
miR-32-5p靶向NFIL3基因促进脂多糖诱导的肺上皮细胞凋亡和炎性反应
2021-02-27 05:18李金秀夏天康寿磊
河北医药 2021年1期
李金秀 夏天 康寿磊
肺泡上皮细胞是肺脏结构和功能的基础,肺泡上皮细胞凋亡是急性肺损伤(acute lung injury,ALI)的一个重要病理变化特征,ALI时肺组织严重破坏,呼吸功能严重受损[1]。炎性反应在ALI的发生发展中有重要地位[2]。从分子生物学水平出发,在细胞层面通过探讨特异性靶点可作为干预治疗ALI的潜在手段,而研究发现部分miRNA在肺损伤和修复过程中异常表达,并通过影响其靶基因的表达而调控炎性反应,与ALI的进展密切相关[3]。miR-32-5p参与结核分枝杆菌中的炎性反应,其过表达减弱了炎性细胞因子肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)和白介素-6(interleukin-6,IL-6)的积累[4]。核因子白介素-3(nuclear factor interleukin-3,NFIL3)是一种关键的转录抑制因子,对于NK细胞和先天淋巴细胞的发育至关重要[5,6]。研究发现NFIL3可通过靶向胰岛素样生长因子2受体抑制缺氧诱导的凋亡细胞死亡[7]。然而miR-32-5p和NFIL3对脂多糖诱导的肺上皮细胞凋亡和炎性反应的影响及机制是否与miR-32-5p调控NFIL3相关还尚未可知,本文研究miR-32-5p是否通过调控NFIL3影响脂多糖诱导的肺上皮细胞凋亡和炎性反应,报告如下。
1 材料与方法
1.1 材料 肺上皮细胞A549购自中国科学院上海细胞库;胎牛血清、DMEM培养基、胰蛋白酶、二甲基亚砜(DMSO)、BCA试剂盒、PBS缓冲液购自美国Sigma公司;Lipofectamine TM 2000转染试剂盒购自美国Invitrogen公司;Trizol试剂、反转录试剂盒、荧光定量试剂盒购自日本TaKaRa公司;膜联蛋白V-异硫氰酸荧光素(Annexin V-FITC)和碘化丙锭(PI)试剂盒、双荧光素酶报告基因检测试剂盒购自北京Solarbio公司;酶联免疫吸附试验(ELISA)检测试剂盒购自上海联科生物技术有限公司;RIPA蛋白裂解液、PVDF膜、SDS-PAGE试剂盒购自上海碧云天生物技术有限公司;抗体均购自上海煊翎生物科技有限公司。
1.2 方法
1.2.1 细胞培养与分组:将肺上皮细胞A549在37℃、含10%胎牛血清的DMEM完全培养液中培养,每天换液1次,将融和至80%左右的细胞无血清饥饿培养12 h后进行分组,用终浓度为30 μg/ml的LPS溶液刺激细胞作为LPS组,以未经任何处理的细胞作为正常对照(NC)组。将miR-con、miR-32-5p、anti-miR-con、anti-miR-32-5p分别转染至未经任何处理的A549细胞中,记为miR-con组、miR-32-5p组、anti-miR-con组、anti-miR-32-5p组。将anti-miR-con、anti-miR-32-5p、pcDNA-con、pcDNA-NFIL3转染至A549细胞中再用30 μg/ml的LPS处理,记为LPS+anti-miR-con组、LPS+anti-miR-32-5p组、LPS+pcDNA-con组、LPS+pcDNA-NFIL3组。将anti-miR-32-5p与si-con、si-NFIL3分别共同转染至A549细胞中再用30 μg/ml的LPS处理,记为LPS+anti-miR-32-5p+si-con组、LPS+anti-miR-32-5p+si-NFIL3组;转染均按照Lipofectamine TM 2000转染试剂盒进行操作。
1.2.2 qRT-PCR检测miR-32-5p和NFIL3 mRNA的表达水平:收集各组细胞,研磨充分后加入Trizol试剂提取总RNA,微量核酸测定仪检测RNA纯度和浓度。使用TaKaRa反转录试剂盒将RNA反转录成cDNA,按照TaKaRa荧光定量试剂盒使用说明配制反应体系,以β-actin为内参进行PCR扩增,每个样品重复3次,循环条件为95℃ 30 s,60℃ 30 s;72℃ 30 s,共40个循环;72℃延长5 min。相对表达量采用2-ΔΔCt法计算。
1.2.3 Western blot检测NFIL3、cleave-caspase-3蛋白表达:收集各组细胞,加入RIPA裂解液裂解,4℃,12 000 g离心15 min,收集蛋白上清液,BCA试剂盒测定蛋白浓度。将蛋白样品进行SDS-PAGE电泳后转至PVDF膜上,5%脱脂奶粉封闭液室温封闭1 h。分别加入一抗(1∶1 000),4℃孵育过夜,TBST洗膜;加入二抗(1∶2 000)室温孵育2 h,TBST洗涤3次,每次10 min,后在暗室中曝光显影,再浸入定影,最后洗去残液晾干,将胶片用Quantity One凝胶分析软件处理,测定各组蛋白条带的吸光度,以目的条带和β-actin条带的比值作为蛋白表达水平。每个蛋白样品重复3次。
1.2.4 流式细胞术检测细胞凋亡:用不含EDTA的胰酶消化各组细胞,离心收集各组细胞,PBS漂洗2次,加结合缓冲液重悬细胞。依据试剂盒说明书,先后加入Annexin V-FITC和PI避光孵育15 min。流式细胞仪检测激发波长488 nm和发射波长530 nm处的荧光强度。实验重复3次。
1.2.5 ELISA法检测TNF-α和IL-6的表达:取各组细胞,离心取上清,然后按照ELISA试剂盒说明书进行检测。
1.2.6 荧光素酶报告实验检测miR-32-5p对NFIL3的靶向调控:TargetScan数据库显示NFIL3 3′UTR区域有miR-32-5p结合位点。构建含有miR-32-5p结合位点的NFIL33′UTR野生型和突变型荧光素酶表达载体(WT-NFIL3和MUT-NFIL3),取对数生长期肺上皮细胞A549接种于24孔板(5×104个/孔),待细胞生长至80%融合时,用LipofectamineTM 2000将WT-NFIL3和MUT-NFIL3组细胞分别转染miR-con和miR-32-5p。依据说明书要求,使用荧光素酶报告基因检测仪进行双荧光素酶报告实验测定。实验结果以荧光素酶活性和Renilla活性的比值进行统计学分析。实验重复3次。

2 结果
2.1 LPS诱导的肺上皮细胞A549中miR-32-5p和NFIL3的表达水平 qRT-PCR和Western Blot检测结果显示,与NG组相比,LPS组A549细胞中miR-32-5p表达水平显著升高,NFIL3 mRNA和蛋白表达水平显著降低(P<0.05)。可见,LPS诱导的肺上皮细胞A549中miR-32-5p高表达,NFIL3低表达。见表1,图1。
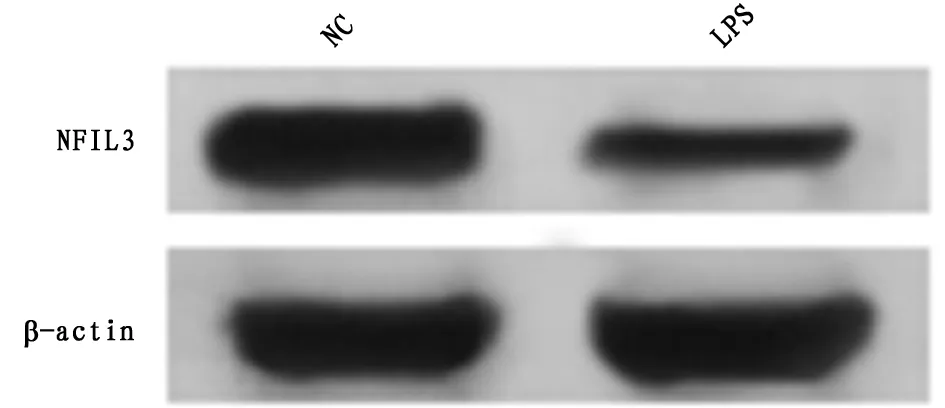
图1 NFIL3蛋白的表达

表1 miR-32-5p、NFIL3mRNA及NFIL3蛋白表达
2.2 miR-32-5p低表达抑制LPS诱导的肺上皮细胞凋亡和炎性反应 与NG组相比,LPS组A549细胞中miR-32-5p、cleave-caspase-3表达水平显著升高,TNF-α、IL-6含量显著升高,细胞凋亡率也显著升高(P<0.05);与LPS+anti-miR-con组相比,LPS+anti-miR-32-5p组A549细胞中miR-32-5p、cleave-caspase-3表达水平显著降低,TNF-α、IL-6含量显著降低,细胞凋亡率也显著降低(P<0.05)。可见,LPS可诱导肺上皮细胞凋亡和炎性反应,而miR-32-5p低表达可抑制LPS诱导的肺上皮细胞凋亡和炎性反应。见表2,图2。
表2 miR-32-5p低表达抑制LPS诱导的肺上皮细胞凋亡和炎性反应
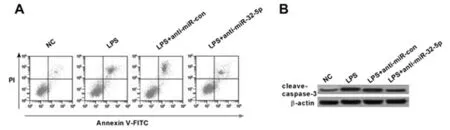
图2 miR-32-5p低表达抑制LPS诱导的肺上皮细胞凋亡和炎性反应;A 流式细胞仪检测细胞凋亡;B cleave-caspase-3蛋白的表达
2.3 高表达NFIL3抑制LPS诱导的肺上皮细胞凋亡和炎性反应 与NG组相比,LPS组A549细胞中cleave-caspase-3表达水平显著升高,NFIL3表达水平显著降低,TNF-α、IL-6含量显著升高,细胞凋亡率也显著升高(P<0.05);与LPS+pcDNA-con组相比,LPS+pcDNA-NFIL3组A549细胞中cleave-caspase-3表达水平显著降低,NFIL3表达水平显著升高,TNF-α、IL-6含量显著降低,细胞凋亡率也显著降低(P<0.05)。可见,高表达NFIL3可抑制LPS诱导的肺上皮细胞凋亡和炎性反应。见表3,图3。
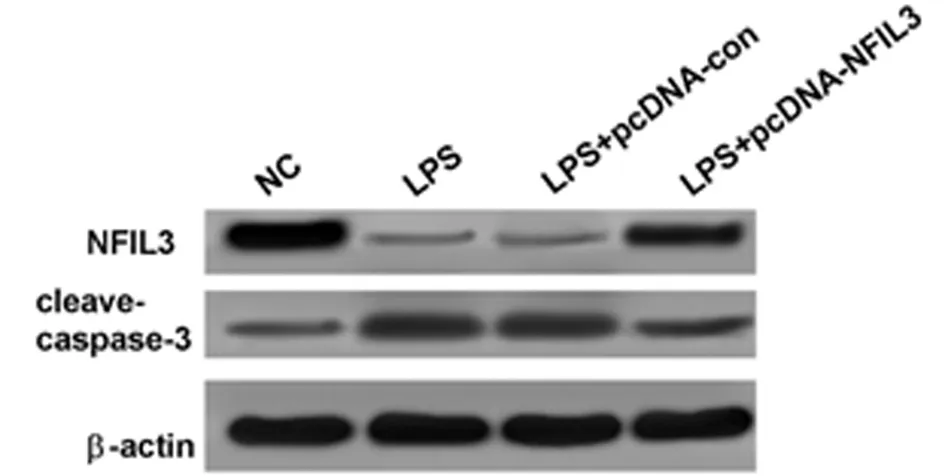
图3 Western Blot检测cleave-caspase-3、NFIL3蛋白的表达
表3 高表达NFIL3抑制LPS诱导的肺上皮细胞凋亡和炎性反应
2.4 miR-32-5p靶向NFIL3 通过TargetScan数据库预测到NFIL3与miR-32-5p存在结合位点。荧光素酶报告基因检测实验结果显示,转染NFIL3基因野生型和突变型表达载体后,相较于miR-con组,miR-32-5p组野生型肺上皮细胞的荧光素酶活性显著降低(P<0.05);而突变型肺上皮细胞的荧光素酶活性差异不
显著(P>0.05)。Western Blot检测结果显示,相较于miR-con组,miR-32-5p组肺上皮细胞中NFIL3表达水平显著降低;相较于anti-miR-con组,anti-miR-32-5p组肺上皮细胞中NFIL3表达水平显著升高(P<0.05)。可见,miR-32-5p可靶向调控NFIL3的表达。见表4、5,图4。
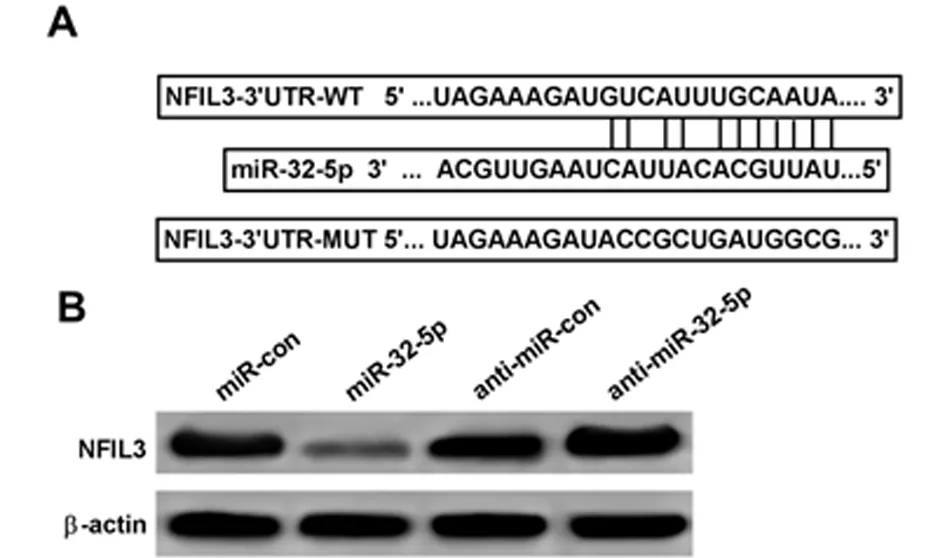
图4 miR-32-5p靶向调控NFIL3表达;A starbase对miR-32-5p和NFIL3结合进行预测示意图;B Western Blot检测NFIL3表达量
表4 miR-con或miR-32-5p与报告质粒共转染A549细胞后双荧光素酶活性检测
2.5 敲减NFIL3可以部分逆转anti-miR-32-5p对LPS诱导的肺上皮细胞凋亡和炎性反应的影响 与LPS+anti-miR-con组相比,LPS+anti-miR-32-5p组A549细胞中NFIL3表达水平显著升高,cleave-caspase-3表达水平显著降低,TNF-α、IL-6含量显著降低,细胞凋亡率显著降低(P<0.05);与LPS+anti-miR-32-5p+si-con组相比,LPS+anti-miR-32-5p+si-NFIL3组A549细胞中NFIL3表达水平显著降低,cleave-caspase-3表达水平显著升高,TNF-α、IL-6含量显著升高,细胞凋亡率显著升高(P<0.05)。可见,敲减NFIL3可以部分逆转anti-miR-32-5p对LPS诱导的肺上皮细胞凋亡和炎性反应的抑制作用。见表6,图5。
表5 Western Blot检测NFIL3的表达
表6 敲减NFIL3可以部分逆转anti-miR-32-5p 对LPS诱导的肺上皮细胞凋亡和炎性反应的影响
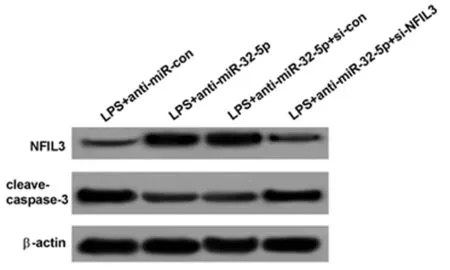
图5 Western Blot检测NFIL3、cleave-caspase-3蛋白表达
3 讨论
ALI为常见且复杂的炎症性肺疾病,ALI发病率和死亡率较高,主要的病理生理改变是弥漫性肺毛细血管内皮-肺泡上皮屏障功能的破坏及炎症损,减轻肺上皮细胞凋亡和炎性反应可减缓ALI的进展[8]。miRNA参与细胞增殖、分化、凋亡、脂肪代谢等生命过程[9],且研究表明ALI发病过程中miRNA表达异常,miRNA可通过与靶mRNA部分结合在转录和转录后水平调节靶基因表达,参与ALI的整个发病过程,miRNA在急性肺损伤中具有重要调节作用[10]。如脂多糖处理中miR-29b高表达,下调miR-29b可保护LPS诱导的大鼠ALI,减轻炎性反应[11]。miR-155反义寡核苷酸(ASO)可以降低IL-6和TNF-α水平,抑制炎性反应,保护内毒素诱导的小鼠ALI[12]。而miR-32-5p对ALI的影响还未见报道。研究发现下调miR-32-5p促进KLF4的表达增加了前列腺癌的化学敏感性,促进细胞凋亡[13]。敲低miR-32-5p极大抑制了大鼠神经性疼痛,并降低了炎性细胞因子IL-1β,TNF-α和IL-6蛋白的表达[14]。本实验结果显示,脂多糖处理促进miR-32-5p的表达,miR-32-5p低表达抑制cleave-caspase-3的表达,抑制脂多糖诱导的肺上皮细胞凋亡;且还降低IL-6、TNF-α的水平。与前人研究结果相似,说明miR-32-5p低表达可以减轻炎性反应,促进肺上皮细胞凋亡。
NFIL3是一种重要的负转录因子,它通过与IGF2R启动子结合,抑制缺氧条件下H9c2心肌细胞中IGF2R信号通路诱导的细胞凋亡[7]。NFIL3过度表达通过抑制TRAIL和拮抗过氧化氢诱导的细胞死亡来支持肿瘤细胞的存活[15]。抑制NFIL3消除了IL-5对地塞米松诱导的细胞凋亡的保护作用[16]。以上结果表明,过表达NFIL3促进细胞存活,抑制细胞凋亡。本实验结果显示,脂多糖处理抑制NFIL3的表达,高表达NFIL3抑制cleave-caspase-3的表达,抑制脂多糖诱导的肺上皮细胞凋亡。此外,CIK细胞中超表达NFIL3能有效减少细菌入侵,并能抑制入侵后细胞的增殖;其与显著激活NF-κB活性,提高干扰素IFN和白介素IL-10基因的表达有关[17]。而本研究中过表达NFIL3也降低了IL-6、TNF-α的水平。本实验还发现miR-32-5p靶向调控NFIL3的表达,敲减NFIL3可以部分逆转抑制miR-32-5p对脂多糖诱导的肺上皮细胞凋亡和炎性反应的抑制作用。提示,miR-32-5p可能通过调控NFIL3影响脂多糖诱导的肺上皮细胞凋亡和炎性反应。
综上所述,miR-32-5p可抑制脂多糖诱导的A549细胞凋亡和减轻炎性反应,其机制与上调NFIL3及抑制炎性因子TNF-α、IL-6的表达有关。可为ALI的治疗提供新思路和新靶点。
猜你喜欢
中老年保健(2021年9期)2021-08-24
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
广东蚕业(2021年1期)2021-03-18
中华养生保健(2020年1期)2020-11-16
山东医药(2020年9期)2020-05-20
中国神经免疫学和神经病学杂志(2019年3期)2019-01-04
药学研究(2015年11期)2015-12-19
医学研究杂志(2015年9期)2015-07-01
食品工业科技(2014年23期)2014-03-11
