
河南地方绵羊品种LHR基因的多态性分析
2021-03-15 13:25邵俊红付俊伟董智豪白俊艳陈梦柯付学言韩志洪崔晗晗
当代畜禽养殖业 2021年1期
邵俊红,付俊伟,董智豪,白俊艳*,陈梦柯,付学言,冀 祥,韩志洪,崔晗晗,彭 彬
(1.河南省汝州市动物卫生监督所,河南 汝州 467500;2.河南科技大学动物科技学院,河南 洛阳 471023)
促黄体素受体 (LHR)是属于G蛋白偶联受体(GPCR)超家族中的糖蛋白亚家族成员,其氨基酸序列和结构有高度同源性,具有7个跨膜区、4个胞质区以及由3个环状区和N端组成的细胞外区,细胞外区中大约有14个不完全重复的富含亮氨酸的序列。其不但有与跨膜区发生反应后介导信号传导的特性,而且还具有结合黄体生成素 (LH)的作用。葛立军等[1](2007)在多种动物包括灵长类动物、猪、牛、火鸡以及人的子宫内都发现有促黄体素(LH)或人绒毛膜促性腺激素(HCG)及其mRNA的结合位点。狄冉等[2](2009)检测了促黄体素受体(LHR)基因外显子11在性早熟、高繁殖力山羊品种(济宁青山羊)和性晚熟、低繁殖力品种(波尔山羊、安哥拉山羊和内蒙古绒山羊)中的单核苷酸多态性,结果发现,仅引物P3和P6的扩增片段具有多态性。申颖等[3](2009)在探究LHR基因在济宁青山羊发情周期的不同阶段于子宫中的表达差异时,发现LHR mRNA在整个发情周期的子宫中都有表达,但各时期表达量不同。王利红等[4](2011)在探究绵羊促黄体素受体基因(LHR)外显子11多态性与繁殖性能的相关性的过程中,发现设计的引物中只有两个引物的扩增片段具有多态性。本研究将PCR-SSCP技术与测序相结合,对小尾寒羊、大尾寒羊和豫西脂尾羊的LHR基因进行种群内和种群间的多态性分析,为绵羊品种的改良和保护提供科学依据。
1 材料与方法
1.1 试验材料
分别采集来自濮阳的小尾寒羊、来自平顶山的大尾寒羊和来自偃师乌龙村的豫西脂尾羊的血样,各48只,采用颈部静脉采血方法,经抗凝处理后在冰壶中低温保存运回实验室,血样于-20℃条件下保存备用。
1.2 试验方法
引物由北京鼎国昌盛生物技术有限责任公司合成。 具体为:F:5'-TCGTTTCCTCATGTGCAATCT-3';R:5'-GGTATGCCATCTTTCTAGTGTGAT-3',扩增产物片段大小为200 bp。PCR反应体系的总体积为15 μL,其中 1 uL 的 DNA,上、下游引物各为 1 μL,去离子水 4.5 μL, 以及 7.5 μL 的 2×Taq PCR Green Mix。PCR 扩增程序为:94 ℃预变性 4 min,35×(94 ℃变性 40 s,58 ℃退火 1 min,72 ℃延伸 80 s),72 ℃继续延伸5 min,最后于4℃条件下保存。PCR产物经1%琼脂糖电泳,在紫外投射仪中进行观察,确定为阳性后在12%非变性聚丙烯酰胺凝胶上进行电泳。利用Excel软件统计基因型频率、基因频率以及多态信息含量等相关指标。
2 结果与分析
2.1 绵羊LHR基因的琼脂糖电泳结果
如图1所示,通过梯度PCR得知,58℃是该引物的最适退火温度。对该退火温度下的PCR产物用1%琼脂糖凝胶进行电泳检测,结果发现特异性扩增良好,无拖尾,与预期片段大小200 bp相符,可进行SSCP分析。

图1 绵羊 LHR基因的琼脂糖检测结果
2.2 绵羊LHR基因的聚丙烯酰胺凝胶电泳结果
对小尾寒羊、大尾寒羊、豫西脂尾羊促黄体素受体(LHR)外显子11的PCR产物进行电泳,结果见图2。可以发现,在小尾寒羊中检测到AB、BB两种基因型,在豫西脂尾羊中检测到AA、AB、BB三种基因型,在大尾寒羊中检测到AB、BB两种基因型。
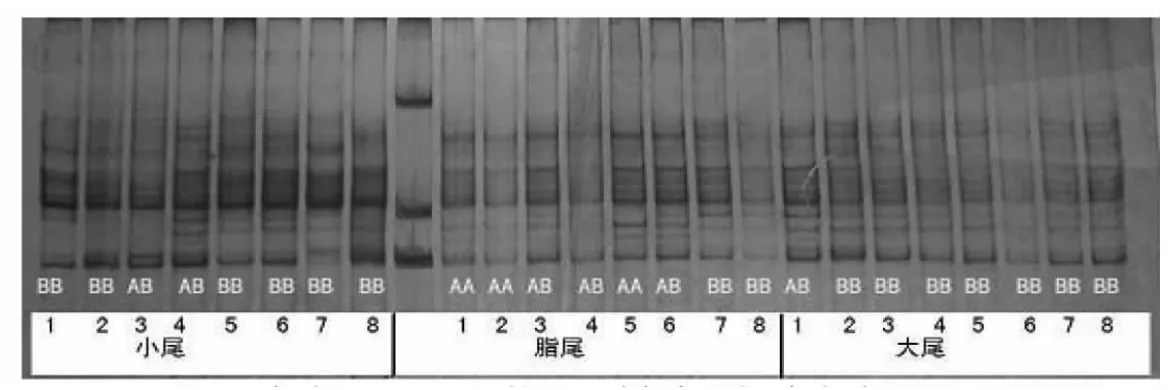
图2 绵羊LHR基因的聚丙烯酰胺凝胶电泳图
2.3 绵羊LHR基因的群体遗传学分析
从表1可以看出,促黄体素受体(LHR)外显子11在小尾寒羊、大尾寒羊和豫西脂尾羊中存在三种基因型,记为AA、BB和AB。其中小尾寒羊AA的基因型频率为0.045,AB的基因型频率为0.295,BB的基因型频率为0.659。豫西脂尾羊AA基因型频率为0.174,AB的基因型频率为0.261,BB的基因型频率为0.565。大尾寒羊AB基因频率为0.098,BB基因频率为0.902。促黄体素受体(LHR)外显子11在小尾寒羊中的遗传杂合度(He)为0.312,有效等位基因数(Ne)为 1.452,多态信息含量(PIC)为 0.263。在豫西脂尾羊中的遗传杂合度 (He)为0.423,有效等位基因数(Ne)为 1.734,多态信息含量(PIC)为 0.256。在大尾寒羊中的遗传杂合度 (He)为0.093,有效等位基因数(Ne)为 1.108,多态信息含量(PIC)为 0.089。
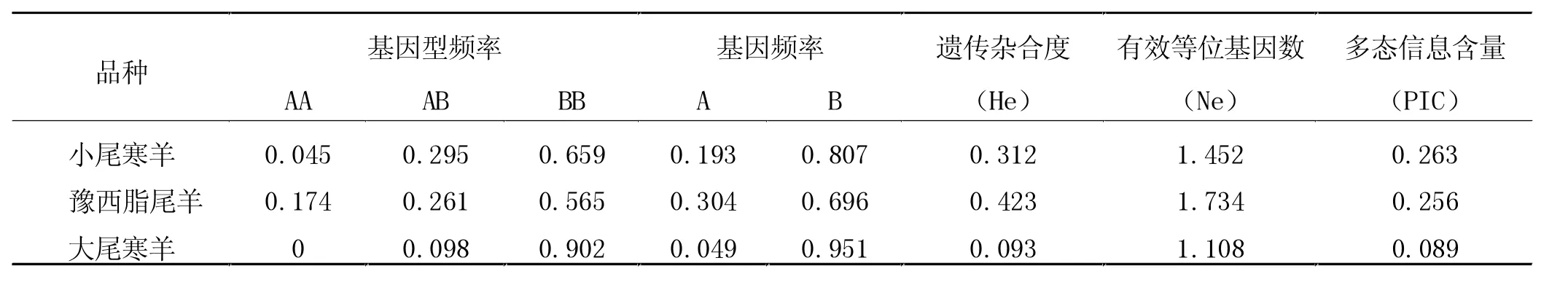
表1 绵羊LHR基因的群体多态性分析
2.4 绵羊LHR外显子11的测序分析
选择LHR外显子11的AA、AB、BB这三种基因型的PCR产物直接测序,测序比对结果见图3。发现不同基因型之间有5个位点发生突变,LHR在37位发生G、A的突变;在49位发生T、C的突变;在93位发生T、C突变;在139位发生T、G突变;在184位发生A、G突变。

图3 绵羊LHR基因的不同基因型测序结果
3 讨论
LHR基因是动物体内较为重要的调节因子之一,国内外许多学者都对其进行了研究。本研究采用PCR-SSCP技术分析了小尾寒羊、豫西脂尾羊和大尾寒羊LHR基因外显子11的多态性,SSCP结果同样检测到了AA、AB、BB这三种基因型。此外,寸静宇[5](2019)在探究绵羊LHR基因的多态性时发现,其所探究的LHR基因7个SNPs在大多数绵羊品种中均表现为中度多态 (0.25<PIC<0.5),且试验中该基因的所有位点在各个绵羊品种中均处于Hardy-Weinberg平衡状态(P>0.05)。 吴翠玲等[6](2019)在研究新吉细毛羊中5个生殖激素受体基因的多态性时发现,LHR基因存在 T1262G、A1991G、C2012T、A2032T 和 T2041A的突变,但这些突变对其产仔数并无显著影响 (P>0.05)。关于其他物种,王爱华等[7](2003)发现,猪 LHR基因外显子11序列发生了一个碱基突变,但并没有造成该段蛋白序列的改变,为沉默突变。以上研究都对LHR基因的SNP位点进行了部分探索,虽与本试验研究目的不一致,但均表明LHR基因具有较高的碱基突变概率。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
中国生殖健康(2020年4期)2021-01-18
意林(儿童绘本)(2019年6期)2019-08-05
小学生必读(低年级版)(2019年12期)2019-04-20
中国生殖健康(2018年4期)2018-11-06
幼儿画刊(2018年7期)2018-07-24
数学大王·低年级(2016年8期)2016-05-14
湖北农业科学(2014年11期)2014-09-10
新疆农垦科技(2014年10期)2014-02-28
当代畜禽养殖业(2014年12期)2014-02-27
