
iTRAQ蛋白质组学分析揭示西瓜响应CGMMV胁迫的蛋白质
2021-02-22 08:48张慧青孙玉燕那冬晨
核农学报 2021年3期
张慧青 孙玉燕 那冬晨 范 敏
(1山西师范大学生命科学学院,山西 临汾 041000;2浙江省农业科学院蔬菜研究所,浙江 杭州 310021)
黄瓜绿斑驳花叶病毒(Cucumber green mottle mosaic virus,CGMMV)属于芜菁花叶病毒科烟草花叶病毒属。自1935年在英国首次被报道以来[1],已有30 多个国家和地区在葫芦科作物生产中发现CGMMV,对葫芦科作物的品质及产量造成严重威胁[2-7]。西瓜感染CGMMV 后,叶片出现浓绿色凹凸斑,茎部出现褐色病斑,果实发生畸形,果肉出现空洞、腐烂且呈暗红色[8]。近年来,CGMMV 传播速度迅速加快,对葫芦科作物尤其是西瓜产业的商业育种、育苗培育、种子贸易、产品生产和零售等带来全球性危害[8]。因此,研究西瓜对CGMMV 的响应,将为阐明其分子机理及后续提高西瓜对CGMMV 的抗性奠定基础。
关于西瓜感染CGMMV 转录和转录后调控的研究已取得一定进展[9-13]。通过对CGMMV 侵染前后的西瓜果实进行转录组分析,发现一些与细胞壁成分和光合作用相关的差异表达基因(differentially expressed genes, DEGs)可能参与CGMMV 病害症状的产生[9]。对CGMMV 侵染前后的西瓜叶片进行转录组分析,共鉴定到1 641 个DEGs,这些DEGs 主要参与光合作用、植物-病原菌互作、次生代谢和植物激素信号转导等代谢途径[10]。对CGMMV 侵染前后的西瓜叶片进行DNA 甲基化分析,发现RdDM(RNA-directed DNA methylation)介导的CHH 甲基化水平降低在西瓜抗病毒防御中发挥重要作用[11]。通过对感染CGMMV 前后的西瓜叶片进行小RNA(micro RNAs,miRNAs)测序,获得多个参与CGMMV 胁迫应答的miRNAs,包括miR164、 miR319 和miR393[12]。对西瓜叶片中CGMMV衍生的短干涉RNA(short intering RNAs,siRNAs)进行研究,发现这些siRNAs 的长度多为21 或22 nt,主要来源于病毒正义链,siRNAs 作用的宿主转录本包括MYB、WD40、bHLH、真核翻译起始因子、细胞色素P450 和纤维素合酶等,表明siRNAs 在病毒-宿主相互作用过程中发挥重要作用[13]。在蛋白质水平上研究西瓜对CGMMV 的胁迫应答,可进一步挖掘CGMMV 胁迫应答的蛋白质并明确其蛋白互作网路。蛋白质组学是研究植物-病毒相互作用和病毒侵染过程中蛋白质功能特征的有力工具,它可以提供比基因组研究更准确的数据。近年来,利用同位素标记相对和绝对定量(isobaric tags for relative and absolute quantitation,iTRAQ)研究病毒诱导的蛋白质变化在植株中已有报道[14-16]。对烟草花叶病毒(tobacco mosaic virus,TMV)侵染的烟草进行iTRAQ 蛋白质组分析,表明烟草中保持D1 蛋白的稳定性和能量的合理利用对抵抗TMV 的侵染至关重要[14]。对烟草叶片进行iTRAQ 蛋白质组学分析,表明RNA 诱导的DNA 甲基化在防御双生病毒卫星DNAgeminivirus-betasatellite 感染中发挥重要作用[15]。利用iTRAQ 技术结合液相二级质谱 ( liquid chromatography-tandem mass spectrometry,LC-MS/MS),在黄瓜叶片中鉴定到38 个参与CGMMV 胁迫应答的差异表达蛋白,其中23 个蛋白质呈现上调表达,15 个蛋白质呈现下调表达[16]。对CGMMV 接种前后的西瓜果实进行iTRAQ 蛋白质组学分析,共鉴定到595 个差异表达蛋白,包括404 个上调蛋白和191 个下调蛋白,其中与叶绿素代谢、丙酮酸代谢、TCA 循环、热休克蛋白、硫氧还蛋白、核糖体蛋白、翻译起始因子和延伸因子相关蛋白的积累水平受到CGMMV 侵染的强烈影响;功能注释表明这些差异表达蛋白主要参与光合作用、碳水化合物代谢、次生代谢物质合成、植物与病原体互作、蛋白质合成与转化等[17]。本研究以iTRAQ 技术为基础,结合LC-MS/MS技术,对CGMMV 侵染前后的西瓜叶片进行蛋白质组学分析,以期揭示参与西瓜叶片响应CGMMV 胁迫的蛋白质。
1 材料与方法
1.1 前处理试验
将易感CGMMV 的西瓜自交系材料JJZ-M(保存于浙江省农业科学院蔬菜研究所西瓜研究室)种植于温度25℃,光照/黑暗(16 h/8 h),湿度60%的光照培养箱中,待两片真叶期进行接种。取人工接种CGMMV 的发病本氏烟草叶片和10 mmol.L-1磷酸盐缓冲液(phosphate buffered saline,PBS,pH 值7.2)按1 ∶5(w/v)的比例混合研磨成糊状病毒汁液。
试验组:取两片真叶期的植株,在叶面散上金刚砂,蘸取制作好的少量病毒汁液轻轻搓揉叶片2 ~3次,用清水冲去多余的金刚砂;对照组(CK,即0 h):植株只接种10 mmol.L-1磷酸盐缓冲液。分别于接种后0 h、48 h 和25 d 取样,3 个取样阶段的叶片症状见浙江省农业科学院蔬菜研究所西瓜组早期的报道[11]。与接种0 h 的植株相比,接种CGMMV 25 d 的叶片症状明显,包括叶片形成绿色凹凸病斑及叶脉萎缩,而接种48 h 的植株无明显病症[11]。每个处理设置2 个生物学重复,用于iTRAQ 测序及分析。
1.2 蛋白质提取、定量、酶解和肽段标记
将接种CGMMV 0 h、48 h 和25 d 的西瓜叶片加入5 倍体积的放射免疫沉淀裂解液,超声破碎处理5 min,于4℃、10 000×g条件下离心10 min;取上层清液,配置标准品溶液,在酶标仪上测定蛋白质的浓度。
每个处理样品中吸取100 μg 蛋白质于EP 管,并用 100 mmol.L-1三乙胺- 碳酸缓冲溶液(triethylammonium bicarbonate,TEAB,pH 值8.5)将体积调整为100 μL;随后加入5 μL 200 mmol.L-1三羧基乙基膦溶液[Tris(2-carboxyethyl) phosphine buffer,TCEP,pH 值8.5],将样品置于55℃条件下孵育1 h;取5 μL 375 mmol.L-1碘乙酰胺溶液(使用前,将9 mg碘乙酸酰胺溶于132 μL 100 mmol.L-1TEAB 溶液中,配制成375 mmol.L-1碘乙酸酰胺)加入样品中,室温下避光孵育30 min;在样品中加入600 μL 预冷丙酮,于-20℃放置不少于4 h;之后,于4℃、8 000×g条件下离心10 min,保留白色沉淀,室温干燥2 ~3 min;取100 μL 50 mmol.L-1TEAB 溶液重溶蛋白质沉淀,添加2.5 μg 胰蛋白酶后在37℃下酶切蛋白过夜。
肽链利用串联质谱标签(tandem mass tag,TMT)试剂标记,标记在室温下进行1 h。标记的肽在分馏前使用TMTsixples 等压质量标记试剂盒(美国赛默飞世尔科技公司)进行操作和干燥。
1.3 肽分离与LC-MS/MS 分析
利用 Pierce High pH Reversed-Phase Peptide Fractionation Kit(美国赛默飞世尔科技公司)分馏已标记的肽,孵育后的样品溶于300 μL 0.1% 三氯乙酸溶液中。在样品中加入300 μL 的5% 乙腈-0.1% 三乙胺去除未反应的TMT 溶液,然后加入300 μL 洗脱液,3 000×g离心2 min,收集片段。真空离心法将每个离心管中的液体蒸发至干,分离的肽在含有0.1%甲酸和2%乙腈的缓冲液中溶解。12 000×g、4℃离心10 min 后,转移到LC-MS 样品瓶中。片段在Easy-nLC 1000(美国赛默飞世尔科技公司)上运行,该nLC 1000与Orbitrap Q ExactiveTMHF 质谱仪耦合(美国赛默飞世尔科技公司)。
1.4 蛋白质鉴定和定量
通过Proteome Discoverer 2.0 将原始数据文件转换为MGF 文件。利用MASCOT v2.3(英国Matrix 科学 公司) 将数据比对到西瓜UniProtKB 数据库(https:/ /www.uniprot.org/)进行蛋白质的鉴定及定量。母离子容差设置为10-5,MS/MS 碎片离子的质量容差设置为0.6 Da,胰蛋白酶消化过程中允许两次缺失裂解。将甲硫氨酸氧化作为可变修饰,将半胱氨酸氨基甲酰化作为固定修饰。选择单一同位素质量,只有在95%置信水平下具有显著得分的多肽是可信的,并用于蛋白质鉴定。
1.5 差异表达蛋白鉴定及KEGG 功能分析
对差异表达蛋白(differentially expressed proteins,DEPs)进行筛选鉴定,将FC≥1.2(或≤1/1.2)和P<0.05 作为筛选标准。FC≥1.2 的蛋白认为是上调,FC≤1/1.2 的蛋白认为是下调。P值由t检验计算得出。对差异表达蛋白进行基于KEGG Pathway 的功能富集分析。
1.6 蛋白质-蛋白质相互作用网络分析
在基于BLAST 算法的搜索数据库(用于检索基因/蛋白质相互作用的搜索工具)中,DEPs 用作原始序列以揭示蛋白质-蛋白质相互作用(protein-protein interaction,PPI)网络。STRING 是收录多个物种预测和试验验证的PPI 的数据库,包括直接的物理互作和间接功能相关。每个相互作用都有一个综合得分,代表蛋白质之间相互作用的可信度,综合得分(0:最低置信度;1:最高置信度)大于0.4 的PPI 做进一步网络分析,并用Cytoscape 将蛋白互作网络图视化。
1.7 RT-qPCR 验证蛋白质在CGMMV 胁迫下的表达
与蛋白质组测序的取样时间一致,分别在接种CGMMV 后0 h、48 h 和25 d 采集西瓜叶片,提取总RNA。用 TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix 试剂盒(北京全式金生物技术有限公司)将总RNA(2 μg)反转录成cDNA。实时荧光定量PCR(quantitative real-time PCR,RT-qPCR)使用TransStart Top Green qPCR Supermix 试剂盒(北京全式金生物技术有限公司),在StepOne Plus Real-Time PCR System(美国ABI 公司)中完成。反应程序:95℃预变性30 s,95℃变性5 s,55℃退火15 s,72℃延伸10 s;40 次循环。内参基因为Tubulin,采用2-ΔΔCT法计算基因相对表达量[18]。设置3 个重复。用于RT-qPCR分析的引物见表1。

表1 RT-qPCR 分析所用的引物Table 1 Primers used for RT-qPCR analysis
2 结果与分析
2.1 基于iTRAQ 分析的蛋白质鉴定
iTRAQ 分析共产生396 个多肽,只有不到4.3%的肽段具有≥20 个氨基酸残基,92.4%的肽段具有7~16个氨基酸残基(图1)。通过比对UniProtKB 数据库,共鉴定获得88 个蛋白质(表2)。通过对每个处理2 个重复间蛋白质表达水平的相关系数(r2)进行分析,发现r2大于0.99,证实了测序结果的可靠性(图2)。

表2 利用iTRAQ 在3 个文库中鉴定到的蛋白质Table 2 Proteins identified using iTRAQ in three libraries

表2(续)

表2(续)

图1 西瓜CGMMV 侵染前后的多肽长度分布Fig.1 Distribution of peptide lengths in watermelon before and post CGMMV infection
2.2 参与西瓜对CGMMV 胁迫应答的DEPs
当蛋白质的FC≥1.2 且P<0.05 时,认为其表达上调;当蛋白质的FC≤1/1.2 且P<0.05 时,认为其表达下调。根据该标准,鉴定到61 个DEPs(表3、图3)。与接种0 h 相比,15 个蛋白质在接种后48 h 呈现差异表达(均为下调表达),37 个蛋白质在接种后25 d 呈现显著差异表达(24 个上调,13 个下调);与接种后48 h 相比,51 个蛋白质在接种后25 d 呈现显著差异表达(45 个上调,6 个下调)。这些DEPs 包括热激蛋白和生物防御素类蛋白、光系统Ⅰ/Ⅱ蛋白、ATP 合成酶、ATP 合酶、NADH 脱氢酶、真核翻译起始因子、核糖体蛋白、乙烯受体、乙烯响应因子、蔗糖合成酶和抗坏血酸过氧化物酶。例如,核糖体蛋白L22(A0A1P8LDU4)、光系统Ⅰ铁硫中心(A0A1P8LDV7)和光系统Ⅱ反应中心蛋白H(A0A1P8LEZ9) 在48 h_0 h、 25 d_0 h 和25 d_48 h 表达下调,而核糖体蛋白S4(D5I3A5)、NADH 脱氢酶亚基9(D5I3C1)、HSP22.8(H6TB36)、真核细胞翻译起始因子6(Q6I663)、蔗糖合酶( Q9SBL8) 和乙烯受体(V9QGW9)在25 d_0 h 和25 d_48 h 表达上调。
2.3 CGMMV 胁迫应答DEPs 的KEGG 功能分析
通过对DEPs 进行KEGG 富集分析鉴定参与CGMMV 胁迫应答的代谢途径。由图4 可知,48 h_0 h的DEPs 主要参与光合作用、核糖体、α-亚麻酸代谢、乙醛酸和二羧酸代谢、氧化磷酸化、光合生物中的碳固定、碳代谢作用、代谢途径和次生代谢产物的生物合成等代谢途径。而25 d_0 h 和25 d_48 h 的DEPs 主要参与植物激素信号转导、类胡萝卜素生物合成、糖酵解/糖异生、嘌呤代谢、嘧啶代谢、RNA 聚合酶、RNA 转运和真核生物核糖体生物合成、光合生物中的碳固定、淀粉和蔗糖代谢、抗坏血酸和醛酸代谢、次生代谢产物的生物合成等代谢途径。

表3 在3 个文库中鉴定的差异表达蛋白Table 3 DEPs identified among three libraries

表3(续)
2.4 西瓜响应CGMMV 胁迫的DEPs 的PPI 分析
在基于BLAST 算法的搜索数据库(用于检索基因/蛋白质相互作用的搜索工具),DEPs 被用作原始序列,并使用综合评分大于0.4 的PPIs 来确定互作网络。48 h_0 h 的13 个DEPs 在CGMMV 胁迫下参与已知的蛋白互作网络,分别是PSAA(A0A1P8LDG4)、NDHI(A0A1P8LDL8)、RPL22(A0A1P8LDU4)、PSAC( A0A1P8LDV7 )、 RPS11 ( A0A1P8LEJ1 )、 rps2(A0A1P8LEM9)、RPS7.2(A0A1P8LEV5)、AtCg00380( A0A1P8LEX7 )、 PSBH ( A0A1P8LEZ9 )、 YCF3( A0A1P8LF51 )、 PSBE ( A0A1P8LF81 )、 RBCL(A5X4H2)和LHCB6(D1MWY9)(图5-A)。
25 d_0 h 的32 个DEPs 在CGMMV 胁迫下参与已知的蛋白互作网络,分别是RPOB(A0A1P8LF53)、RPL22(A0A1P8LDU4)、PSBH(A0A1P8LEZ9)、NAD7( D5I3D0 )、PETD ( A0A1P8LF98 )、 YCF3( A0A1P8LF51 )、 RBCL ( A5X4H2 )、 RPOC2( A0A1P8LDN2 )、 PSAC ( A0A1P8LDV7 )、 ABA1(F8QV17)、PSBE(A0A1P8LF81)及NAD9(D5I3C1)等(图5-B)。

图3 3 个文库中DEPs 的聚类分析Fig.3 Cluster analysis of DEPs identified from three libraries
25 d_48 h 的43 个DEPs 在CGMMV 胁迫下参与已知的蛋白互作网络,分别是RPL2.2(A0A1P8LDK7)、 RPS12A ( A0A1P8LF74)、 RPL22( A0A1P8LDU4 )、 NAD7 ( D5I3D0 )、 RPS11( A0A1P8LEJ1 )、 PSAC ( A0A1P8LDV7 )、 PSBB( A0A1P8LDA4 )、 RPOB ( A0A1P8LF53 )、 rps2(A0A1P8LEM9)、AtCg00380(A0A1P8LEX7)及PSBH(A0A1P8LEZ9)等(图5-C)。
这些 DEPs 中, PSAC (A0A1P8LDV7)、 RPL22(A0A1P8LDU4)和PSBH(A0A1P8LEZ9)共同参与了上述互作网络,表明它们在西瓜CGMMV 胁迫应答中起着关键作用。
2.5 RT-qPCR 验证DEPs 的基因表达
选取SUS4(Q9SBL8)、RPL22(A0A1P8LDU4)、NAD7(D5I3D0)、PSBH(A0A1P8LEZ9)、Defensin-like protein 5 ( V5LFZ0)、 ATPF ( A0A1P8LD68)、 LSP1(C7SG38) 和HSP22.8(H6TB36) 蛋白质进行RTqPCR 验证(图6)。SUS4 ( Q9SBL8 )、RPL22(A0A1P8LDU4)、ATPF( A0A1P8LD68) 和PSBH(A0A1P8LEZ9)在接种后48 h 和25 d 呈逐渐下调表达。HSP22.8(H6TB36)在接种后48 h 和25 d 呈逐渐上调表达。NAD7(D5I3D0)在接种后48 h 的表达下调,而后在接种后25 d 维持与接种后48 h 相似的表达水平。Defensin-like protein 5(V5LFZ0)在接种后48 h 的表达上调,而在接种后25 d 的表达显著下调。LSP1(C7SG38)在接种0 h 和接种后48 h 维持稳定的表达水平,而在接种后25 d 的表达水平显著上调。
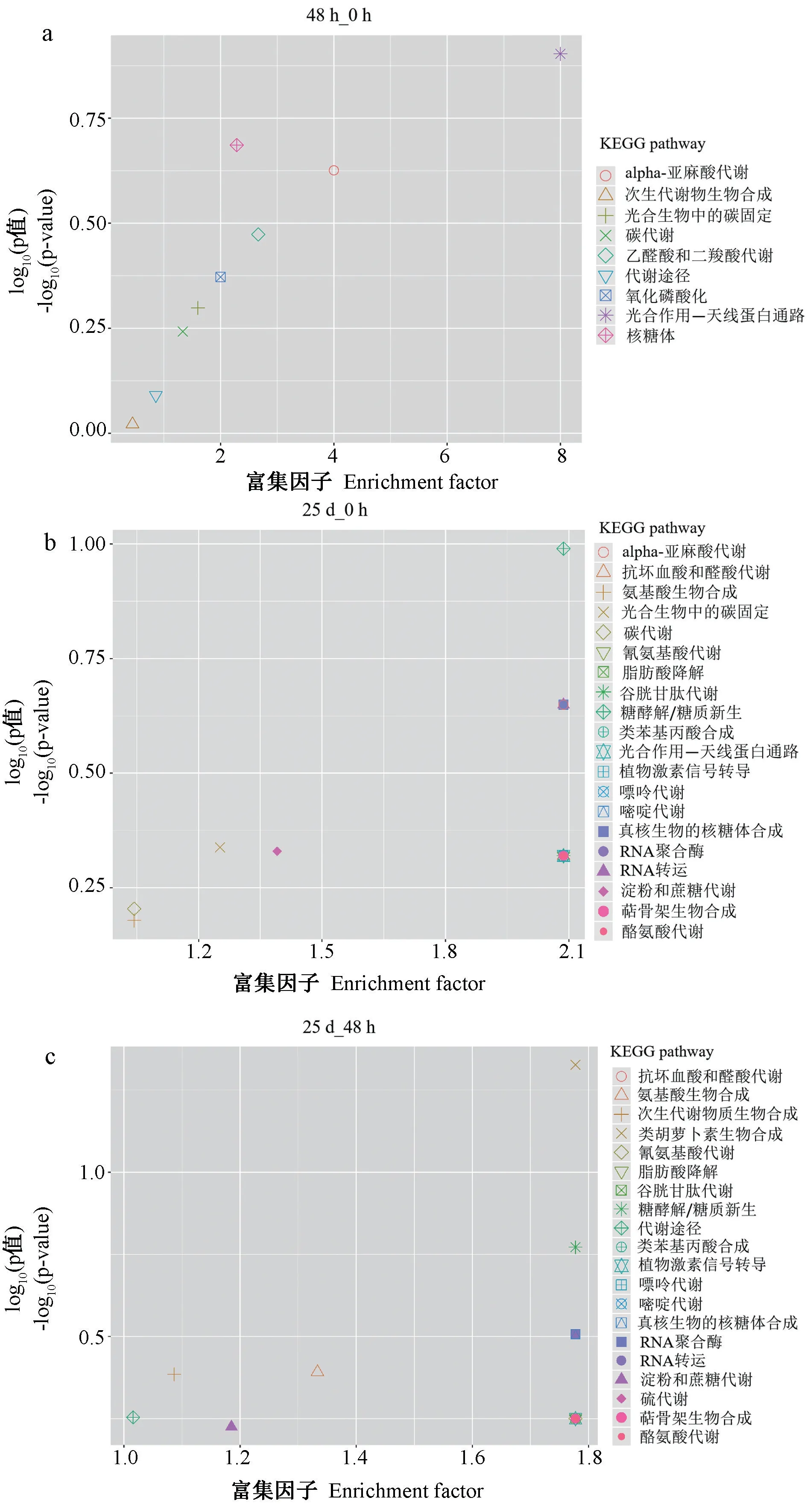
图4 DEPs 的KEGG 功能富集分析Fig.4 KEGG pathway enrichment analysis of DEPs
3 讨论
mRNA、miRNAs 和siRNAs 等诸多因子参与西瓜对CGMMV 的胁迫应答[9-13]。为了在蛋白质水平上鉴定参与CGMMV 胁迫应答的蛋白质,本研究利用iTRAQ 结合LC-MS/MS 技术共获得61 个DEPs。与黄瓜响应CGMMV 胁迫的DEPs 相比,本研究中的部分DEPs,如细胞色素b6、核糖体蛋白、肌动蛋白及NADP(H)氧化还原酶等,参与西瓜对CGMMV 的胁迫应答。

图5 西瓜响应CGMMV 的DEPs 的蛋白质-蛋白质相互作用网络Fig.5 Protein-protein interaction analysis of DEPs in response to CGMMV stress in watermelon
3.1 CGMMV 胁迫影响西瓜诸多生理生化过程
受病毒感染的植物通常表现为光合速率降低,这表明光合作用是宿主抗病毒防御应答中被抑制的主要生命活动[19]。本研究中,7 个光系统Ⅰ/Ⅱ蛋白( A0A1P8LDA4、 A0A1P8LDG4、 A0A1P8LDM9、A0A1P8LDV7、 A0A1P8LEY8、 A0A1P8LEZ9 和A0A1P8LF51)被鉴定为DEPs,这与之前的报道一致。例如,转基因番茄中与光系统Ⅰ/Ⅱ相关的蛋白质在CMV 的胁迫应答中呈现差异表达[20]。其中,光系统Ⅱ反应中心蛋白H(A0A1P8LEZ9)是光系统Ⅱ反应中心核心的重要组成部分,可将光能转换为电化学能[21],其在CGMMV 感染后48 h 和25 d 呈下调表达。此外,光系统Ⅱ47 kDa 蛋白和光系统ⅡD1 蛋白在接种CGMMV 后25 d 呈上调表达,表明维持正常光合作用对西瓜抵御CGMMV 的侵染至关重要。
能量代谢受植物-病毒相互作用的影响。本研究中,3 个ATP 合酶 亚基(A0A1P8LD68、D5I3A7 和D5I3B4) 和2 个NADH 脱氢酶亚基(D5I3C1 和D5I3D0)参与西瓜对CGMMV 的胁迫应答,其中ATP酶(D5I3A7 和D5I3B4)和NADH 脱氢酶(D5I3C1 和D5I3D0)的表达上调。拟南芥ATP 酶CRT1 可与多种抗性蛋白相互作用,调节拟南芥对芜菁皱缩病毒的抗病性[22]。水稻OsAAA-APTase1 在SA 介导的对稻瘟病菌(Magnapor the oryzae)的防御反应中发挥重要作用,过表达OsAAA-ATPase1 可显著提高致病相关基因的表达和对M. oryzae的抗性;反之,RNAi 沉默该基因抑制水稻对M. oryzae的抗性[23]。
核糖体蛋白质参与rRNA 结构的产生及蛋白质合成机制的运作,同时也可保护植物免受病原菌的侵害[24-25]。核糖体蛋白编码基因RPL12 和RPL19 在本氏烟草中被沉默后,非寄主细菌诱导的过敏反应延迟;拟南芥AtRPL12 和AtRPL19 突变体的非寄主抗性也降低,说明RPL12 和RPL19 通过同一途径获得非寄主抗性[24]。棉花中GaRPL18的表达受到黄萎病菌(Verticillium dahliae)侵染的诱导,GaRPL18 的表达模式与不同品种的抗病性水平一致;GaRPL18 基因沉默使抗性材料中免疫相关因子的丰度降低,抗性材料变为易感材料;GaRPL18 过表达拟南芥对黄萎病的抗性明显提高[25]。本研究中,9个核糖体蛋白( A0A1P8LDK7、A0A1P8LDU4、A0A1P8LEJ1、A0A1P8LEJ2、A0A1P8LEM9、A0A1P8LEV3、A0A1P8LEV5、A0A1P8LEX7 和D5I3A5) 在CGMMV侵染后呈现差异表达,表明这些核糖体蛋白在CGMMV 胁迫应答中发挥重要作用。
乙烯作为重要的信号可调控植物对一系列生物和非生物胁迫的应答。研究报道烟草中EREBP/AP2 过表达可诱导多个致病相关基因的表达,提高烟草对病原体及盐胁迫的耐受性[26]。本研究发现了2 种参与乙烯信号转导的差异表达蛋白,分别是乙烯受体(V9QGW9)和乙烯响应因子(X2KY76)。其中,乙烯受体在25 d_0 h 和25 d_48 h 呈上调表达,乙烯响应因子在25 d_48 h 呈上调表达,表明诱导的乙烯信号蛋白与CGMMV 的防御应答有关。
3.2 CGMMV 胁迫下的重要蛋白质及调控机理
植物病毒可以在感染周期通过宿主植物的翻译因子来翻译其自身的病毒RNA,翻译起始因子在植物-病毒相互作用中赋予植物抵抗病毒感染的能力[27-29]。与0 h 和48 h 相比,本研究中的2 个真核翻译起始因子:eIF (iso) 4E (C7SG38) 和eIF6A (Q6I663), 在CGMMV 侵染后25 d 呈现显著上调表达,参与对CGMMV 的胁迫应答。植物eIF4E 家族成员是众所周知的宿主因子,在多种病毒感染中起关键作用[27-29]。植物eIF4E 的主要作用是通过结合5′cap [m7G(5′)ppp(5′)N]和其他翻译因子,进而控制蛋白质在真核细胞中的表达[30]。大白菜eIF(iso)4E 基因(retr02)的剪接变异体在第1 个外显子处产生一个终止密码子,产生一个截短、无功能的蛋白质,芜菁花叶病毒(turnip mosaic virus,TuMV)可以在2 个位点上使用eIF(iso)4E 的拷贝,使大白菜产生持久的TuMV 抗性[31]。TuMV 感染植物时,需要病毒末端结合蛋白(viral protein genome-linked,VPg)与宿主eIF4E 或eIF(iso)4E 相互作用才能启动翻译[32]。拟南芥eIF(iso)4E 突变导致对TEV、TuMV 产生广谱抗性[33]。此外,TuMV VPg 仅与拟南芥eIF(iso)4E 基因(AT5G35620)相互作用,而不与拟南芥的其他eIF4E 发生互作[34]。eIF6 是60S 核糖体核仁生物合成所必需的蛋白[35],但目前鲜有报道表明其参与植物对病毒的胁迫应答。本研究中,eIF6A 与RAN3、LSP1、RPL23.2、RPS15A 等蛋白互作,参与CGMMV 的胁迫应答。
热激蛋白(heat shock proteins,HSPs)被认为具有分子伴侣的功能,在植物中普遍存在,当机体受到不同胁迫时,HSPs 可以保护其靶蛋白不变性,不发生错误折叠和聚集[36-37]。sHSPs 作为HSPs 的重要成员,其分子量为15 ~42 kDa,在应答植物生物胁迫中发挥重要作用[38]。本研究中sHSPs 成员HSP22.8(H6TB36)的表达在CGMMV 接种后25 d 显著上调。水稻HSP20 的表达受水稻条纹病毒(RSV)感染的影响,此外,发现HSP20 与RSV 蛋白(RdRp) 和运动蛋白(NSVc4)相互作用,在RSV 复制和运动中发挥重要作用[39-40]。CGMMV 侵染西瓜后,sHSP 的表达上调,进一步通过酵母双杂、GSP pull-down 及免疫共沉淀分析发现,CGMMV 解旋酶(helicase HEL)可与西瓜sHSP互作,说明sHSP 可能调节宿主防御机制以应对CGMMV 的侵染[41]。
3.3 CGMMV 胁迫下的蛋白质组与转录组比较分析
浙江省农业科学院蔬菜研究所西瓜组前期对CGMMV 侵染前后(0 h 及24 h)的西瓜叶片进行转录组测序,共获得1 641 个差异表达基因(DEGs),包括886 个上调和755 个下调DEGs[10]。本研究共鉴定到61 个DEPs。与DEPs 相似,KEGG 功能分析显示这些DEGs 同样参与光合作用、植物激素信号转导、次生物质代谢等KEGG 代谢途径[10]。通过DEGs 与DEPs 的比较分析,发现叶绿素a/b 结合蛋白、NADH 脱氢酶、核糖体蛋白、光系统Ⅰ/Ⅱ相关蛋白、β-葡萄糖苷酶、蔗糖合成酶、HSP22 等在蛋白水平和基因水平上均呈现差异表达。如叶绿素a/b 结合蛋白(D1MWY9)在CGMMV 侵染后48 h 和25 d 均呈现显著下调表达;叶绿素 a/b 结合蛋白基因(Cla004746、 Cla018117、Cla022573、 Cla019105、 Cla017325、Cla022963、Cla011748、 Cla012368、 Cla019595、 Cla013826 及Cla011145)在CGMMV 侵染后24 h 均呈现显著下调表达。与蛋白表达水平一致,核糖体蛋白、NADH 脱氢酶及光系统Ⅰ/Ⅱ相关蛋白的编码基因在CGMMV 侵染后24 h 呈现显著下调表达。β - 葡萄糖苷酶(G9BSA4)及蔗糖合成酶(Q9SBL8)在CGMMV 侵染后25 d 呈现显著上调表达,而β-葡萄糖苷酶基因(Cla004804) 及蔗糖合成酶基因(Cla017392、Cla011131 及Cla009124)在CGMMV 侵染后24 h 即呈现显著上调表达。上述结果表明,参与西瓜光合作用的蛋白和基因在CGMMV 胁迫早期即表现出响应;而参与次生物质代谢的蛋白对CGMMV 胁迫应答的响应较基因要延迟一些。
4 结论
本研究通过iTRAQ 技术鉴定到61 个参与CGMMV 胁迫应答的蛋白质,这些DEPs 参与光合作用、植物激素信号转导、次生物质代谢等KEGG 代谢途径。此外,本研究还明确了这些蛋白质之间的相互作用和调控网络。本研究结果在蛋白质水平上明确了西瓜响应CGMMV 的胁迫应答,为后续提高西瓜对CGMMV 的抗性及阐明其分子机理奠定了基础。
猜你喜欢
内蒙古民族大学学报(自然科学版)(2022年2期)2022-11-22
生物化学与生物物理进展(2022年9期)2022-09-22
科学导报(2022年29期)2022-05-26
当代水产(2022年1期)2022-04-26
中国瓜菜(2019年8期)2019-09-19
小天使·一年级语数英综合(2019年3期)2019-04-19
小天使·一年级语数英综合(2017年6期)2017-06-07
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
文理导航·趣味课堂(2016年6期)2016-09-09
