产力可以生产出更多果实吸引种子传播者取食,然后通过种子传播者扩散到更多的同种寄主树种上,种子传播者为桑寄生植物的聚集分布起到了正反馈的作用[37]。刺槐、水杉、玉兰、紫叶李、女贞、银杏等寄主植物多作为行道树成行栽培,相互株距很近,这种种植模式可能会产生对寄生植物扩散和生长有利的边缘效应。即在破碎化森林的林缘且较为高大的寄主树木上,其寄生植物密度较大且生长较好[21,23]。此外,在稳定的环境中,寄生强度的增加与桑寄生植物种子数量的多少呈显著正相关[28]。
3)城市污染加剧可能是桑寄生植物种群增长的原因之一。桑寄生植物的含氮量一般高于其寄主植物,可获得性的氮元素是其成功定居的关键性制约因素[38]。有研究发现,大气氮沉降的增加可导致部分木本植物的叶片含氮量增加[39]。Kołodziejek 等[18]对波兰罗兹市(Lodz)桑寄生的研究发现白果槲寄生Viscum album多发生在城市二氧化氮污染较为严重的区域。近年来,成都市城市化进程发展迅速,空气污染物中含氮化合物的排放量趋于增加[40],使得大气氮沉降趋于增加。因此,假设桑寄生植物在城市污染环境下可能更易从其寄主上获得氮元素,从而有利于其繁殖和扩散,这可能是研究区域内7年间桑寄生植物种群扩张的原因之一。
4 结 论
为揭示桑寄生植物的种群扩散机制,本研究以桑寄生植物及其寄主为研究对象,分别于2012和2019年,在成都市区55.4 hm2范围内对寄生植物与寄主植物的种类、数量及其分布进行调查,分析了两种桑寄生植物寄主种类、寄生感染率和寄生强度的动态。研究区域内2012年有寄主植物41 种,寄主个体991 株;2019年有寄主植物48 种,寄主个体1 294 株。在2012 和2019年,桑寄生植物的寄生强度在寄主植物上的分布格局均呈聚集分布。相比2012年,2019年新增寄主植物13 种,减少5 种。2019年总体寄生感染率(27.2%)显著大于2012年的总体寄生感染率(18.1%)(P<0.05)。2019年的总体寄生强度(1 294)显著大于2012年的总体寄生强度(991)(P<0.05)。通过Fisher 精确检验,我们检验了假说(桑寄生植物的寄主选择与其潜在寄主植物的多度呈正关联关系)的正确性,并发现在2012 和2019年,个体数量≥20 株的寄主种类显著均多于个体数量<20 株寄主的种类(P<0.05),即寄主的树种主要集中于个体数量≥20 株的树种上。桑寄生植物种群的扩散机制需要进行进一步探索,本研究提出的桑寄生植物的种群动态与大气污染物之间联系的假设则有待于进一步验证。
致谢:感谢张华夏对2012年数据调查的贡献和初始研究工作,感谢张晓娜和刘琼在论文撰写过程中提供的宝贵意见。
猜你喜欢
环球时报(2022-04-07)2022-04-07
智慧健康(2021年5期)2021-03-30
世界最新医学信息文摘(2020年15期)2020-03-30
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
热带病与寄生虫学(2019年4期)2019-02-12
江苏农业科学(2017年7期)2017-05-23
小学阅读指南·低年级版(2016年6期)2016-05-14
工业设计(2015年12期)2015-10-21
