
CTAB对乳腺癌细胞MCF-7和MDA-MB-231凋亡影响
2021-02-24 09:22张运秋,潘悦,陶旭锋,肖桂山
大连理工大学学报 2021年1期
张 运 秋,潘 悦,陶 旭 锋,肖 桂 山
( 大连理工大学 化工学院, 辽宁 大连 116024 )
0 引 言
乳腺癌的发病率和致死率位居高位,并且发病率逐年升高,这导致乳腺癌逐渐成为威胁女性健康的首位恶性肿瘤[1-3].2018年全球乳腺癌占新发癌症病例的24.2%,占癌症死亡病例的15.0%.在全球范围内,乳腺癌的发病率和死亡率分别在154个和104个国家排名第一[4],每4个女性癌症患者中就有1个是乳腺癌患者[5].乳腺癌治疗主要以外科手术和化疗为主[6-7].广大女性乳腺癌患者因手术治疗存在创伤面大、复发率高等缺点而选择化疗[8],然而大部分的患者经过长期化疗之后会出现耐药现象[9].这一现象严重影响了乳腺癌患者的生活质量及疾病预后[10-12],因此发现新的抗乳腺癌药物意义重大.
抗肿瘤药物杀伤肿瘤细胞的主要方式有诱导凋亡、影响细胞周期、自噬和坏死,而凋亡作为主要的方式之一,发挥着巨大的作用.细胞凋亡是细胞通过遗传编码进行自杀的过程,并且整个凋亡过程有着严格的调节程序[13-14].细胞凋亡与许多基因有关,其被相关基因严格把控着.作为一个与肿瘤增殖和凋亡密切相关的蛋白,p53的激活可以通过促进细胞凋亡起到抗肿瘤的作用[15].诱导线粒体内源性凋亡是抗肿瘤药物发挥抗肿瘤活性的主要机制之一[16-17],大量文献表明,p53可以激活Bax,从而打开线粒体通道使细胞色素C释放到胞浆中,进而激活下游的Caspase-9以及凋亡执行蛋白Caspase-3,从而激活线粒体内源性凋亡通路.除此之外,Bad作为Bcl-2家族的促凋亡基因之一,在肿瘤发生发展中起着重要的作用,Schimmer等[18]发现,Bad蛋白可以直接诱导凋亡.Bim是Bcl-2家族的重要凋亡调节因子,有研究表明DNA损伤会影响Bim的表达从而引起细胞的凋亡[19].
十六烷基三甲基溴化铵(CTAB)主要作为纳米材料合成过程中的打孔剂[20],与阴离子和非离子等均具有良好的配位性,这样的结构特性导致其也是较好的两性表面活性剂,并且具有良好的乳化、抗静电、杀菌及生物降解性.近期有研究表明以CTAB为首的一系列具有长碳链季铵盐结构的化合物具有抗肿瘤活性,其对结肠癌、肺癌和肝癌等均具有明显的治疗作用[21-22],低剂量CTAB能够增强乳腺癌的药物敏感性[23],但是CTAB是否能够具有抗乳腺癌的作用,还有待探索.因此,本研究拟初步探讨CTAB是否对乳腺癌细胞系MCF-7和MDA-MB-231的凋亡有影响,以及其是否影响这两种细胞系的内源性凋亡信号通路.
1 材料与方法
1.1 材料与仪器
1.1.1 细胞系 乳腺癌细胞系MCF-7和MDA-MB-231购自美国ATCC细胞库.
1.1.2 主要试剂与仪器 CTAB(H9151-100g,纯度≥99%)购自美国Sigma公司,FBS、DMEM高糖培养基、Trypsin-EDTA胰酶购自美国Gibco公司.青链霉素混合液(100×)、MTT(1g)试剂、BCA蛋白浓度测定试剂盒购自北京索莱宝生物科技有限公司.AV-PI双染色凋亡试剂盒购自沈阳万类生物科技公司.RIPA裂解液、Western Blot显影液购自北京普利莱公司.DMSO购自天津市富宇精细化工有限公司,蛋白免疫印迹实验相关化学试剂购自天津大茂化学试剂厂.Tween20购自美国Sigma公司.β-actin、p53、Caspase-3、Caspase-9、Bax、Bim、Bcl-2、P-bcl-2和Bad一抗均购自美国Cell Signaling Technology(CST)公司,二抗羊抗兔和羊抗鼠购自美国Proteintech 公司.其他仪器包括生物安全柜购自美国Thermo公司、酶标仪购自美国Bio TeK公司、荧光显微镜购自日本奥森巴斯公司、Life Attune 声波聚焦流式细胞仪购自美国Thermo Fisher公司、凝胶成像仪购自美国伯乐公司.
1.2 实验方法
1.2.1 细胞培养 乳腺癌细胞系MCF-7、MDA-MB-231采用含10%FBS和1%双抗的DMEM高糖培养基,置于37 ℃、5%CO2的培养箱中培养.每隔2 d换一次液,细胞密度达到80%~90%时传代培养或用于其他实验.
1.2.2 细胞活性MTT检测 用胰酶适度消化生长状态良好的MCF-7、MDA-MB-231细胞,然后用FBS终止消化,离心弃去胰酶和FBS的混合液,用PBS冲洗离心后弃上清,用新的培养基将细胞按照5 000/孔的密度接种到96孔板中,每孔200 μL培养液.过夜培养待细胞贴壁后,按照设定的给药浓度梯度进行给药.设置每个浓度梯度6个复孔,重复铺两板,在分别给药24 h和48 h后,每孔加入20 μL MTT溶液(5 mg/mL),在37 ℃ 孵箱孵育4 h后,抽出每孔中含有MTT的培养液,然后每孔加入150 μL DMSO,将96孔板放在摇床上振荡10 min,用酶标仪检测490 nm波长下每组的吸光值,用GraphPad Prism 5.0计算不同浓度药物对细胞的抑制率以及药物的IC50值.
1.2.3 细胞形态的观察 待MCF-7、MDA-MB-231细胞生长至对数期,取IC50值(相近的浓度值)进行给药,对照组给同等体积的空白溶剂,给药24 h后,用PBS轻轻冲洗掉死细胞,放置显微镜下观察细胞形态的变化并在光学显微镜(200×)下观察拍照.
1.2.4 细胞凋亡率的检测 培养MCF-7、MDA-MB-231细胞生长至对数期,根据MCF-7、MDA-MB-231细胞的IC50值,设定MCF-7的给药浓度为0.75、1.5、3 μg/mL,设定MDA-MB-231的给药浓度为0.5、1.0、1.5 μg/mL,给药24 h后,收集用CTAB处理的细胞及空白对照组的细胞,用不含EDTA的胰蛋白酶消化细胞,消化时间不宜过长以避免假阳性,消化适当时加入完全培养基终止消化反应,用PBS清洗两次得到干净的细胞,然后根据说明书对细胞进行染色,在1 h内用流式细胞仪检测细胞凋亡率.
1.2.5 Western Blot检测相关蛋白表达 用胰酶消化待提取的细胞,PBS清洗2遍,离心留取细胞,加入RIPA裂解液和PMSF的混合液(混合比例50∶1),置于冰上孵育10 min,每隔5 min涡旋振荡30 s,离心后留取上清液保存于-80 ℃.使用BCA蛋白浓度检测试剂盒定量蛋白浓度,按比例加入5×上样缓冲液后95 ℃、10 min,保存于-20 ℃冰箱.每条泳道加入30 μg蛋白样品,设置恒压70 V约30 min跑完浓缩胶,然后调至恒压140 V跑完分离胶.切取目标蛋白条带转移至甲醇活化好的PVDF膜上,转膜恒流300 mA约90 min,牛奶封闭1 h,用TBST清洗膜3次(5 min/次),加入一抗(1∶1 000稀释)4 ℃ 孵育过夜,再用TBST清洗3次(10 min/次),室温孵育二抗1 h,TBST清洗3次(10 min/次),将膜用ECL高敏显影液避光孵育2 min,然后置于凝胶成像仪中显影.
1.3 统计学分析
用GraphPad Prism 5.0对数据进行处理,结果以均值±标准差表示,两组间差异比较采用t检验,以P<0.05为有显著性差异.
2 实验结果
2.1 CTAB对乳腺癌细胞的杀伤作用
CTAB化学结构如图1所示,本研究选取两种乳腺癌细胞系MCF-7和MDA-MB-231,经预实验将6.25 μg/mL设定为MCF-7细胞系的最大给药终浓度,按照2/3梯度稀释依次设定浓度梯度(0、0.37、0.55、0.82、1.23、1.85、2.78、4.17、6.25 μg/mL).同理,经预实验将1.56 μg/mL设定为MDA-MB-231细胞系的最大给药终浓度,按照4/5梯度稀释依次设定浓度梯度(0、0.33、0.41、0.51、0.64、0.80、1.00、1.25、1.56 μg/mL).用以上设定浓度分别处理乳腺癌细胞系24 h和48 h,结果显示CTAB对乳腺癌细胞有明显的杀伤作用(图2,P<0.05),经计算作用24 h,MCF-7的IC50是1.32 μg/mL,MDA-MB-231的IC50是0.82 μg/mL(表1).因此,CTAB对于乳腺癌细胞确实具有一定的杀伤作用.
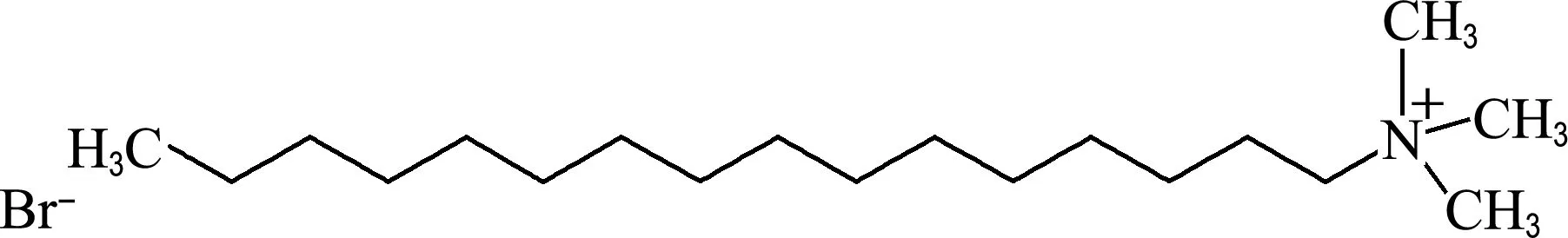
图1 CTAB的化学结构
2.2 CTAB对乳腺癌细胞形态的影响
为了更直观地观察CTAB对乳腺癌细胞的影响,本研究分别选用两种细胞系的IC50值(附近浓度值)1.5 μg/mL和1.0 μg/mL处理MCF-7和MDA-MB-231细胞系,24 h后观察细胞形态是否发生变化.实验结果表明,CTAB能够引起乳腺癌细胞形态的明显变化,与对照组相比,给药后细胞变细变长(图3),说明给药后细胞状态不是很好,可能是发生了早凋现象.由此本研究得出CTAB能够引起乳腺癌细胞形态发生明显变化,推测CTAB可能能够引起乳腺癌细胞发生凋亡现象.

表1 CTAB在不同时间下对乳腺癌细胞的IC50值
2.3 CTAB对乳腺癌细胞凋亡诱导作用
为了进一步明确CTAB杀伤乳腺癌细胞的作用机制,本研究用AV-PI双染的方式检测CTAB对乳腺癌细胞系的作用.根据MCF-7细胞给药24 h的IC50值,设定给药浓度为0.75、1.5、3 μg/mL;根据MDA-MB-231细胞给药24 h的IC50值,设定给药浓度为0.5、1.0、2.0 μg/mL.结果表明,CTAB杀伤乳腺癌细胞可能是通过诱导其凋亡实现的,随着CTAB的浓度增加,其凋亡率也明显增加(图4、5).用0.75、1.5、3 μg/mL的CTAB处理MCF-7细胞24 h后,其凋亡率分别为5.12%、8.68%和22.21%(图4);用0.5、1.0、2.0 μg/mL的CTAB处理MDA-MB-231细胞24 h 后,其凋亡率分别为8.83%、10.19%和17.75%(图5).以上结果表明,CTAB能够一定程度上诱导乳腺癌细胞系MCF-7和MDA-MB-231凋亡.
2.4 CTAB诱导乳腺癌细胞线粒体内源性凋亡机制
为了研究CTAB诱导乳腺癌细胞凋亡的作用机制,本研究用1.5 μg/mL CTAB处理MCF-7,24 h,检测细胞中p53蛋白表达量.结果显示,经CTAB处理的MCF-7细胞中p53蛋白的表达量R明显增加(图6),因此本研究认为CTAB可能通过激活p53来诱导乳腺癌细胞凋亡.在化疗药物引起的凋亡里,线粒体内源性凋亡一直扮演着非常重要的角色.为了进一步研究CTAB诱导乳腺癌细胞凋亡的方式,本研究检测了线粒体内源性凋亡的相关蛋白Bcl-2、P-bcl-2、Bax、Bad、Bim、Caspase-9、Cleaved Caspase-9、Caspase-3以及Cleaved Caspase-3在MCF-7细胞中给药前后的表达量.结果显示,CTAB给药24 h内抑制Bcl-2蛋白的磷酸化,并且激活与线粒体内源性凋亡相关的促凋亡蛋白Bax、Caspase-9、Cleaved Caspase-9、Caspase-3、Cleaved Caspase-3、Bad以及Bim(图6).因此本研究提出CTAB可能是通过激活p53诱导乳腺癌细胞线粒体内源性凋亡.
3 结果讨论
目前有文献表明CTAB对p53野生型结肠癌具有更好的抗肿瘤活性.因此本研究猜测CTAB的抗肿瘤作用可能与p53相关[14-15].本研究采用Western Blot法的实验结果显示CTAB处理MCF-7细胞24 h后p53表达量明显增加,证明了CTAB确实与p53相关,且CTAB能够促进p53蛋白的表达,更加直接明了地验证了CTAB能够激活p53这一关系.
Bcl-2家族在调节线粒体内源性凋亡的通路中发挥着重要的作用,该家族中和凋亡相关的成员众多,其中包括本研究中所探索的Bax等促凋亡蛋白和Bcl-2等抗凋亡蛋白[24],文献显示,Bcl-2/Bax可调控细胞凋亡并在乳腺癌治疗中发挥着非常重要的作用[25].叶惠荣等[26]研究表明,灵菌红素通过上调Bim抑制人乳腺癌MCF-7细胞增殖并促进其凋亡.梁欢等[27]研究显示维生素D可以上调乳腺癌细胞的促凋亡蛋白Bax的表达,下调抗凋亡蛋白Bcl-2的表达从而促进乳腺癌细胞的凋亡.本研究采用一系列浓度梯度CTAB处理人乳腺癌MCF-7和MDA-MB-231细胞24 h和48 h后,发现随着CTAB浓度增加,细胞增殖明显被抑制;流式细胞技术检测结果显示,MCF-7和MDA-MB-231细胞凋亡率随着CTAB浓度梯度逐渐增加.并且经CTAB处理后,乳腺癌细胞系MCF-7中的Bax、Bad、Bim等蛋白表达量升高,P-bcl-2的表达量降低.Caspase-3、Cleaved Caspase-3、Caspase-9及Cleaved Caspase-9在细胞的再生与凋亡代谢中发挥着关键作用.本研究中,CTAB用于MCF-7细胞后,Cleaved Caspase-3及Cleaved Caspase-9表达均较对照组增加,提示CTAB通过促进线粒体内源性凋亡相关蛋白表达诱导MCF-7细胞凋亡.
目前越来越多的纳米材料被应用于抗肿瘤的治疗,近期许多研究表明CTAB在一定的剂量范围内,也可以起到抗肿瘤作用,同时还具有一定的选择性杀伤作用,有一定的临床应用价值[28],因此本研究的实验结果初步证明CTAB具有良好的抗乳腺癌作用,为其应用于未来乳腺癌的治疗提供了一定的理论依据.但是本研究没有进一步具体验证在CTAB促进人乳腺癌细胞MCF-7和MDA-MB-231凋亡的进程中,p53和线粒体内源性凋亡相关蛋白的具体作用关系,有待进一步研究.
4 结 语
本文实验结果证明CTAB是通过激活p53抑制Bcl-2的磷酸化,增加Bax、Bad以及Bim蛋白的表达量,进而打开线粒体通道激活下游的Caspase-9以及凋亡执行蛋白Caspase-3激活线粒体内源性凋亡通路,从而诱导乳腺癌细胞凋亡.
猜你喜欢
中国医学物理学杂志(2021年10期)2021-11-04
昆明医科大学学报(2021年8期)2021-08-13
科学(2020年3期)2020-11-26
哈尔滨轴承(2020年2期)2020-11-06
湖南大学学报·自然科学版(2019年6期)2019-07-26
决策与信息·下旬刊(2015年8期)2015-10-22
现代养生·下半月(2015年6期)2015-09-07
医学研究杂志(2015年4期)2015-06-10
中国医药导报(2015年26期)2015-02-28
现代检验医学杂志(2015年6期)2015-02-06
