
小麦TaGSK5基因的克隆与表达分析
2021-02-24 08:00张令慧张沛沛刘媛陈涛王彩香程宏波杨德龙
甘肃农业大学学报 2021年6期
张令慧,张沛沛,刘媛,陈涛,王彩香,程宏波,杨德龙
(1.甘肃省干旱生境作物学省部共建国家重点实验室培育基地,甘肃 兰州 730070;2.甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
小麦(Triticumaestivum)是我国四大主粮作物之一,主要分布在北方雨养农业区.随着全球气候的变化,淡水资源日益匮乏,土壤盐渍化加剧,小麦在发育时期更易遭受干旱、低温、干热风、盐渍化等非生物胁迫,致使小麦生长发育异常,产量和品质显著下降[1].为应对各种非生物胁迫的危害,小麦在长期的进化过程中形成了一系列的生理生化调控机制[2-4].许多基因在小麦非生物逆境胁迫应答中发挥着重要的作用[5-6].因此,深入发掘优异的抗逆基因,解析其生物学功能,对小麦抗逆分子遗传改良具有重要意义.
糖原合成激酶(glycogen synthase kinase 3/shaggy like kinase,GSKs)是一类保守的丝氨酸/苏氨酸蛋白激酶家族[7],在响应植物非生物胁迫和参与植物生长发育调控方面起重要作用.近几年,GSKs基因相继在拟南芥(Arabidopsisthaliana)[8]、苜蓿(Medicagosativa)[9]、棉花(Gossypiumspp.)[10]和水稻(Oryzasativa)[11]中被发现与克隆.研究发现,GSK家族成员蛋白的激酶结构域高度保守的[12],在水稻和拟南芥GSK-like激酶结构域的结构相似性达75%[13].GSKs能够磷酸化多个下游底物来发挥生物学功能[14].BR信号通路的GSK3的同系物BIN2是通过磷酸化和去磷酸化控制BR信号传导下游的转录因子BRZ1和BES1/BZR2来阻止信号转导,通过负向调节作用进而诱导其他生化反应[15],而拟南芥ASKa是通过激活Glu-6-磷酸脱氢酶(G6PD)来调节应激耐受性[16].植物中的GSKs是一个多基因家族[17],在拟南芥中已经鉴定出10个GSKs家族成员[18],水稻中包含9个[19].进化分析表明,植物GSKs家族成员可分为Ⅳ个亚家族[20],其中已经被鉴定的第Ⅳ亚家族基因AtSK11[21]、OsGSK5[22]和MsK4[9]受非生物胁迫诱导并且具有激酶活性.在拟南芥中,AtSK11受NaCl、干旱、ABA、热诱导上调表达[21],研究发现高盐胁迫能够快速诱导糖原合成激酶MsK4上调表达[9].水稻OsGSK5通过调节碳水化合物的代谢平衡从而调高了植物对盐胁迫的耐受性[22].上述研究表明,植物GSKs的第Ⅳ亚家族成员在逆境胁迫响应中发挥重要作用.然而,截止目前有关小麦GSK基因的克隆和响应非生物胁迫的研究尚未见报道.因此,本研究拟克隆小麦TaGSK5基因并对其序列进行生物信息学分析和表达分析,以期为进一步研究小麦TaGSK5基因生物学功能提供理论依据.
1 材料与方法
1.1 试验材料
选取陇中1号为供试材料,该品种适宜在甘肃兰州和陇东地区、宁夏固原地区、陕西延安地区的旱地冬麦区种植.陇中1号在本实验室基地种植抗旱效果较好.
1.2 处理方法
挑选籽粒大小一致、圆润饱满的陇中1号小麦种子置于培养皿上,4 ℃黑暗处理2 d后置于人工气候室内,在光照强度为62.5 μmol/(m2·s),光周期为16 h/8 h,温度为25 ℃条件下进行培养.待幼苗长至两叶一心时(14 d),对小麦幼苗进行非生物胁迫处理.将采用水培法培养的小麦幼苗置于20% PEG-6000溶液和200 mmol/L的NaCl的溶液中培养,分别在处理后0、1、3、6和12 h的时间点采集小麦幼苗各组织,液氮速冻后,-80 ℃超低温保存备用.
1.2.1TaGSK5基因cDNA的引物设计 以水稻OsGSK5的cDNA为参考序列(XP_015633107),在小麦数据库网站(http://202.194.139.32/blast/viroblast.php)中检索其在小麦中的同源基因,通过序列比对获得小麦TaGSK5的4个同源候选基因,命名为TaGSK5-1A,TaGSK5-4A,TaGSK5-5B及TaGSK5-5D.利用Primer premier 5.0软件设计通用引物进行小麦陇中1号的TaGSK5基因扩增:引物序列为TaGSK5-F(5′→3′):CAATCAGCTAGGAGATGGCGTA和TaGSK5-R(5′→3′):CACCCACACAAAGCTACAGCT,引物由生工生物工程(上海)股份有限公司合成.

1.2.4 生物信息学分析 运用ExPASy-ProtParam对小麦TaGSK5的二级结构及蛋白的理化性质进行预测;运用ExPASy-ProtParam构建TaGSK5蛋白的三级结构;使用WoLFPsort (www.gens cript.com/wolf-psort.html)预测TaGSK5的亚细胞定位;在NCBI网站搜索不同物种的GSK同源蛋白并利用DNAMAN软件进行氨基酸序列比对;使用STRING(https://string-db.org/cgi/input)预测TaGSK5蛋白的互作网络;采用MEGA 7.0软件构建小麦、水稻、拟南芥、苜蓿、大麦和粗山羊草等8个物种GSK同源蛋白的系统进化树;在WheatExp(https://wheat.pw.usda.gov/ WheatExp/)数据库下载小麦不同组织(根、茎、幼穗、叶和籽粒)TaGSK5基因的表达谱数据,利用TBtools软件绘制表达谱数据热图.
2 结果与分析
2.1 TaGSK5基因cDNA的克隆及基因结构
以小麦陇中1号叶片的cDNA为模板,使用特异性引物TaGSK5-F和TaGSK5-R扩增目的片段,扩增产物经电泳检测,在1 200 bp左右出现1个特异性条带(图1).将目的条带回收纯化、转化大肠杆菌,挑选阳性克隆测序验证.测序结果与中国春的4条序列进行比对.结果表明,不同单克隆菌的序列信息与1A、4A、5B和5D染色体上中国春释放的序列信息相一致,将其命名为TaGSK5.所得到的4个TaGSK5基因cDNA序列,分别位于小麦的1A、4A、5B和5D的染色体上,故将其分别命名为TaGSK5-1A、TaGSK5-4A、TaGSK5-5B和TaGSK5-5D.通过基因结构分析发现,TaGSK5的4个部分同源基因都编码426个氨基酸且均由12个外显子和11个内含子组成(图2).

M:DL5000 DNA marker;1:TaGSK5基因的扩增产物.M:DL5000 DNA marker;1:The amplification products of TaGSK5 gene.图1 TaGSK5基因cDNA序列扩增Figure 1 The cDNA amplificated products of TaGSK5 gene

图2 TaGSK5的基因结构Figure 2 The gene structure of TaGSK5
2.2 TaGSK5基因编码的氨基酸序列比对及进化树分析
使用DNAMAN对小麦TaGSK5与水稻(XP_015633107)、苜蓿(AF432225)和拟南芥(At1g 09840)的氨基酸序列进行多重比对(图3),结果表明,小麦TaGSK5的4个同源基因氨基酸相似性达到99.43%,4个物种间的同源性在70%以上,TaGSK5与水稻相似性较高(>90%),与苜蓿和拟南芥的同源性较低.他们的GSK激酶(88-375)结构域高度相似,其中的功能基序TREE、CDFGSAK和SYICSR的序列较一致,表明TaGSK5蛋白在进化过程中具有较强的保守性.进化分析表明,TaGSK5的4个部分同源基因和粗山羊草处于同一个分支(图4),说明祖先种来源于粗山羊草.
2.3 TaGSK5蛋白二级及三级结构预测
通过(http://bioinf.cs.ucl.ac.uk/)在线软件预测了TaGSK5的4个同源蛋白的二级结构组成,结果表明这4个蛋白二级结构的比例均由α-螺旋、无规则卷曲、延伸链和β-折叠构成(表1).TaGSK5-5B、TaGSK5-5D、TaGSK5-4A和TaGSK5-1A蛋白的二级结构α-螺旋分别占36.71%、36.71%、36.85%和37.32%,无规则卷曲分别占42.72%、40.24%、42.02%和40.85%,延伸链分别占15.26%、16.24%、15.73%和15.49%,β-折叠分别占6.34%、6.82%、5.40%和6.34%.通过(https://web.expasy.org/protparam/)在线软件构建TaGSK5蛋白的三级结构,因TaGSK5的4个同源蛋白的氨基酸序列高度相似,同源建模选用的蛋白模型相同,故使用TaGSK5-5B蛋白序列构建三级结构(图5).由图5可知,TaGSK5-5B蛋白的结构两端是由少部分的β-折叠,中间是α-螺旋由无规则卷曲串联起来的空间结构,主要是以α-螺旋和β-折叠形成的特殊结构来行使激酶功能.

表1 TaGSK5蛋白的二级结构预测

黑色方框:GSK蛋白激酶催化结构域;绿色背景:ATP结合位点;黄色背景和灰色背景:分别为高度保守基序CDFGSAK和SYICSR,仅存在于丝氨酸/苏氨酸蛋白激酶GSK-3亚家族成员;红色背景:植物中的保守基序TREE.Black box:GSK protein kinase catalytic domain;Green background:ATP binding site;Yellow and gray backgrounds shows highly conserved motifs CDFGSAK and SYICSR,respectively,which are only present in the members of the GSK-3 subfamily of serine/threonine protein kinases;Red background:TREE is the conservative motif in plants.图3 小麦TaGSK5与其他物种同源蛋白的氨基酸序列比对Figure 3 Alignments of amino acid sequences of wheat TaGSK5 with homologous proteins of other species

图4 小麦与其他物种GSK蛋白的进化分析Figure 4 Phylogenetic tree of GSK from wheat and other species

图5 TaGSK5蛋白的三级结构图Figure 5 Prediction of the tertiary structure of TaGSK5
2.4 TaGSK5蛋白的理化性质、亲疏水性及磷酸化位点分析
利用EXPASY-ProtParam在线网站预测TaGSK5蛋白的理化性质(表2).结果显示,这4个同源蛋白的理化性质非常接近.TaGSK5-5D、TaGSK5-5B、TaGSK5-4A和TaGSK5-1A的等电点非常接近,等电点分别为8.53、8.57、8.79和8.54;分子量分别为48 226.51、47 602.69、48 237.52和48 314.58.通过EXPASY-ProtParam网站预测TaGSK5蛋白质的亲疏水性,亲水性的氨基酸明显多于疏水性蛋白,表明TaGSK5蛋白质为亲水性蛋白(图6).利用EXPASY-ProtParam网站预测小麦TaGSK5蛋白磷酸化位点,发现TaGSK5蛋白有丝氨酸位点13个、苏氨酸位点8个、酪氨酸位点6个.表明该蛋白可能通过不同的磷酸化底物实现基因的调控功能(图7).
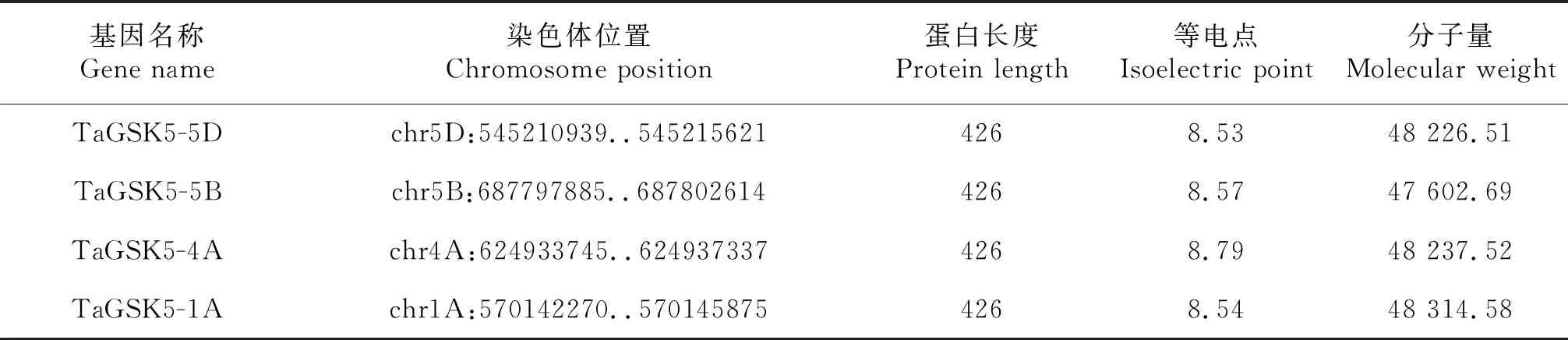
表2 TaGSK5基因编码的蛋白质的理化性质

垂直线零点以下代表相对的亲水区,零点以上为疏水区.The vertical line below the zero point represents the relative hydrophilic area,and the vertical line above the zero point represents the hydrophobic area.图6 TaGSK5蛋白的亲疏水性预测Figure 6 Prediction of hydrophilicity and hydrophobicity of TaGSK5

图7 TaGSK5蛋白磷酸化位点预测Figure 7 Prediction of phosphorylation site of TaGSK5
2.5 TaGSK5蛋白的亚细胞定位及互作蛋白网络
利用WOLFPSORT在线软件预测分析TaGSK5蛋白亚细胞定位,这4个同源蛋白主要分布在细胞质、细胞核、线粒体和过氧化物酶体中,但在细胞核和线粒体中的比例较高(表3),且TaGSK5蛋白没有信号肽结构,表明该蛋白不属于分泌蛋白.利用STRING在线软件对TaGSK5的互作蛋白预测(图8)发现,与TaGSK5互作的蛋白中有4个丝氨酸/苏氨酸特异性蛋白磷酸酶、2个cGMP依赖性蛋白激酶、1个具有Kelch样重复结构域的蛋白磷酸酶、1个PKG_like.1-ACG激酶和1个蛋白磷酸酶.

表3 TaGSK5的亚细胞定位预测
2.6 TaGSK5基因的表达特性分析
通过分析WheatExp数据库中TaGSK5基因在不同组织的表达谱数据,发现TaGSK5基因在小麦不同组织中均有表达,茎和穗中表达高于其他组织,在穗中的表达量最高(图9).通过qRT-PCR检测发现,小麦幼苗在干旱胁迫处理0~6 h时该基因表达量显著下调,在6~12 h时该基因的表达量上升.在NaCl处理下TaGSK5基因的表达量没有发生显著变化(图10).
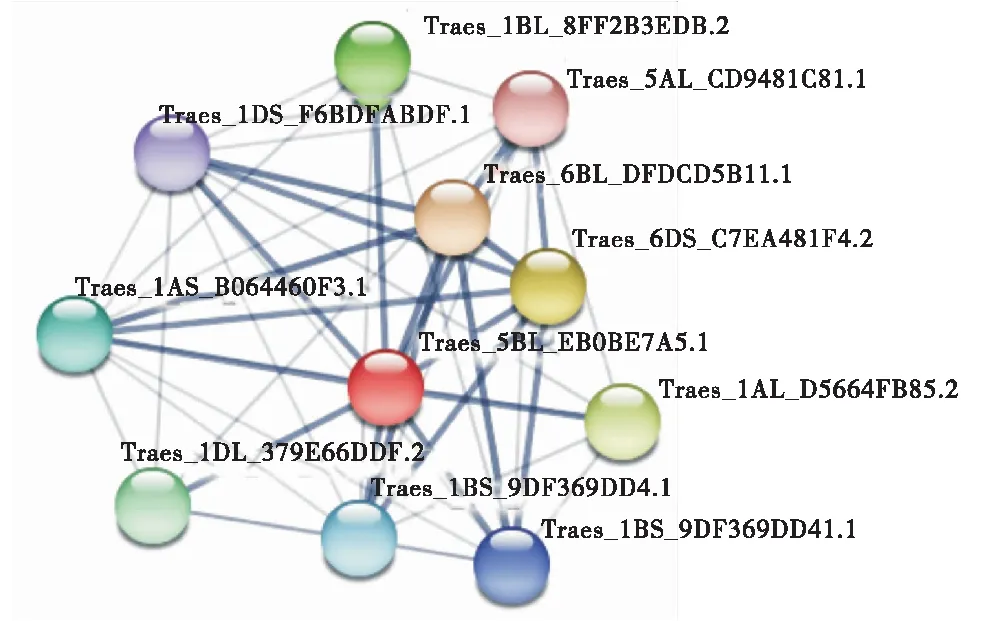
Traes_1DL_379E66DDF.2和Traes_1BL_8FF2B3EDB.2:拓扑异构酶1相关因子1;Traes_1AS_B064460F3.1、Traes_1DS_F6BDFABDF.1和Traes_1BS_9DF369DD41.1:Kelch重复域的蛋白磷酸酶;Traes_6BL_DFDCD5B11.1:AGC_PKA / PKG_like.1-ACG激酶;Traes_5AL_CD9481C81.1:丝氨酸/苏氨酸蛋白磷酸酶;Traes_6DS_C7EA481F4.2:蛋白磷酸酶2C;Traes_1AL_D5664FB85.2,Traes_1BS_9DF369DD4.1与Traes_1BS_9DF369DD41.1:含八糖肽或四三肽重复(TPR)的蛋白质.Traes_1DL_379E66DDF.2 and Traes_1BL_8FF2B3EDB.2:Topoisomerase 1 related factor 1;Traes_1AS_B064460F3.1,Traes_1DS_F6BFABDF.1 and Traes_1BS_9DF369DD41.1:Kelch repeat domain protein phosphatase;Traes_6BL_DFDKG_PKGC5B11.1:AGC_PKA / PKG_like.1-ACG kinase;Traes_5AL_CD9481C81.1:serine/threonine protein phosphatase;Traes_6DS_C7EA481F4.2:protein phosphatase 2C;Traes_1AL_D5664FB85.2,Traes_1BS_9DF369DD4.1 and Traes_1BS_9DF369DD41.1:proteins containing octa-glycopeptide or Tetratrieptide Repeat (TPR) domain.图8 TaGSK5蛋白的互作网络Figure 8 Interaction network of TaGSK5

G:籽粒;L:叶片;R:根部;S:穗部;ST:茎部.G:Grain;L:Leaf;R:Root;S:Spike;ST:Stem.图9 TaGSK5基因在不同组织中的表达量Figure 9 TaGSK5 gene expression in different tissues
3 讨论
糖原合酶激酶3(GSK3)是一种丝氨酸/苏氨酸激酶,是一种多功能激酶[7].大多数已经被克隆出的GSK基因参与非生物胁迫响应[22-23].本研究从小麦陇中1号中克隆了4个TaGSK5的cDNA序列,序列长度1 281bp.在水稻中,OsGSK5包含12个内含子和11个外显子[6],与在本研究中TaGSK5的4个同源基因的基因结构一致.他们所编码的蛋白无论从基因结构、理化性质还是蛋白结构域都高度相似[22].氨基酸序列比对显示,TaGSK54个部分同源基因所编码的蛋白的序列相似性达98%,与拟南芥、水稻和苜蓿中同源基因具有很高的相似性[9,11,21].本研究发现小麦TaGSK5的4个部分同源基因的激酶结构域高度保守,其中TREE结构域在不同物种中都存在[11].最近研究发现,GSK激酶结构域在不同植物中的功能基序高度相似,拟南芥bin2-1突变体出现矮化的表型是由于TREE基序突变,抑制了GSK蛋白激酶的活性所引起[11],表明功能基序在植物中激活激酶活性及蛋白功能的发挥具有作用.

A:PEG处理;B:NaCl处理.图柱上不同字母表示差异显著(P<0.05).A:PEG treatment;B:NaCl treatment.Different letters on the bar indicate significant differences (P<0.05).图10 在PEG和NaCl处理下TaGSK5的差异表达量Figure 10 Differential expression of TaGSK5 under PEG and NaCl treatment
进一步研究表明,GSK基因通过磷酸化下游蛋白来调控植物的生理过程[23-24].在水稻中的OsSK22介导OsBZR1磷酸化从而调控BR信号通路[25],本研究预测出TaGSK5蛋白有13个丝氨酸、8个苏氨酸和6个酪氨酸磷酸化位点,推测小麦TaGSK5蛋白通过磷酸化下游底物来调控不同的生物学过程.基因表达谱分析显示,TaGSK5在小麦的根、茎、叶和穗中均有表达,但在不同组织间的表达水平存在显著差异,其中在茎和穗中的表达量较高,而在叶和根中表达量较低.不同物种GSK基因在非生物胁迫处理下基因表达量也不尽相同.Wang等[10]报道了棉花中的第四亚家族成员GhSK42在盐胁迫处理下没有显著性的变化,而苜蓿MsK4和水稻OsGSK5过表达后分别提高了苜蓿和水稻对非生物胁迫的耐受性[9,22],本研究发现TaGSK5基因在干旱胁迫下表达量逐渐降低,而在盐胁迫下表达量没有显著变化.结果说明,不同植物中的GSK5基因在非生物胁迫中的响应机制不尽相同,本研究推测小麦TaGSK5基因负调控小麦耐旱性.关于该基因在非生物胁迫中的具体调控机制,还有待于进一步的研究.
4 结论
本研究克隆出的小麦TaGSK5基因,包含4个部分同源基因,其基因结构、保守结构域及蛋白的理化性质非常相似.TaGSK5基因在不同组织具有不同表达模式,在干旱胁迫TaGSK5下调表达,不响应盐胁迫,研究结果为进一步探究小麦TaGSK5基因的功能提供了理论依据.
猜你喜欢
汉字汉语研究(2021年2期)2021-08-30
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
国际放射医学核医学杂志(2020年4期)2020-07-27
汉字汉语研究(2019年2期)2019-08-27
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
上海农业学报(2017年3期)2017-04-10
河北书画研究(2016年3期)2016-04-28
中国医药生物技术(2015年4期)2015-12-26
天然产物研究与开发(2014年6期)2014-04-27
