
杂交桉家系在桂北生长及优良性评价
2021-03-17 06:43李光友徐建民李昌荣陈健波郭东强陆海飞
中南林业科技大学学报 2021年2期
李光友,徐建民,李昌荣,陈健波,郭东强,陆海飞
(1.中国林业科学研究院 热带林业研究所 广东 广州 510520;2.广西林业科学研究院 广西 南宁 530002)
林木新品系可通过自然授粉选择获得,也能通过亲缘关系相近种源/家系的人工控制授粉来获得,而人工方法能更快实现基因重组、个体分化和目标子代组合[1]。优良个体源自具优良目的基因的亲本组配而成,增加亲本未表现的变异并实现优良特性的聚合,体现杂种优势,得到更多新品系,达到适应环境变化的需求和基因遗传给后代之目的[2]。桉属Eucalyptusspp.作为热带树种,具有生长快、适应性强的特点,且花粉易收集、花期长,花大易采用控制授粉方法实现人工杂交[3]。上世纪80年代以来,以尾叶桉和巨桉为亲本杂交育种受到许多国家重视并取得显著成效,一批优良无性系经选育后引种和推广,其中有来自巴西的优良巨尾桉杂种E.grandis×E.urophylla、中澳合作项目和热林所桉树研究团队培育的抗逆性强且速生的尾巨、尾细等杂交无性系影响较大,得到广泛推广种植[4-8]。
人工杂种创制为选育适应性更强、目标更明确的品种创造了条件。为评价和选择适应华南抗逆性强的桉树用材品种,采用人工杂交方法,在优良母本上授以抗逆性强桉花粉以获得优良F1 代,通过子代测定及抗性调查选择杂种优势强的家系或单株,能满足新品种的不断产出和推广,也有利于营建无性系种子园或高世代杂交育种。本研究以早期种子园选择并保存的优良尾叶桉等为母本,与抗性强的巨桉、邓恩桉和尾叶桉[9]等花粉进行人工控制授粉产生的杂交种并育苗造林,进行子代生长、抗性等研究,同时对子代进行遗传评估及综合性状评价,获得区域适应性强和表型优异的种质资源,以期不断满足可供利用的新无性系系列材料,为持续利用优良亲本和进一步开展分子和抗性育种[10-11]研究提供实践数据和试验材料。
1 材料与方法
1.1 试验材料
亲本来源于中国林业科学研究院热带林业研究所尾叶桉核心育种群体、四川林科院黑龙滩种质资源保存基地和广东饶平林科所赤桉种源测定林。
通过选择尾叶桉、邓恩桉、赤桉、巨桉及其广泛推广种植的尾巨桉优良无性系作为母本,并收集邓恩桉、尾叶桉、细叶桉、赤桉、巨桉、刚果 12 号桉和尾细杂交桉等优良单株花粉材料进行人工控制授粉。杂种组合及参试材料见表1。2006年8—12月开展控制授粉研究工作,2007—2008年收获控制授粉组合及自由授粉种子,2009年10月播种,2010年春季布置田间试验[8],以DH32-29 无性系为对照。
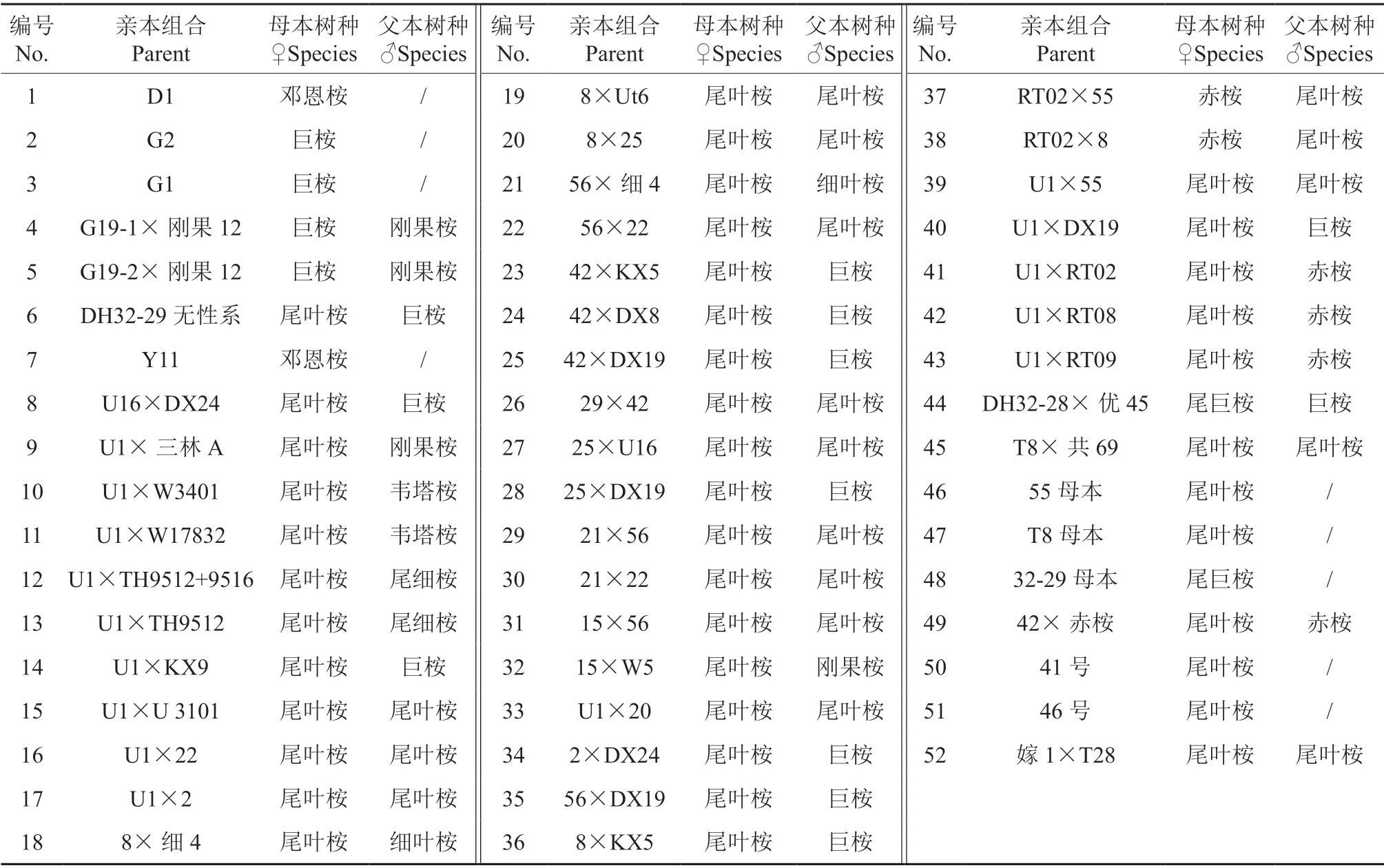
表 1 参试杂种桉组合及亲本概况Table 1 Parents information of the hybirds in trials
1.2 试验地概况与试验设计
试验点广西黄冕林场,位于109°43′46″~109°58′18″E,24°37′25″~24°52′11″N,高丘地貌,海拔高度300~350 m,林地土壤主要是砂页岩发育而成的红壤。林场属中亚热带气候,温暖多雨、水热同季、光照充足、干湿季节明显,造林所在地年平均气温19℃,年平均降水量1 750~2 000 mm,降水集中在4—8月;年均蒸发量1 426~1 650 mm。土壤类型为砂页岩为主形成的红壤,表层土厚度20~25 cm,土层厚度50~70 cm,pH 值5.8,属酸性土类型[12-13]。造林地前茬为杉木Cunninghamia lanceolata,林下植被有三叉苦Evodia lepta、野漆Toxicodendron succedaneum、毛叶算盘子Glochidion hirsutum、铁芒箕Dicranopteris dichotoma、纤毛鸭嘴草Ischaemum indicum和竹类Bambusaspp.等。试验为RCB 设计,单行6 株,6 次重复。
1.3 观测和调查
调查不同年度树高、胸径等生长因子;0.5 a时调查统计各株受枝瘿姬小蜂危害情况,嫩芽呈现一定的危害症状,虫瘿数不少于5 个则计入受害,反之则未受害,最后按家系统计受害率%(或健康率%=100-受害率%);3.5 a 时调查干形指标并打分,干形3 个等级:主干通直圆满,得3 分;主干直、不圆满,得2 分;主干稍弯曲、不圆满,得1 分。具体调查和分析方法参见陈健波等[12]的论文。
1.4 统计分析
1.4.1 数据处理及方差分析
干形、健康率和生长性状采用各年度调查数据进行计算分析。单株蓄积计算按如下公式[13]:
SV=1/30 000×H×DBH2。
用SAS System 广义线性模型完成不平衡数据方差分析,具体参考黄少伟等[15]的方法及模型。
1.4.2 多性状综合坐标得分评定
采用多性状综合坐标评定法[16],通过数据转换后计算得分值。计算公式为:Σpi2=ΣKj(1-aij)2,得分值Σpi2越小则表明该家系越优良。
1.4.3 遗传力和遗传增益
利用SAS 的Proc Varcomp 程序来计算单株和家系遗传力,根据综合坐标法评定出的优良家系确定入选率,按估算公式求遗传增益ΔG=(-)×h2/[11],采用黄少伟等[15]和朱映安等[17]的方法。
2 结果与分析
2.1 不同桉类型的生长变化趋势
一般认为,杂种中母本性状占优势[1],由此以母本进行树种生长性状的统计,将参试材料以母本来源进行归类,共归入6 大类(无性系DH3229、尾巨桉、邓恩桉、巨桉、赤桉及尾叶桉),比较不同类型桉树在3 个林龄(3.5、5.5 和8.5 a)时的生长变化趋势,获得不同桉类型的生长变化规律,结果见图1。
由图1可见,不同桉类平均树高、胸径、单株蓄积随着林龄的增加而增加,在3.5 a 时无性系与家系生长差别最大,8.5 a 时性状间差别最小;各类桉单株蓄积3.5~5.8 a 时以尾巨桉家系增幅最大,对照DH3229 增幅最小,5.8~8.5 a时以邓恩桉家系增幅最大,对照增幅最小,两个年龄段单株蓄积增幅减小的只有尾巨桉家系;3.5~8.5 a 期间林分保存率家系由71.02%降为18.92%,无性系由88.89%降为41.67%,无性系保存率高于家系;不同类桉保存率随着林龄增加而降低,8.5 a 时对照无性系保存率最高,巨桉家系保存率最低,保存率在5.8~8.5 a 期间降幅大,8.5 a 时以对照无性系保存率最高,巨桉家系保存率最低,保存率降幅最大的是尾叶桉家系。
2.2 不同家系/无性系生长性状的方差分析
对杂交组合3 个年度调查的生长数据进行方差分析,结果见表2。
从表2可以看出,42 个杂交和9 个自由授粉家系性状在不同林龄的家系间均达到极显著,区组仅3.5 a 树高、3.5 a 单株蓄积、5.8 a 树高/胸径和材积以及8.5 a 胸径达到显著或极显著差异,交互作用间仅3.5 a 树高、3.5 a 单株蓄积达到极显著差异,其他性状间则没有差异。结果表明杂交子代及自由授粉家系间的生长水平存在较大差异,由此可进行该立地条件下的优良家系选择。
2.3 生长性状多重比较
以8.5 a 性状分析数据作Duncan 显著性检验,部分结果见表3。结果表明,在9 个自由授粉子代中,以47 号尾叶桉、7 号邓恩桉和3 号巨桉家系生长排名前3。对照DH3229 在8.5 a 时树高和胸径分别为22.89 m 和19.25 cm,单株蓄积仅排名第21 位。

图1 不同性状在3 个年份的变化图Fig.1 Changing trend of traits at three different ages

表2 参试家系3 个年度生长性状方差分析†Table 2 Variance analysis for growth traits at different ages

表 3 51 个85年杂交桉家系不同性状的邓肯氏检验†Table 3 Duncan test of different traits among 51 hybrids at 8.5 a
经比较,8.5 a 时单株蓄积高于对照DH3229无性系平均值0.288 0 m3的有17 个杂交和3 个自由授粉家系。最优自由授粉家系47 号(尾叶桉)排在全部参试家系的第7 位,表明有6 个杂交家系具有明显的杂交优势;参试杂交组合中大于母本48 号(DH3229 自由授粉子代)的杂交组合有26 个和3 个自由授粉家系。树高生长前3 名杂交家系是36、13 和28 号,分别为24.87、24.19 和23.92 m;胸径生长前3 名杂交家系是20、39 和36 号,分别为24.80、22.95 和21.01 cm;单株蓄积最大杂交家系前3 名是20,39 和36 号,分别为0.479 8、0.414 5 和0.367 9 m3,20、39 和36 号健康率均达到100%,20 和36 号家系干形得分排名第7、9 名;干形最优前3 杂交家系为23、6 和44 号,仅23 号得分为3.00;无枝瘿姬小蜂危害的家系有42 个,占82.35%。其中3、4 和5 号的危害最严重,母本均为巨桉。
2.4 家系优良性评价
对参试51 个家系及1 个对照无性系测定性状进行坐标综合法评定,按总平均分值±1 个标准差值(0.037 4 和0.144 4)为标准,低于0.037 4值的家系归于适生型(adaptation-A)家系,共7 个家系;高于0.144 4 值的家系归于淘汰型(elimination-E)家系,共6 个家系;其他39 个家系归于中生型(mean-M)家系(含对照无性系)。现将所有家系归属类型列于表4。

表4 参试家系/无性系按得分值划分类型Table 4 Three types of all families according to coordinate assessment
由表4可见,入选的7 个适生家系(含自由授粉)其母本均为尾叶桉,父本包括尾叶桉、巨桉和细叶桉;淘汰型6 个家系母本包括尾叶桉和巨桉,父本包括尾叶桉、刚果桉、韦塔桉和细叶桉,其中两个巨×刚杂交桉枝瘿姬小蜂危害严重,得分值也较低,表明生长综合水平较低。
2.5 参试家系遗传力和遗传增益
按坐标综合法得分标准归纳的各入选类型中,7 个适生家系的入选率为13.73%;38 个中生家系的入选率为74.51%;6 个淘汰家系占总参试家系的11.76%。以DH3229 无性系(对照)平均生长为标准,以平均得分值大于该无性系的家系为适生家系,此时共15 个家系入选,入选率为29.41%。基于以上两种标准下,进行3 个年度的遗传力及遗传增益计算。结果见表5。
由表5可见,不同入选率条件下,相同林龄的单株和家系遗传力相同;入选率为7/51 时(低于坐标综合法得分值标准0.037 4 的7 个适生型家系),其遗传增益高于入选率为15/51(低于DH3229 无性系对照得分值0.053 5 的15 个家系)时的遗传增益。分析表明,性状的家系遗传力在0.216 4~0.852 9 范围,单株遗传力在0.174 5~0.828 5 范围,家系遗传力高于单株遗传力;7/51入选率下,家系遗传增益值在1.69%~21.36%范围,单株遗传增益在1.37%~19.70%;15/51 入选率下,家系遗传增益值在1.13%~14.74%范围,单株遗传增益在0.91%~13.60%。

表5 尾叶桉家系遗传力、单株遗传力以及遗传增益†Table 5 The heritability and genetic gain of the traits at different ages
2.6 家系内优良单株的选择
以生长最优的DH3229 单株得分值(0.085 8)为对照,将林分中所有存活的单株得分值与之比较,得分值小于对照则入选为优良单株,共有48株入选,入选株所属家系及各家系入选株见表6。

表 6 各杂交家系入选优良单株及平均得分Table 6 Traits values and plus tree in families according to coordinate assessment
从表6可见,共有27 个家系的48 株入选为优良单株,其中入选单株最多的3 个家系分别是40 号、14 号和28 号,分别有6 株、4 株和4 株入选。
3 结 论
全部参试材料以母本进行归类,共归入无性系对照、尾巨桉、邓恩桉、巨桉、赤桉及尾叶桉家系6 类;不同桉类生长水平存在差异,3.5~5.8 a以尾巨桉家系生长最快,5.8~8.5 a 以邓恩桉家系增幅最大,对照增幅最小,两个年龄段单株蓄积增幅减小的只有尾巨桉家系。
42 个杂交、9 个自由授粉家系和1 个对照无性系各年度生长在家系间均达到极显著差异,在其他变异来源上的差异存在较大变化,表明本批试验材料中有值得选优的家系存在,能选择到一批具有杂种优势的子代家系。
自由授粉子代中,以47 号尾叶桉、7 号邓恩桉和3 号巨桉家系生长排名前3,说明尾叶桉、邓恩桉和巨桉在该立地的适应性较强,其中最优自由授粉家系47 号(尾叶桉)排在全部参试家系的第7 位,表明有6 个人工杂种家系(20、39、36、28、14 和18 号,包 括2 个 尾× 尾、3 个 尾×巨和1 个尾×细杂交桉)具有较高的杂交优势,是今后持续杂交育种值得关注的材料;对照DH3229 在8.5 a 时单株蓄积仅排名第21 位,表明20 个家系的生长水平较高,利用价值好。
利用坐标综合法,通过生长和抗性指标将参试家系分为3 大类型,7 个适生型家系、39 个中生型家系和6 个淘汰型家系。
以DH3229 最优生长株得分值为标准,从27 个家系中选择到48 株入选,优株入选率为14.95%,其中入选单株最多的3 个家系分别有6 株、4 株和4 株入选,表明该3 个家系具有整体的优良度,且3 个家系均为尾巨杂交桉,表明尾巨桉值得关注和持续利用。
4 讨 论
参试的无性系对照、尾巨桉、邓恩桉、巨桉、赤桉及尾叶桉家系6 类中,以对照无性系具有稳定的增长和较强的适应性,原因可能在于无性系最初的苗木质量较好,造林后更快成活从而有较强抗逆性,而其它类别的表现是基于家系的平均表现,与无性系表现具有一定差距,并不表明家系优株的利用价值不高或不适合优树选择,其今后利用方向以优株无性系化,通过无性系测定与目前对照表现来比较才更合理,适应性才更具参考价值。6 类材料中,有的类保存率较低,但研究和今后应用的材料选择以优良人工或自然杂交家系为主,因此在51 个家系和1 个无性系中,由于某些家系表现不佳影响了其所归类的保存率,反而这是研究需要找到或发现的目标家系,家系表现好就保留,家系表现不好就淘汰。同类材料中存在保存率高的家系和优株,是今后材料持续利用的基础,而合并归类后可能会造成该类的平均保存率或高或低,但并不影响其中优良家系和优株的入选。
研究材料中无枝瘿姬小蜂危害的家系有42 个,占82.35%,受害最严重的家系母本均为尾叶桉,父本为细叶桉、赤桉等[12]。综合其他研究者资料[10,19],具有细叶桉、赤桉等桉树易受枝瘿姬小蜂危害,本研究与之结论相同,不同的是,3 个受害严重的以巨桉为母本的家系,除1 个自由授粉父本未知,另2 个父本为刚果桉,此严重受害家系与张照远等[20]研究的结论相同,但还可能与父本刚果桉易受虫害危害有关。
适生型母本均为尾叶桉,父本包括尾叶桉、巨桉和细叶桉,表明尾叶桉适合在杂交育种中作为母本,以尾叶桉和巨桉为父本,易选择到优良子代;而淘汰型6 个家系的父本除了包括易染虫害的细叶桉外,还包括尾叶桉、刚果桉和韦塔桉。结合虫害在林分幼龄期存在较大危害,严重影响生长和保存,可以直接利用健康率低来淘汰这几个家系,试验中保留其继续生长,通过研究其得分值同样验证了由于抗性差影响了生长综合表现,最终将其归于淘汰型家系是合理的。
在遗传力相同条件下,入选率低则遗传增益较高;性状的遗传力随着林龄增加有降低的趋势,可能与林分立地条件影响下保存率下降有关;相同入选率条件下,性状的家系遗传力高于单株遗传力[21],入选率越高则遗传增益越小。
综合分析自由授粉家系以尾叶桉、邓恩桉和巨桉为母本具有较好表现,杂交桉按母本归类的以尾巨桉、邓恩桉和巨桉具有较好表现,按得分值归类的适生型家系以尾叶桉×尾叶桉和尾叶桉×巨桉具有较好表现,这表明在桂北立地下尾巨桉等具有生长适应性和较高的生长水平,今后为防止病虫害暴发而采用多系种植人工桉用材林,则需要加入表现较好的不同尾叶桉家系间的杂交子代,同时考虑加入邓恩桉、尾巨桉和巨桉为母本的杂交家系或由家系优株选育的无性系。
致谢:协助和参与部分工作的还有项东云、任世奇、邓玉华等,在此一并致谢!
猜你喜欢
世界热带农业信息(2022年11期)2022-10-18
养猪(2022年4期)2022-08-17
环球时报(2022-06-09)2022-06-09
现代园艺(2021年8期)2021-05-13
绿色科技(2017年3期)2017-03-14
现代农业科技(2016年20期)2016-12-20
当代体育·扣篮(2016年15期)2016-09-27
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
山东农业科学(2014年1期)2015-03-09
