
miR-638靶向下调MACC1抑制胃癌细胞的恶性进展
2021-03-03 09:16吴现磊秦二云张令巧
承德医学院学报 2021年6期
吴现磊,秦二云,孟 妍,张令巧
(濮阳市油田总医院消化内科,河南濮阳 457001)
胃癌(gastric carcinoma,GC)是全世界第6大常见癌症和第2大癌症致死原因[1]。GC的早期诊出率较低,大多数GC患者五年生存率不足20%[2,3]。尽管GC的治疗手段在不断发展,但现阶段临床上GC的一线治疗依然以手术和化疗为主。靶向治疗可能是缓解GC疾病负担的重要途径[4]。因此,探究调控GC细胞发生发展的分子学机制,进而寻找GC中存在潜在治疗作用的靶点,具有重要的临床意义。
MicroRNAs(miRNAs)通过特异性结合靶标的3'端非翻译区(3'-untranslated region,3'-UTR)端来抑制基因表达,介导靶标mRNA降解或抑制靶标mRNA翻译[5]。近年来,miRNAs调控肿瘤发生发展的相关研究越来越多,也包括GC[6,7]。例如,miR-194和miR-93-5p在GC中属于致癌基因,miR-194通过激活wnt信号通路促进了GC的发生发展过程[8],miR-93-5p则是通过激活Hippo途径增强GC细胞的增殖、迁移和侵袭[9],两者在GC中均具备治疗潜力。此外,miRNA可作为早期筛查GC耐药患者的生物标志物[10],例如miR-604低表达大概率指示GC耐药性的存在[11]。上述研究均证实miRNAs和GC发生发展密切相关。
结肠癌转移相关基因1(MACC1),最初被发现是作为原发性和转移性结肠癌的癌基因,其位于7号染色体位置的7p21.1,可通过HGF/MET信号传导通路促进肿瘤增殖、浸润和转移[12-14]。此外,越来越多的研究证明MACC1和其他肿瘤的发生进展也密切相关,如神经胶质瘤[15],肺腺癌[16],声门癌[17]和肝细胞癌[18]等。GC组织中MACC1水平较正常组织明显升高,且其表达水平可作为GC预后的独立指标[19,20]。目前关于MACC1在GC中的研究依然比较缺乏。
本研究探究了miR-638在GC细胞中的生物学功能。我们发现,miR-638在GC组织和细胞中的表达显著降低。此外,过表达miR-638可显著抑制GC细胞的增殖、迁移、侵袭能力。更重要的是,我们证明了MACC1是miR-638的直接调控靶点,miR-638靶向下调MACC1抑制GC的发生发展,可能是治疗GC的一个潜在的治疗靶点。
1 材料与方法
1.1 细胞培养
人胃粘膜上皮细胞GES1(货号:BNCC337969),人胃癌细胞MKN-45(货号:BNCC337682)和MKN28(货号:BNCC341748)均购自北纳生物细胞库。GES1和MKN28培养于含10%胎牛血清的高糖DMEM(含4mM 左旋谷氨酰胺、丙酮酸钠)培养基,MKN-45培养于含10%胎牛血清的RPMI-1640培养基。均置于37 ℃、5% CO2的恒温培养箱进行培养。
1.2 细胞转染
取对数生长期的GC细胞接种于6孔板,数量大约控制在转染前一天细胞密度达到50%最佳。按照LipoFiter 3000试剂盒操作说明进行转染,以50nM的最终浓度稀释于500μL无血清培养基中配置转染液。取10μL Lipofectamine 3000稀释于500μL无血清培养基中,室温孵育5 min后分别与各组转染液混匀,共分为4组。miR-mimics组:转染miR-638 mimics;NC-mimics组:转染NC mimics;NC+oe-MACC1组:转染oe-MACC1质粒;miR-mimics+oe-MACC1组:共转染miR-638 mimics和oe-MACC1质粒。孵育20 min后加无血清培养基培养6h,弃去原培养基,换成10%血清的培养基进行转染。转染结束后将GC细胞细胞置于37 ℃、5% CO2的恒温培养箱进行常规培养。转染24 h后可用于后续实验研究。
1.3 qRT-PCR
胰酶消化各实验组细胞后,使用Trizol溶液裂解细胞,按照RNA试剂盒操作说明提取总RNA,按照TaqMan的逆转录试剂盒说明书操作说明,将RNA逆转录成cDNA。按照qSYBR-Green-containing PCR kit(Qiagen,USA)操作说明进行操作,在BioRad IQTM5Multicolor 实时荧光定量系统对各实验组细胞进行qRT-PCR。分别以U6和GAPDH作为miR-638和MACC1的内参,2-ΔΔCt用以计算相对表达量。每组重复3次。引物序列见附表。

附表 引物序列
1.4 Western blot
胰酶消化处于对数生长期的各实验组细胞,裂解液裂解细胞后提取总蛋白。使用二喹啉甲酸(BCA)法测定总蛋白。取10 μg样品99 ℃下加热5 min高温变性,进行10%的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离蛋白,转至聚偏二氟乙烯(PVDF)膜。室温下用5%脱脂奶粉封闭30 min后,加入稀释的一抗MACC1(Abcam,Cambridge,UK)和GAPDH(Abcam,Cambridge,UK),4 ℃下孵育过夜。Tris盐酸缓冲液(TBST)洗膜3次,加入辣根过氧化酶标记的二抗山羊抗兔IgG(Abcam,Cambridge,UK),室温黑暗条件下孵育1h,TBST洗膜3次,每次10min,加入ECL发光剂进行发光反应并保存条带图片,使用ImageJ软件对条带灰度进行定量。每组重复3次。
1.5 CCK8
收集处于对数生长期的各实验组细胞,进行胰酶消化,加入完全培养基重悬并调整细胞密度为2.5×103个/mL,96孔板中每孔接种5000个细胞,边缘孔填充无菌PBS。根据CCK8试剂盒操作说明进行检测,每孔加入10 μL CCK-8溶液后,于37 ℃,5% CO2恒温培养箱培养3h后,通过酶标仪测量其450 nm处的吸光值。每组重复3次。
1.6 划痕愈合
所有器械均进行提前灭菌,直尺和marker笔需要在超净台的紫外台中照射 30 min进行灭菌处理,预先在6孔板背面用marker笔和直尺均匀画横线进行标记,每孔至少标记5条直线。各实验组细胞经胰蛋白酶消化后接种于6孔板中,每孔加入5×105个细胞,每组设置3个复孔。次日使用100 μL无菌移液管枪头垂直于背后横线并经过孔中心进行划痕画一条线,PBS清洗细胞3次,加入无血清培养基置于37℃、5% CO2的恒温培养箱继续培养,在培养0h和24h时于取样拍照记录,使用Image J软件计算24h时各实验组的细胞迁移率,用以评估GC细胞迁移能力的差异。
1.7 Transwell
铺加基质胶稀释液到transwell小室中,于37℃,5%CO2恒温培养箱培养4 h后成膜。胰蛋白酶常规消化各实验组细胞,吸弃培养液后加入含BSA的无血清培养基进行重悬,上室加入200 μL细胞密度为1×105个/mL的GC细胞悬液,下室加入500 μL含血清的完全培养基,于37℃,5% CO2恒温培养箱培养24 h。取24孔板,每孔加入600 μL 4%多聚甲醛,取出小室并清除残留培养液,轻轻放入24孔板固定15 min,用湿润的棉签轻轻擦去上室膜内侧细胞,将小室反转晾干。取空白的24孔板,每孔加入600 μL 0.1%结晶紫甲醇染液,放入小室染色30 min后取出,PBS清洗残留染液,放在倒置荧光显微镜下观察拍照并计数。每组重复3次。
1.8 流式细胞术周期分析
收集处于对数生长期的各实验组细胞,于无血清的培养基培养24 h后进行胰酶消化,可收获同步化的G0期细胞。将各实验组细胞接种于96孔板中,每孔大约5000个细胞,经预冷的PBS清洗两次后,加入3.5 mL预冷的75%冰乙醇轻柔混匀固定,在4℃冰箱过夜。1500 rpm离心5 min后弃上清,经预冷的PBS再次洗涤清除乙醇,加入0.2 mg糖酸酶 A(RNase A)和1 mL PI/Triton X-100染色液(20 μg PI/0.1% Triton X-100),37℃避光水浴30 min,经流式细胞仪(Beckman)检测细胞周期,通过ModFit LT 4.1软件进行分析。每组重复3次。
1.9 双荧光素酶实验
将含有miR-638结合位点的MACC1的3'-UTR克隆到pGL3荧光素酶报告载体(Promega,Madison,WI,USA)中。使用Quick-change site-directed mutagenesis kit(Agilent Technologies,Santa Clara,CA)对结合位点进行突变。然后参考细胞转染步骤将构建的载体分别与miR-638 mimic或NC-mimic共转染到GC细胞。在37℃,5% CO2的恒温培养箱中培养48 h后,使用裂解液裂解细胞。按照荧光素酶检测试剂盒操作说明书,经单光子检测仪(Biorad,USA)检测荧光素酶活性。每组验重复3次。
1.10 统计学分析
本研究使用GraphPad Prism 8.0进行制图,Image J、ModFit和SPSS 26.0进行实验数据的统计分析,所有数据均以mean±sd的形式表示,两组数据比较采用t检验,多组数据比较采用单因素方差分析,以P<0.05表示差异具有统计学意义。
2 结果
2.1 miR-638在GC细胞中显著低表达
TCGA的GC数据集显示miR-638在GC组织中显著低表达(图1A),miR-638的低表达与GC患者预后不良显著相关(图1B)。随后,qRT-PCR结果同样显示miR-638在GC细胞中的表达水平显著下调(图1C)。综上,miR-638在GC细胞中显著低表达。

图1 miR-638在GC细胞中低表达
2.2 过表达miR-638抑制GC细胞的增殖、迁移、侵袭和生长
为了探究miR-638在GC中的调控作用,我们对各处理组GC细胞进行CCK8、划痕愈合、transwell和流式细胞实验。首先,通过qRT-PCR实验验证单独转染miR-mimics使得GC细胞中的miR-638表达量显著升高(图2A)。相比于NC mimics组GC细胞,CCK8实验发现miR-mimics组的GC细胞活力显著降低(图2B、2C);划痕愈合和Transwell表明miR-mimics组的GC细胞的迁移率和侵袭细胞数目显著降低(图2D、2E);细胞周期结果显示,过表达miR-638显著上调了GC细胞的G1期比例(图2F),表明GC细胞生长受到抑制。综上,过表达miR-638抑制GC细胞的增殖、迁移、侵袭和生长。
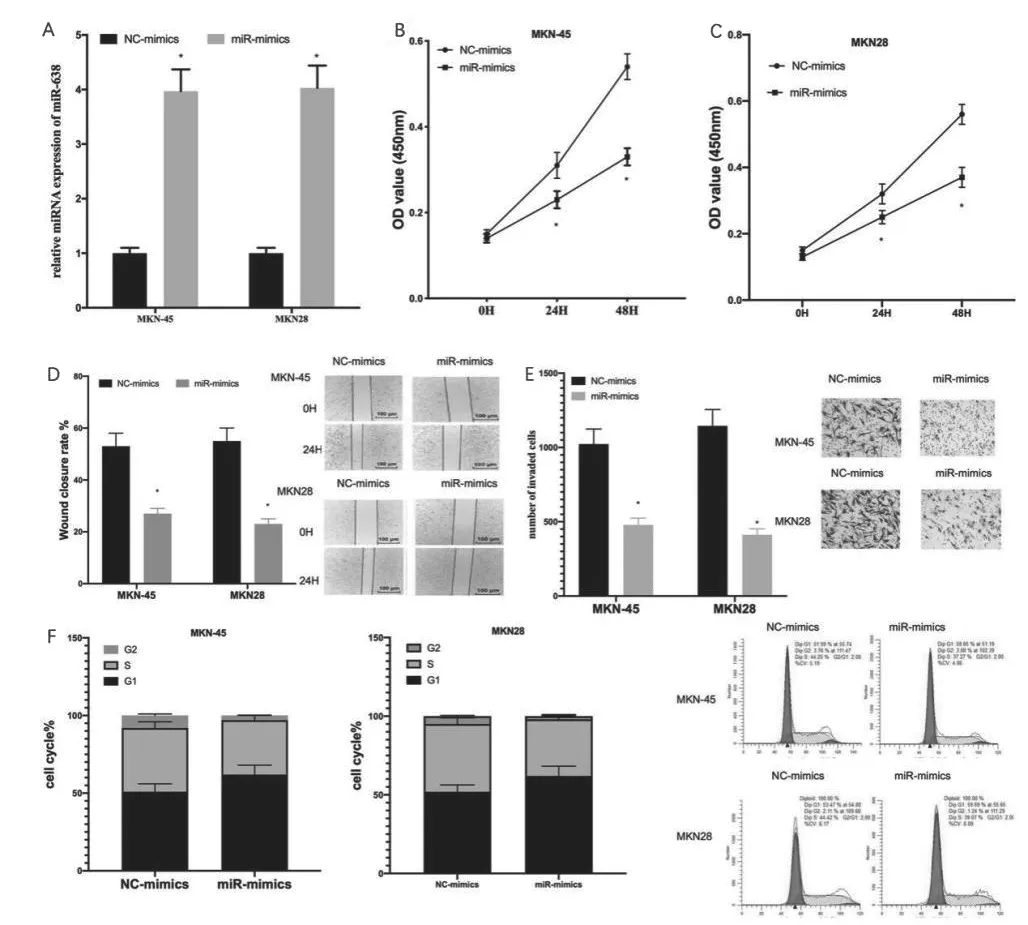
图2 miR-638抑制GC细胞的增殖、迁移、侵袭和生长
2.3 miR-638直接靶向MACC1
TargetScan数据库显示,miR-638和MACC1存在结合位点(图3A)。双荧光素酶报告基因实验结果也表明,过表达miR-638显著抑制MACC1野生型的荧光素酶活性,而对MACC1突变型未产生显著影响(图3B、3C)。qRTPCR表明GC细胞中MACC1显著高表达(图3D)。qRT-PCR和western blot实验发现,过表达miR-638抑制GC细胞中的MACC1的表达水平(图3E)。总结上述实验,证实GC细胞中,MACC1是miR-638的直接靶点,miR-638负调控MACC1。
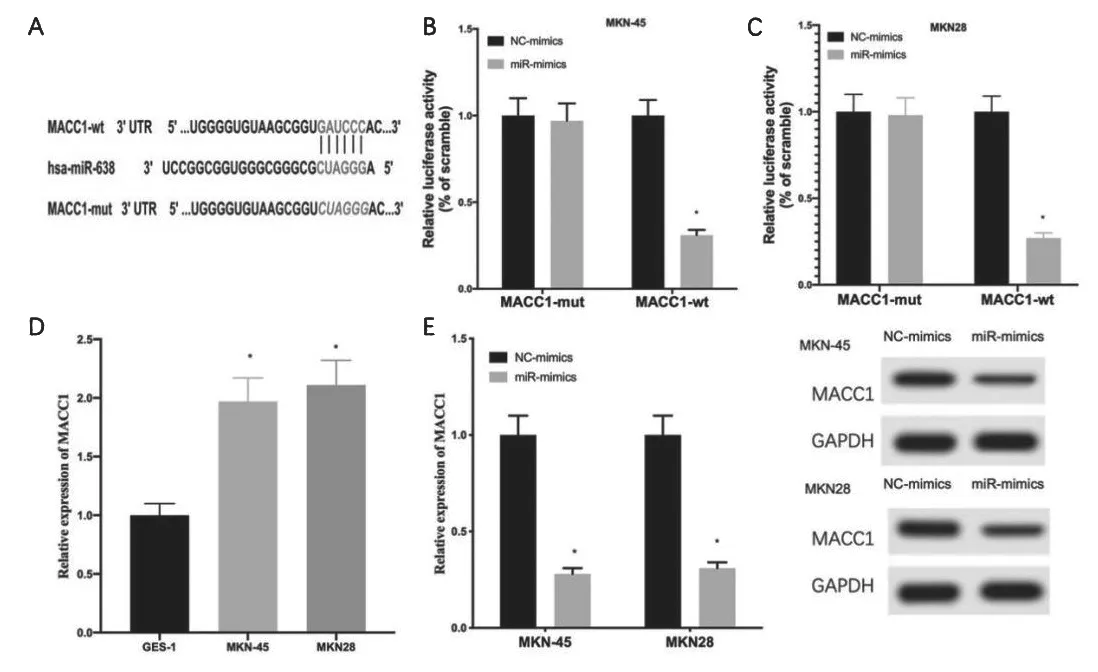
图3 miR-638靶向抑制MACC1
2.4 miR-638靶向下调MACC1抑制GC细胞的增殖、迁移、侵袭和生长
qRT-PCR和western blot结果表明,仅转染oe-MACC1时,GC细胞中MACC1的表达水平显著上调,而同时过表达miR-638后,GC细胞中MACC1的表达水平和NC-mimics组无明显差异(图4A)。比较3组处理组GC细胞的细胞功能学实验结果,发现相比于NC-mimics组,单独过表达MACC1后,GC细胞的OD值、迁移率和侵袭细胞数目均显著升高,GC细胞的G1期比例显著降低,同时过表达miR-638和MACC1时,GC细胞的OD值、迁移率、侵袭细胞数目和G1期比例均与NC-mimics组无明显差异(图4B~4F)。综上所述,MACC1能够促进GC细胞的增殖、迁移、侵袭和生长,而miR-638能够逆转MACC1的促癌作用。

图4 miR-638靶向下调MACC1抑制GC细胞的增殖、迁移、侵袭和生长
3 讨论
GC作为最常见的恶性肿瘤之一,目前仍缺乏预后较为理想的治疗方案。随着靶向治疗相关研究的不断丰富,近期有关miRNAs在GC中靶向治疗的报道也逐渐增多,对GC治疗有所裨益[21],因此研究miRNAs用于GC的靶向治疗备受临床关注。
之前的报道证实miR-638在GC组织和细胞中的表达显著降低[22],本研究也表明miR-638在GC细胞显著低表达。本研究通过一系列细胞功能实验探究miR-638在GC中的调控作用,发现过表达miR-638抑制了MKN-45和MKN28细胞的增殖、迁移、侵袭和生长,这与先前Shen等人的研究结论达成一致[22]。
MACC1和miR-638存在结合位点,此外,MACC1是促进GC生长和转移的致癌基因[23],也能够调节GC细胞中的PDL1表达和肿瘤免疫力,表明MACC1可能是GC的治疗靶点[24]。本研究证实,GC细胞中MACC1的显著高表达后,通过双荧光素酶实验验证了MACC1是GC细胞中miR-638的靶基因。随后的一系列细胞功能实验表明,miR-638能够逆转MACC1对GC细胞增殖、迁移、侵袭和生长的促进作用,即miR-638通过靶向下调MACC1的表达抑制GC细胞增殖、迁移、侵袭和生长,证实miR-638是通过与GC细胞中的MACC1特异性结合,进而发挥其调控作用。
总之,本研究证实了GC细胞中的miR-638表达下调,MACC1表达上调,经实验验证了miR-638靶向MACC1发挥对GC细胞发生发展的抑制作用。综上,本研究认为MACC1和miR-638对GC细胞的调控机制,可为GC的靶向治疗提供另一条有希望的途径。
猜你喜欢
核科学与工程(2022年3期)2022-10-18
现代仪器与医疗(2021年6期)2022-01-18
河北农机(2020年10期)2020-12-14
广州化工(2020年6期)2020-04-18
河北省科学院学报(2020年4期)2020-03-19
食品工程(2020年3期)2020-01-05
现代矿业(2018年9期)2018-10-16
大陆桥视野·下(2017年11期)2017-10-31
计算机测量与控制(2017年6期)2017-07-01
电子制作(2017年24期)2017-02-02
