
不同产地木荷优树无性系表型多样性1)
2021-03-06 09:00徐肇友王云鹏
东北林业大学学报 2021年2期
徐肇友 王云鹏
(浙江省龙泉市林业科学研究院,龙泉,323700) (浙江省林木育种技术重点实验室(中国林业科学研究院亚热带林业研究所))
肖纪军 沈斌 王帮顺 周志春
(浙江省龙泉市林业科学研究院) (浙江省林木育种技术重点实验室(中国林业科学研究院亚热带林业研究所))
表型是物种各种形态特征的组合,是植物遗传多样性的最直观反应,也是植物自身基因表达及对环境适应性的综合表现。表型性状作为基因型的外在表征,研究种群的表型多样性可以直观地揭示种群的变异大小。植物经过长期的自然选择,其表型性状对生长环境具有较强的适应性,有研究发现,根据不同种源乔松(Pinuswallichiana)的球果、种子和幼苗的表型变异特点,可将其种群聚为5类,发现地理上遥远的种子源在遗传上却是相近的[1]。不同种群云南松(Pinusyunnanensis)的针叶表型在群体内变异大于群体间,基于针叶性状把所测种群分成了4类[2]。不同种群红松(Pinuskoraiensis)表型性状的种群内变异也大于种群间的变异,具有中等水平的表型分化,将种子园的6个种群归为4类[3]。大量的表型多样性研究表明,相同植物的表型性状在不同地区具有相似性,掌握表型变异规律及对其分类是引种驯化和遗传改良的基础,可为种质资源地收集、保存、挖掘和利用提供理论依据。
木荷(Schimasuperba)为山茶科(Theaceae)木荷属常绿阔叶大乔木,自然分布在北纬31°以南,东经105°以东的广大地区[4],是亚热带常绿阔叶林的主要建群种,也是我国南方各省区重要的生态修复树种、高效的生物防火树种和珍贵的用材树种[5]。前期利用RAPD分子标记研究种源遗传多样性表明,木荷种内遗传多样性丰富,分别有27.4%和72.86%的遗传变异来自种源间和种源内个体间[6]。然而,现阶段木荷的育种工作急需在系统分析表型多样性的基础上,进一步得到有实际意义的育种类群,为良种选育工作提供资源。鉴于此,本文选用来自17个产地的170个木荷优树无性系,分析其生长、形质和叶片性状的变异及相关性,以揭示木荷优树无性系表型性状的多样性,并通过主成分和Q型聚类分析,结合各性状的综合表现形成特定用途的育种类群,以期为优异种质资源地发掘和利用提供理论依据。
1 材料与方法
供试的木荷优树无性系为2013年保存于浙江省龙泉市林科院上圩基地的种质资源库,地理坐标为东经119°5′52″~119°6′27″,北纬28°0′23″~28°1′47″,海拔30~50 m,属中亚热带季风气候,年均气温14.7 ℃,年降水量1 400~2 275 mm,年日照时间1 712~1 825 h,无霜期180~280 d。参试的170个木荷优树无性系来自江西、湖南和福建等3个省份的17个产地,利用1年生容器苗作砧木,于当年3月利用带休眠芽的小枝进行嫁接,接口离地面皆在5 cm以下,试验林采用完全随机设计,定植株行距4 m×4 m。
2018年3月,每个产地选择10个无性系,每个无性系选择生长正常的5个分株进行性状测定。生长性状包括树高、地径和冠幅,形质性状包括树干通直度、枝下高、最大分枝角和最大分枝粗。树高、冠幅和枝下高均用米尺测量,嫁接口地径(即地面以上20 cm处的直径)和最大分枝粗用胸径尺测量,最大分枝角(一级分枝与树干的最大角度)用量角器测量。树干通直度分为通直、较通直、一般、弯曲、严重弯曲5级分别记分为5、4、3、2和1,数字越大越通直。在每个无性系分株靠阳面的冠层中部随机采集10张叶片,叶片使用EPSON LA2400扫描,Win RHIZO软件测定,叶片性状参数包括叶长、叶宽(从叶尖到叶基最宽的位置)、叶面积、叶周长、叶宽长比、叶平均宽(从叶尖到叶基宽度的平均值)。手持GPS记录各采样地的经纬度,气候因子数据来源于世界气候数据库World Clim(http://www.worldclim.org/),1950—2000年世界各地气象站的气候观测数据,利用Diva-Gis软件提取。
方差分析统计模型为Yijk=u+Pi+C(P)i(j)+Eijk,式中,Yijk为第i个产地内第j个无性系第k个分株的观测值,u为群体平均值,Pi为第i个产地的效应,C(P)i(j)为第i个产地内第j个无性系的效应,Eijk为机误。在方差分析时,树干通直度经X1/2数据转换,最大分枝角经反正弦转换,最大分枝粗经1/X转换。采用SAS8.0软件的GLM等过程进行性状单因素方差分析,各方差分量用PROC VARCOMP过程中REML方法计算,CORR过程计算Pearson表型相关,采用SPSS软件进行主成分和系统聚类分析。
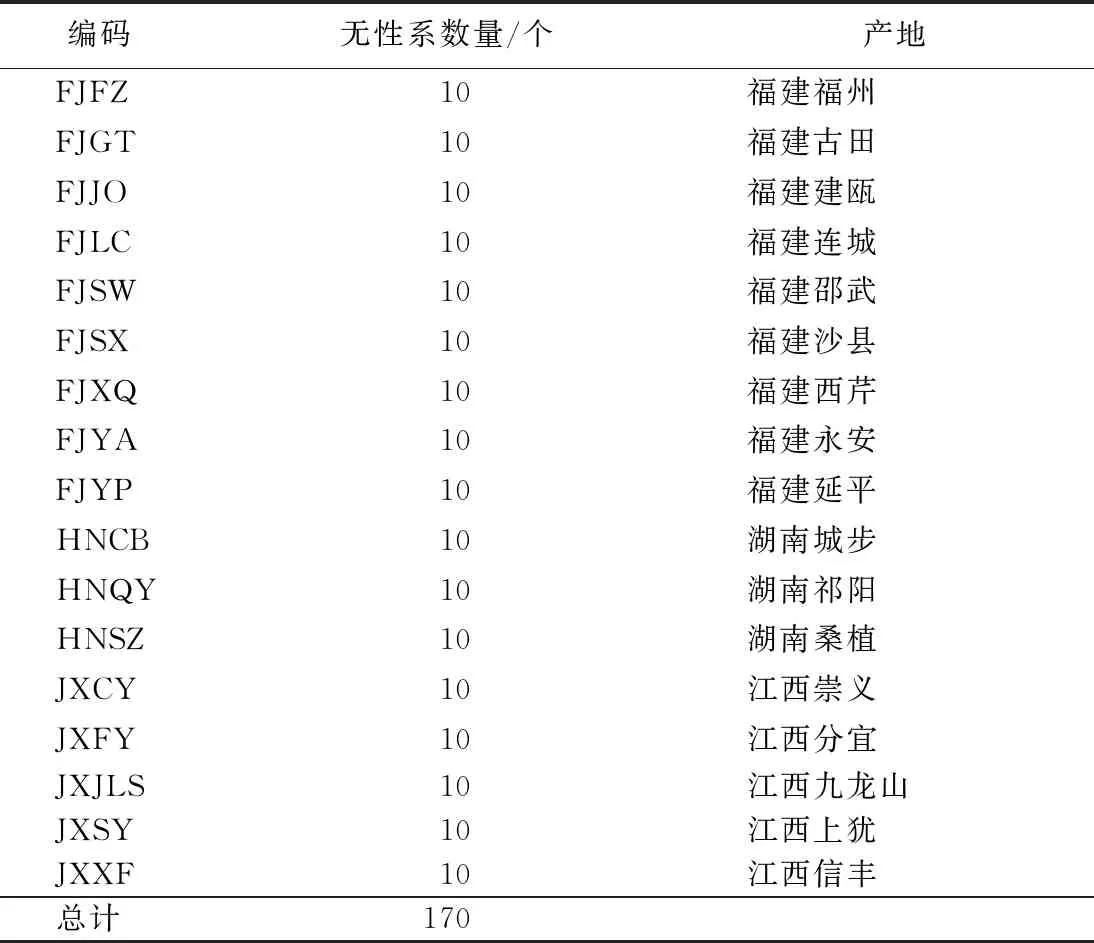
表1 木荷优树无性系来源
2 结果与分析
2.1 木荷优树无性系表型变异
木荷优树无性系表型变异在产地间和产地内均呈极显著的差异(表2)。生长和形质性状的变异系数均很大,变化范围为18.84%~26.60%,其中生长性状中冠幅变异最大(20.38%),树高的变异系数相对较小(18.84%);形质性状中枝下高的变异系数最大(26.6%),变异系数最大无性系高出最小无性系8.37倍,树干通直度次之(25.24%),最大分枝角变异最小(20.77%)。叶片性状的变异系数范围为10.17%~21.25%,其中叶面积变异系数最大(21.25%),变幅为13.34~34.85 cm2,最大无性系高出最小无性系1.61倍,叶平均宽变异系数最小(10.17%),变幅为1.88~2.86 cm。木荷优树无性系在生长、形质和叶片性状存在的显著变异,为以珍贵优质用材和生态修复等培育目标的优良无性系选育提供了很大的选择潜力。
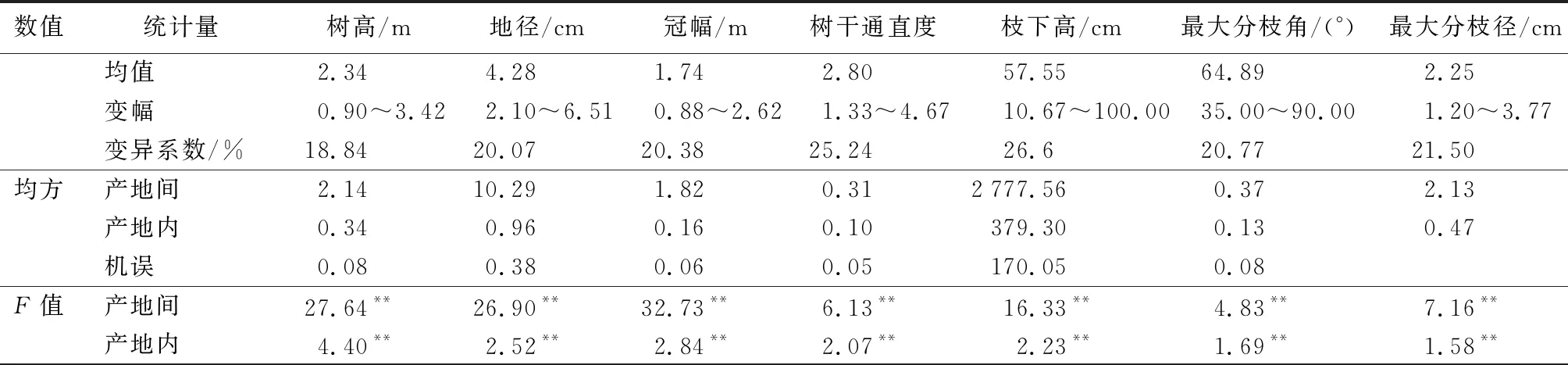
表2 木荷优树无性系表型相关性
2.2 木荷优树无性系表型分化
木荷优树无性系的树高、地径、冠幅和枝下高的产地间变异大于产地内,而其他性状产地间的变异均小于产地内(表3)。13个表型性状产地间和产地内的平均方差分量百分比为25.05%和31.42%,说明木荷优树的表型变异主要来自产地内,产地内多样性大于产地间。产地间和产地内的平均表型分化系数为43.24%和56.76%,说明木荷优树处于中等分化水平。表型分化系数最大的为冠幅(69.79%),最小的为叶平均宽(11.56%),其中生长性状、形质性状和叶片形状的平均表型分化系数分别为63.78%、49.12%和29.04%,说明生长性状的变异是木荷优树表型多样性的主要因素。
2.3 木荷优树无性系表型相关性
优树无性系树高、地径和冠幅3个性状相互之间均呈极显著的正相关(表4),相关系数在0.661~0.764。枝下高与树高、地径、冠幅间,以及树干通直度与树高、枝下高间皆呈极显著的正相关,而树干通直度却与最大分枝角呈极显著的负相关,说明树干的通直度与其速生性有关,速生的优树无性系其树干通直、枝下高大。最大分枝粗与树高、地径、冠幅呈极显著的正相关,而最大分枝角与树高、地径相关性不显著,意味着速生的优树无性系其分枝较粗,而高径生长似与最大分枝角相互独立,因此对速生无性系的选育要加强早期修枝,以培育优质干材。
木荷优树无性系生长性状与叶片性状的相关性分析结果显示(表5),树高、地径、冠幅与叶面积、叶周长、叶长、叶宽、叶平均宽均呈极显著的正相关,说明速生的优树无性系其叶片宽且长、叶面积大。然而树高、地径却与叶宽长比呈显著的负相关,这暗示叶片狭长的无性系生长较慢,而叶片宽圆的无性系可能生长更快。
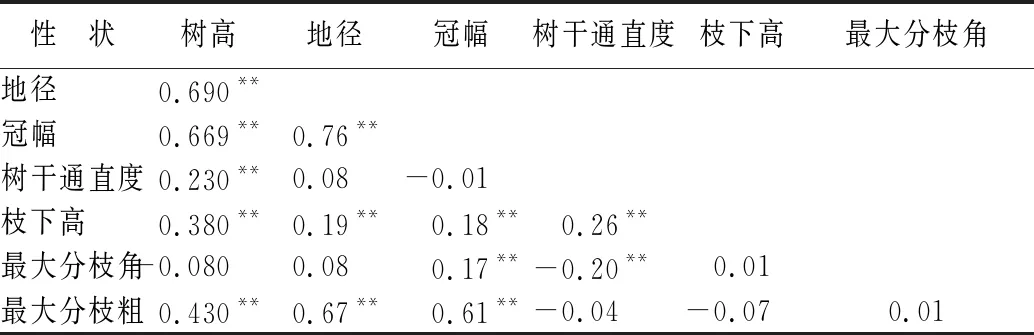
表4 木荷优树无性系生长与形质性状间的相关性
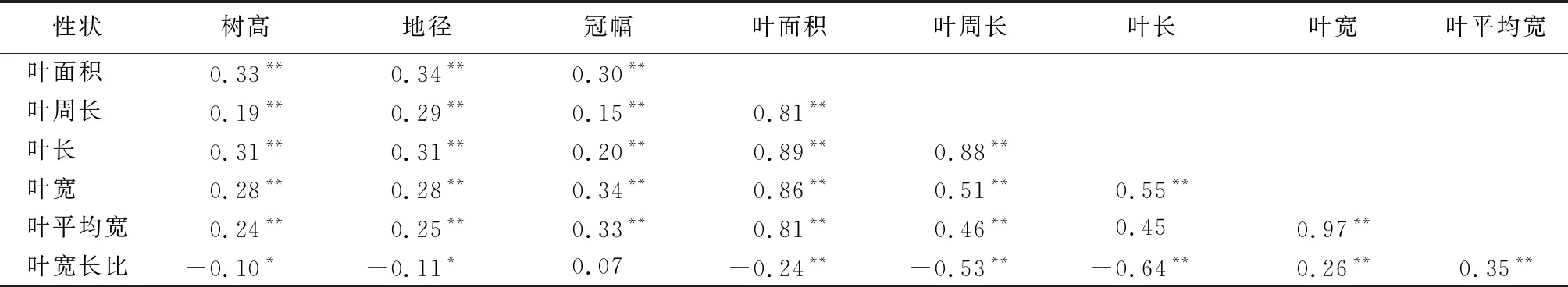
表5 木荷优树无性系生长性状与叶片性状间的相关性
2.4 木荷优树无性系表型与地理气候因子间相关性
优树无性系树高与其产地纬度呈显著负相关(表6),说明来自南部地区的木荷优树生长较快。树干通直度与经度和最冷季降水量呈显著或极显著的负相关,说明来自西部且最冷季降水量少的地区,木荷优树树干通直度较高。最大分枝角与经度、年均气温、最暖季平均气温、最冷季平均气温和最冷季降水量呈显著或极显著正相关,说明在东部气温较高且冷季降水量大的地区,木荷优树的分枝角度较大。叶周长与经度和年均气温、最暖季平均气温、最冷季气温呈显著或极显著负相关,说明在东部且气温较高的地区,木荷优树的叶周长较小;叶长与经度、年均气温、最暖季平均气温和最冷季降水量呈显著或极显著负相关,而叶宽长比相反,说明在东部年均气温较高且冷季降水量大的地区,优树无性系的叶片较短且宽圆。
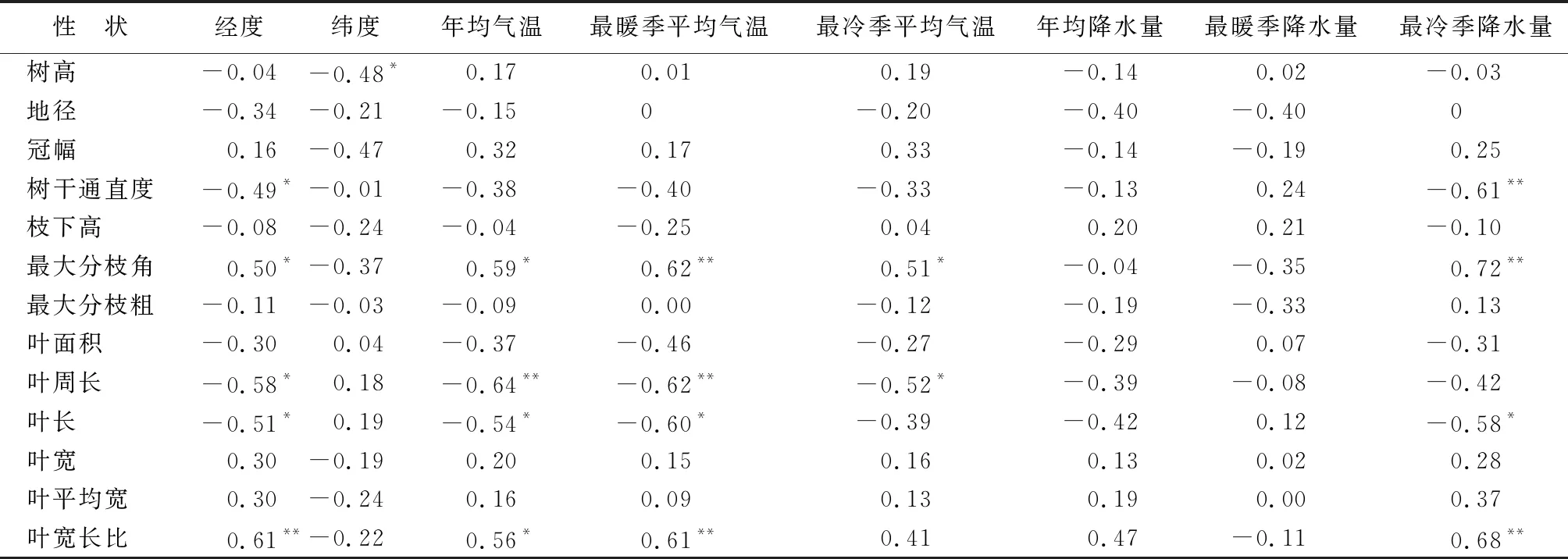
表6 木荷优树无性系表型与地理气候因子间的相关性
2.5 木荷优树无性系表型的主成分分析
对木荷优树无性系13个性状进行主成分分析(表7),前面4个主成分的特征值均大于1,有效包含了13个性状82.11%的信息,表明这4个主成分代表了13个性状的变异信息。由表4可知,主成分1(PC1)主要涵盖了叶面积(0.909)、叶长(0.807)、叶宽(0.791)和叶周长(0.753)等性状,其中叶面积起主导作用;主成分2(PC2)主要涵盖了冠幅(0.650)、地径(0.545)、最大分枝粗(0.486)和树高(0.480)等性状,其中冠幅起主导作用;主成分3(PC3)主要涵盖了叶宽长比(0.727),叶平均宽(0.609)和叶宽(0.551)等性状,其中叶宽长比起主导作用;主成分4(PC4)主要涵盖了枝下高(0.630)和树干通直度(0.523)等性状,其中枝下高起主导作用。表明冠幅、枝下高和叶片性状中的叶面积、叶宽长比是优树无性系的主要变异因子。
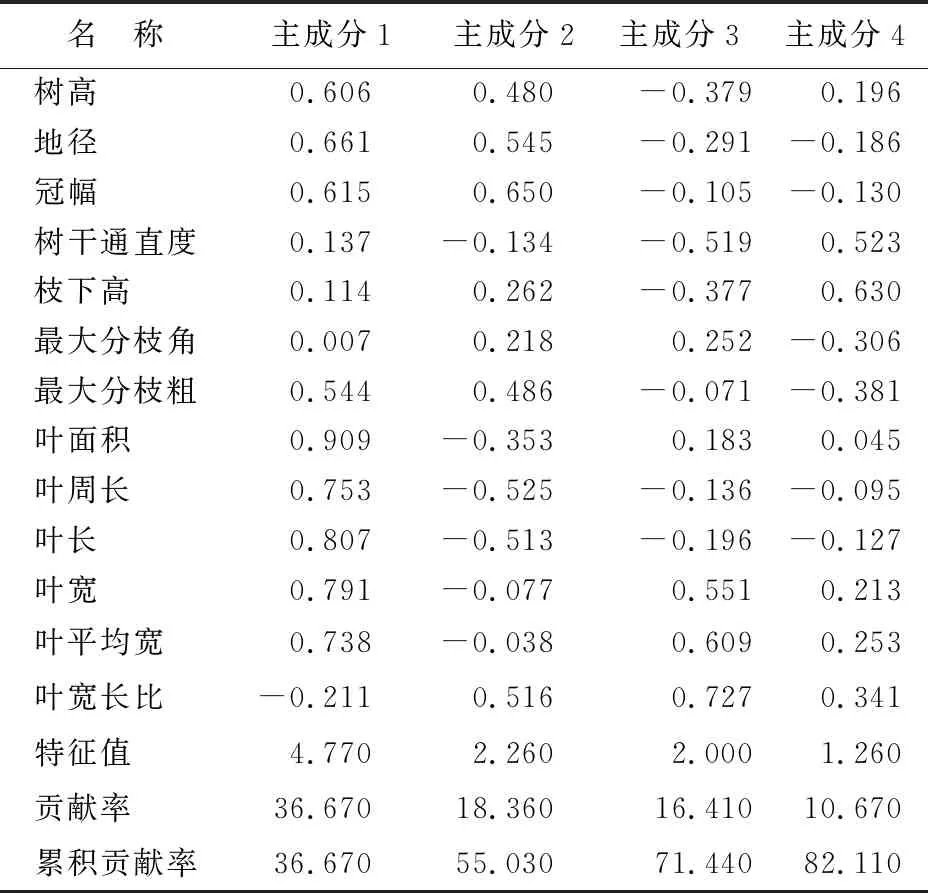
表7 木荷优树无性系表型的主成分分析结果
2.6 木荷优树无性系聚类分析
基于13个生长、形质和叶片形态等表型性状对17个木荷优树无性系产地进行Q型聚类分析,在欧氏距离7.5处,将参试的木荷无性系归为4种类型(图1),各类型的主要特征见表8。类型Ⅰ包括福建连城、沙县,江西九龙山、分宜等4个产地,来自中部种源区,占全部产地的23.53%,主要特征是树高、地径、枝下高、树干通直度和叶长最小,而叶宽、叶平均宽和叶宽长比最大,说明此类型优树无性系生长相对较慢,树干通直度较低,叶片较短且宽圆。类型Ⅱ包括福建建瓯、古田、福州,江西上犹和湖南祁阳等5个产地,主要来自水热资源优越的中心种源区,占全部产地的29.41%,主要特征是树高、地径、冠幅和最大分枝粗最大,说明此类型产地的无性系速生性较强。类型Ⅲ包括福建永安、邵武、西芹、延平和江西崇义、信丰等6个产地,主要来自中心或中部种源区,占全部产地的35.29%,主要特征是枝下高和最大分枝角最大,最大分枝粗和叶面积、叶周长最小,树高仅次于速生性最强的类型Ⅱ,说明此类型产地的无性系较速生,叶片较小。类型Ⅳ包括湖南的城步和桑植等2个产地,来自纬度较高、温度较低的北部种源区,占比11.76%,主要特征是树干通直度和叶面积、叶周长、叶长最大,冠幅、最大分枝角和叶宽、叶平均宽、叶宽长比最小,说明此类型的无性系树干通直度较高,冠幅小,叶片较大且狭长。
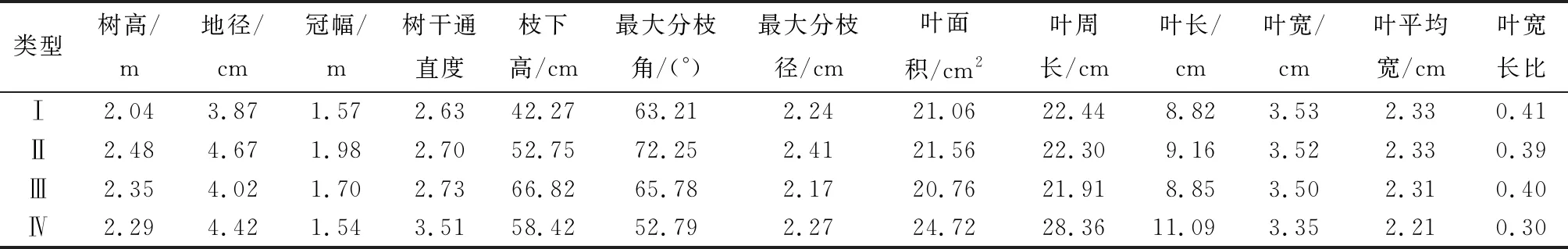
表8 不同类型木荷优树无性系表型性状平均值
3 结论与讨论
因自然选择、基因流和遗传漂变引起的表型变异,在种内十分常见[7]。木荷是中国亚热带常绿阔叶林的主要建群种,广泛分布于南方各省(区、市)[8],生长和分枝性状存在显著的种源效应[9]。本研究表明参试的170个木荷优树无性系的生长、形质和叶片性状在产地间和产地内均存在极显著的遗传变异,冠幅、枝下高和叶面积变异较大,表明这3个性状可能主导木荷优树的变异,树高、最大分枝角和叶平均宽变异较小,表明这3个性状稳定性较高,在一定程度上表明这3个性状受环境的影响较小。从另一面也表明木荷优树无性系在表型性状上具有较大的改良潜力,为选育不同需求的品种提供了重要的种质资源,这也意味着可以根据不同无性系的当代生长表现来初选育种亲本和种子园的建园材料。
表型分化系数是表述表型多样性主变因占扣除环境因素误差后的总方差的比值,其数值可近似地揭示群体间分化的大小,也暗示着该物种适应环境的广泛程度。张翠琴等[10]研究五角枫(Acermono)的表型分化系数为48.82%,种群内变异大于种群间;童跃伟等[3]研究发现种子园内不同种群红松的表型分化系数为12.39%,主要变异来源于种群内;李文英等[11]研究发现蒙古栎(Quercusmongolica)的表型分化系数为53.97%,群体间变异大于群体内变异,李伟等[12]研究发现皂荚(Gleditsiasinensis)的表型分化系数为20.42%,群体内变异大于群体间变异。由此可见,不同物种的群体间表型分化程度是不同的,群体间和群体内不同的分化程度与生态环境、地理隔离、物种个体的发育过程和人为活动有关[10]。本研究发现木荷优树的表型分化系数为43.24%,处于中等分化水平,侧面反应了木荷能较广泛地适应环境的能力。产地内变异大于产地间变异,这与张萍等[6]利用RAPD分子标记和杨汉波等[13]利用SSR标记研究木荷遗传多样的结果具有一致性,可能原因是优树采集地的种群较大,呈集群分布,产地间存在基因交流,导致产地间未发生较大的遗传分化,也有可能与产地间存在相似的形成机制和演化历史有关,但还需进一步验证。
速生性是木荷用材和生态修复的主要选育目标,分析性状的相关性可为良种选育的间接选择提供理论指导。相关性分析结果显示,优树无性系树高与枝下高、树干通直度、最大分枝粗,冠幅与最大分枝角、最大分枝粗皆呈极显著的正相关,这表明枝下高、树干通直度和最大分枝粗均与速生性存在紧密的关联性,而最大分枝角除与冠幅、树干通直度存在极显著的正相关和负相关外,与其它性状的相关性都较低,意味着最大分枝角与速生性似相互独立,这与马尾松(Pinusmassoniana)生长性状和最大分枝角之间相关性的研究结果一致[14]。由于速生无性系的分枝较粗大,为培育速生的优质干材,应加强早期修枝。无性系树高、地径与叶面积、叶周长、叶长、叶宽、叶平均宽皆呈极显著的正相关,而与叶宽长比呈显著负相关,意味着不但叶片大小,而且叶形也与速生性存在紧密的关联性,叶片大和宽圆的无性系生长较快,木荷家系苗期研究[15]发现其苗高与叶片长也存在极显著的正相关关系,这一相关性结果也意味着在木荷良种选育过程中可通过叶片性状来进行间接选择速生性基因型。
植物长期对生长环境的适应和响应,致使其表型性状与环境因子间具有密切的关系,研究它们之间的相关性可以更准确地揭示植物变异格局。本研究发现木荷优树无性系速生性主要受其产地纬度的影响,南部地区的木荷优树速生性较强;形质性状主要受其产地经度和降水量的影响,西部降水量少的地区,木荷优树无性系树干通直度较高,最大分枝角较小。这与3年生不同种源木荷生长性状和分枝性状在福建建瓯试验点的地理变异模式研究结果一致[9]。植物的叶片大小与温度、水分和风速之间存在显著的相互关系,有研究表明,尾叶樱桃(Cerasusdielsiana)的叶片性状自西向东随经度的增加,叶片变小[16]。桂花(Osmanthusfragrans)的叶长和叶宽与温度季节性变化存在显著正相关,叶长宽比与降水量呈显著负相关[17]。灰木莲(Manglietiaconifera)叶周长和叶长宽比与经度呈显著正相关[18]。本研究发现叶片性状受经度、温度和降水量的综合影响,东部温度和降水量较高的地区,木荷优树的叶片较小、短且宽圆,这可能与适应亚热带东部地区高温高湿和强风降水的气候特征有关。
主成分是通过几个变量来揭示多个变量内部结构的统计方法。对170个木荷优树无性系主成分分析结果表明,前4个主成分累计贡献率为82.11%,可以综合反映13个性状的信息。综合主成分分析,从13个性状中筛选出枝下高、冠幅是生长性状中的主要区别性状,叶面积、叶宽长比是叶片性状的主要区别性状。所得结论与桉树(Eucalyptus)中叶长、叶长宽比是主要的区别特征[19],灰木莲中叶长、叶周长和叶面积是主要区别特征的结果[18]有一定的一致性,表明基于表型多样性对木荷优树无性系进行主成分分析的简化分类是有效的。
聚类分析是系统描述和鉴定种质资源的基本方法,牟洪香等[20]根据文冠果(Xanthocerassorbifolia)15个表型性状的差异,将14个分布区分为4类;张翠琴等[10]基于五角枫的23个表型性状将19个种群分为2支。本研究基于13个表型性状,通过聚类分析将参试的17个木荷优树无性系产地归为4个类型,在每个类型内木荷优树无性系的表型性状相近,而在不同类型中则有明显的差异。根据王秀花等[21]基于7年生生长和木材基本密度进行种源聚类,大致将木荷划分为中心种源区、中部种源区和北部种源区。本研究类型Ⅰ产地无性系主要来自中部种源区,其生长相对缓慢,树干通直度较低,叶片较短且宽圆;类型Ⅱ和类型Ⅲ中的无性系主要来自中心种源区和中部种源区,其速生性较强,叶片较小;类型Ⅳ的无性系主要来自北部种源区,树干通直度较高,冠幅小,叶片较大且狭长。根据不同类型的特征,将不同的无性系进行杂交,有望筛选出具有优良特性的子代。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29
林业科学(2022年5期)2022-08-20
绿色科技(2022年11期)2022-07-02
散文诗世界(2022年5期)2022-05-23
绿色天府(2022年2期)2022-03-16
绿色科技(2021年21期)2021-11-26
食品安全导刊(2021年21期)2021-08-30
绿色科技(2016年17期)2016-12-28
