
能源甜菜转录因子BvMYB44基因响应镉胁迫的表达特性及生物信息学分析
2021-02-15 09:26:12周婉婷李思琪王雪倩王录红李王胜李佳佳刘大丽
中国农学通报 2021年35期
汪 曼,周婉婷,周 翔,李思琪,王雪倩,王录红,李王胜,李佳佳,高 卓,3,刘大丽
(1国家甜菜种质中期库,黑龙江大学,哈尔滨 150080;2省高校甜菜遗传育种重点实验室/黑龙江大学现代农业与生态环境学院,黑龙江大学,哈尔滨 150080;3黑龙江省普通高等学校分子生物学重点实验室/黑龙江大学生命科学学院,哈尔滨 150080)
0 引言
随着工业化进程的加速,中国已成为全球土壤污染最严重的国家之一。工业排放、污水灌溉、化肥农药的不合理施用及大气沉降等原因已造成不同程度的土壤重金属污染[1]。中国每年因重金属污染土壤而导致的粮食减产达1000万t以上[2],其中重金属镉(Cd)污染居所有污染物的首位[3]。因此围绕Cd元素展开土壤修复探讨比较具有代表性,生物技术治理作为一种土壤修复领域中的新型治理措施,正在被广泛推广并运用,而这其中目前最具实践意义的是植物修复[4]。
能源甜菜(Beta vulgarisL.)因其非生物胁迫适应能力强,生物学产量高,乙醇转化率高,是新兴的可再生能源作物,在重金属污染土壤的修复中表现出了极大的潜力,是植物修复的优良目标物种[5]。而植物修复这一系列复杂的生理生化过程中就涉及到了转录因子的调控[6]。转录因子是真核生物体内一大类转录调控因子,能够通过特异性结合相关顺式作用元件来激活或抑制下游相关基因的表达[7]。植物基因组比其他真核生物基因组包含更多的转录因子,其中bZIP、WRKY、AP2/ERF和MYB转录因子与抗逆反应密切相关,在植物基因的表达调控系统中一直备受关注[8]。
MYB转录因子家族作为植物中最大的转录因子家族之一,已被证明在植物应对非生物胁迫反应中有重要调控作用[9]。MYB转录因子由高度保守的MYB DNA结合结构域定义,一般位于N末端,MYB结构域中分别具有1、2、3和4个肽重复序列,根据其相邻不完全重复序列的数量,MYB转录因子被分为4个亚家族 ,分别为 MYB-related(1R-MYB)、R2R3-MYB(2RMYB)、R1R2R3-MYB(3R-MYB)和4R-MYB转录因子亚家族[10-11]。MYB转录因子最初于1982年在禽成髓细胞瘤病毒(AMV)中被发现,并被命名为“v-MYB”[12]。5年后,第一个被成功克隆的MYB基因为玉米(Zea maysL.)中色素合成相关基因ZmMYBC1[13]。迄今为止已在多种植物中发现MYB转录因子家族,其中在模式植物拟南芥中已发现有339个MYB家族的成员,水稻(Oryza sativaL.)中已发现有230个[14-15]。
目前关于MYB转录因子参与非生物胁迫反应的研究大多集中在干旱胁迫、盐胁迫、温度胁迫以及磷胁迫[16-18],而关于重金属胁迫的报道相对较少。甜菜中共筛选出79个MYB转录因子,其中42个在干旱胁迫下差异表达;70个为R2R3-MYB基因,其中BvMYB12为第一个具有实验证明功能的甜菜R2R3-MYB,编码黄酮醇调节因子[19-20]。总的来说甜菜中MYB基因的相关功能研究鲜有报道,且未见其参与重金属胁迫的相关研究。
本研究中的能源甜菜转录因子BvMYB44基因是在本课题组前期镉胁迫转录组表达谱研究中得到的差异表达基因,且表达极为显著,推测其在重金属胁迫中发挥着重要作用。因此本研究对能源甜菜转录因子BvMYB44进行了一系列生物信息学分析,并通过实时荧光定量PCR(qRT-PCR)实验来探究转录因子BvMYB44基因响应重金属镉胁迫的表达特性,为进一步解析转录因子BvMYB44基因在能源甜菜重金属胁迫中的调控作用奠定基础,同时也为抗重金属能源甜菜的种质创新以及拓展植物修复提供候选基因和理论基础。
1 材料与方法
1.1 能源甜菜BvMYB44基因的生物信息学分析
利用NCBI数据库的ORF Finder在线分析BvMYB44基因的开放阅读框和编码的氨基酸。利用GSDS2.0分析该基因外显子和内含子的结构。利用ProtParam tool分析该蛋白的分子量、等电点相关理化性质。通过ProtScale来预测该蛋白的亲/疏水性。通过SignalP-5.0 Server预测该蛋白信号肽的位置。利用TMpred分析该蛋白的跨膜结构域。利用SOPMA在线预测分析BvMYB44蛋白的二级结构。利用SWISS-MODEL在线预测该蛋白的三级结构。利用SMART在线工具对该蛋白的结构域进行分析。利用MEME软件对该蛋白的保守位点进行分析。利用NCBI数据库的BLASTP获得与BvMYB44氨基酸序列同源性较高的不同物种,用DNAMAN8软件进行多序列比对,利用MEGA7软件的最大似然法构建进化树,Bootstrap值设置为1000次。利用NetPhos 3.1 Server预测该蛋白的磷酸化位点。利用BaCelLo对BvMYB44蛋白的亚细胞定位进行预测。利用PlantCARE在线工具分析该基因启动子区的顺式作用元件。
1.2 时间与地点
实验于2020年10月—2021年4月,在黑龙江大学省高校甜菜遗传育种重点实验室实施并完成。
1.3 材料与处理
植物材料能源甜菜种子‘780016B/12优’由国家甜菜种质资源中期库(国家甜菜种质资源平台)提供使用。将能源甜菜种子‘780016B/12优’在培养室内,用14 h光照25℃,10 h黑暗18℃条件培养,四周后用CdCl2进行镉胁迫处理。镉胁迫浓度为0.5 mmol/L,分别于0、6、12、24、48 h收集镉胁迫下的叶和根组织,液氮冷冻后置于-80℃冰箱备用。
1.4 总RNA的提取
将上述镉胁迫处理的甜菜叶和根组织用EasyPure Plant RNA Kit(TransGen)试剂盒进行总 RNA的提取,用1.0%的琼脂糖凝胶电泳检测所提取RNA的质量,并通过超微量分光光度计测定RNA的浓度和纯度。
1.5 反转录cDNA
根据TransStart One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen)反转录试剂盒说明,取适量总RNA进行cDNA第一链的合成。将42℃孵育30 min的合成产物cDNA置于-20℃冰箱备用。
1.6 能源甜菜BvMYB44响应镉胁迫的表达分析
根据BvMYB44序列,利用NCBI的Primer-BLAST在线工具设计特异性的荧光定量PCR引物(F:5’-ACC GGAAAGAAG TGGACC GGATAAA-3’,R:5’-AAT CAA AGA CCA GTT TCT TGG GCC G-3’)。以BvGAPDH为内参基因(F:5’-GCT TTG AAC GAC CAC TTC GC-3’,R:5’-ACG CCG AGA GCA ACT TGA AC-3’)。 以 SYBR Green 为 标 记 物 ,采 用SuperReal PreMix Plus(TIANGEN)试剂盒进行 qRTPCR表达分析。实时荧光定量PCR反应体系为20 μL,包含 SuperReal PreMix Plus(2×)10 μL、ROX Reference Dye(50×)0.4 μL、正反向引物(10 μmol/L)各 0.6 μL、cDNA模板1 μL、RNase-free ddH2O 7.4 μL。扩增两步法反应程序为95℃预变性15 min;95℃变性10 s,60℃退火/延伸32 s,40个循环,实验3次重复。利用SPSS 19.0软件统计荧光定量PCR试验数据进行差异显著性分析,采用 2-ΔΔCT法计算基因相对表达量,用Microsoft Excel软件绘制图表。
2 结果与分析
2.1 BvMYB44基因结构特征分析
ORF Finder分析结果显示,BvMYB44基因cDNA全长为942 bp,共编码313个氨基酸。为了解BvMYB44基因的结构特征,用GSDS2.0确定了BvMYB44基因外显子和内含子的数量和位置。结果显示(图1),BvMYB44外显子数量为1,且不含有内含子,符合先前报道的MYB结构域编码序列中一般含有0~2个内含子的特征。

图1 BvMYB44基因结构特征分析
2.2 BvMYB44蛋白的理化性质分析
ProtParamtool在线软件分析BvMYB44蛋白分子式为C1450H2295N435O467S20;BvMYB44分子量为33935.01 Da;理论等电点(Theoretical Pi)为7.57,数值大于7,推测其为碱性蛋白;不稳定系数为51.28,数值大于40,推测其为不稳定蛋白;脂肪指数为62.68;平均亲/疏水系数为-0.576,推测其为亲水蛋白。
2.3 BvMYB44蛋白亲/疏水性、信号肽的预测以及跨膜结构域分析
用ProtScale分析亲/疏水性显示(图2),BvMYB44蛋白氨基酸位点亲水性在第167位最强,数值为-3.289;在第231位疏水性最强,数值为1.400,且整体亲水性位点多于疏水性位点,因此亲/疏水性预测结果表明该蛋白为亲水蛋白,同时这与ProtParam tool分析结果一致。同时,预测分析表明BvMYB44蛋白无信号肽。且该蛋白不存在跨膜区域。
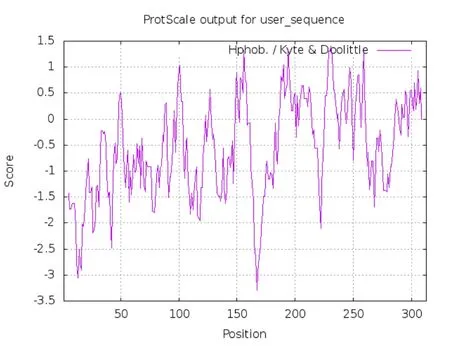
图2 BvMYB44蛋白亲/疏水性预测
2.4 BvMYB44蛋白二级和三级结构预测分析
SOPMA预测的二级结构结果显示,BvMYB44蛋白的组成包含27.16%的α-螺旋,4.79%的延伸链,3.51%的β-转角和64.54%的无规卷曲,因此α-螺旋和无规卷曲为该蛋白二级结构主要元件(图3)。
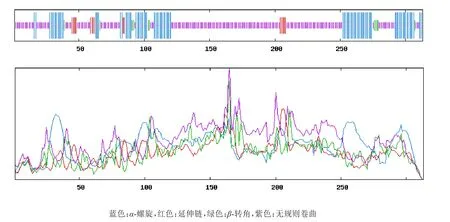
图3 BvMYB44蛋白二级结构预测
如图4所示,使用SWISS-MOLD数据库预测BvMYB44蛋白的三级结构,并进行同源性3D建模,根据评分选择最佳模型,序列同源性为56.73%,数值大于40%,模型可信度高。3D模型显示该蛋白主要由α-螺旋和无规卷曲构成,与SOPMA预测的二级结构结果一致,且具有螺旋-转角-螺旋(HTH)结构,符合MYB结构特征。
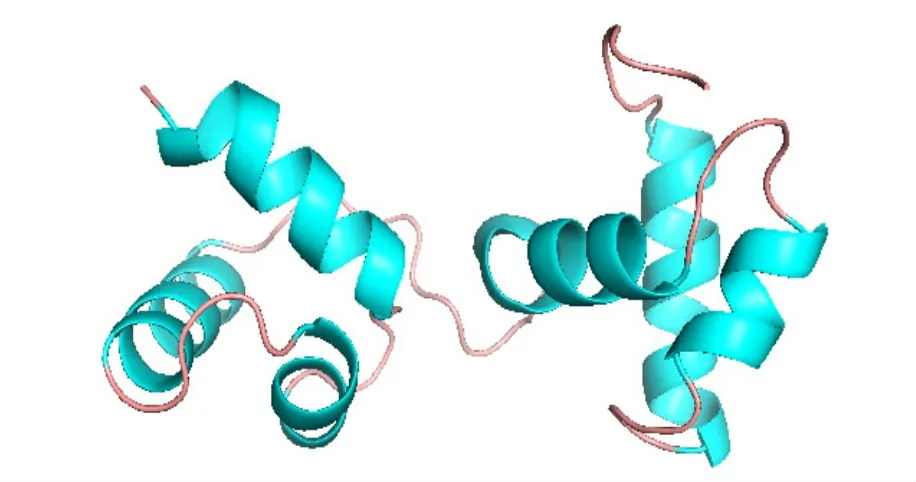
图4 BvMYB44蛋白三级结构预测
2.5 BvMYB44蛋白保守结构域分析
SMART分析BvMYB44蛋白结构域,结果显示其包含2个重复的MYB的DNA保守结构域SANT,这2个SANT结构域分别位于第21~70位和第73~121位氨基酸处,符合典型的MYB家族基因结构域特点,推测其属于R2R3-MYB转录因子亚家族成员(图5)。
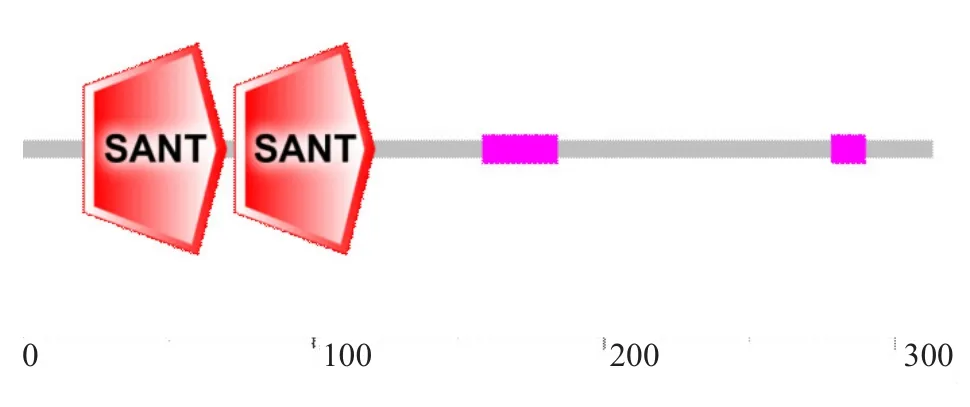
图5 BvMYB44蛋白结构域预测
2.6 BvMYB44蛋白保守位点分析
为进一步了解BvMYB44蛋白保守结构域的具体特征,通过MEME分析其保守位点。字母越高即代表该位点上氨基酸残基出现的概率越大,保守程度也就越高。如图6所示,高度保守的色氨酸(W)残基作为植物MYB蛋白的标志,在R2、R3基序中有一定规律的间隔分布形成疏水中心。除色氨酸残基外图中还包含其他保守的氨基酸残基,它们极有可能共同维持了R2R3-MYB转录因子的HTH结构,在序列特异性的DNA结合中起着关键作用,以保证蛋白能够正确折叠从而行使功能。

图6 BvMYB44蛋白保守位点分析
2.7 BvMYB44蛋白的多序列比对及系统发育分析
用NCBI数据库的BLASTP获得与能源甜菜BvMYB44氨基酸序列同源性较高的其他不同物种,藜麦(Chenopodium quinoa)、菠菜(Spinacia oleracea)、葡萄(Vitis riparia)、相思子(Abrus precatorius)、野生大豆(Glycine soja)、蒺藜苜蓿(Medicago truncatula)、川桑(Morus notabilis)、栓 皮 栎 (Quercus suber)、蔓 花 生(Arachis duranensis)、豇豆(Vigna unguiculata),将以上物种序列和能源甜菜BvMYB44一起用DNAMAN8进行多序列比对,并用MEGA7软件的最大似然法构建系统发育树。
比对结果表明(图7),BvMYB44蛋白的MYB结构域位于N末端,序列保持高度一致,且含有R2、R3不完全重复序列,不完全重复序列间每隔约18个氨基酸残基由1个色氨酸残基间隔开来。而C端序列相似性较低,这有利于该区域转录调控功能的行使。进化结果显示(图8),系统发育树整体分支绝大多数数值大于70%,表明构建的该进化树可信,其中与能源甜菜BvMYB44进化关系最近的是藜麦CqMYB44-like和菠菜SoMYB44-like且同源性高。

图7 BvMYB44与其他MYB类同源蛋白多序列比对
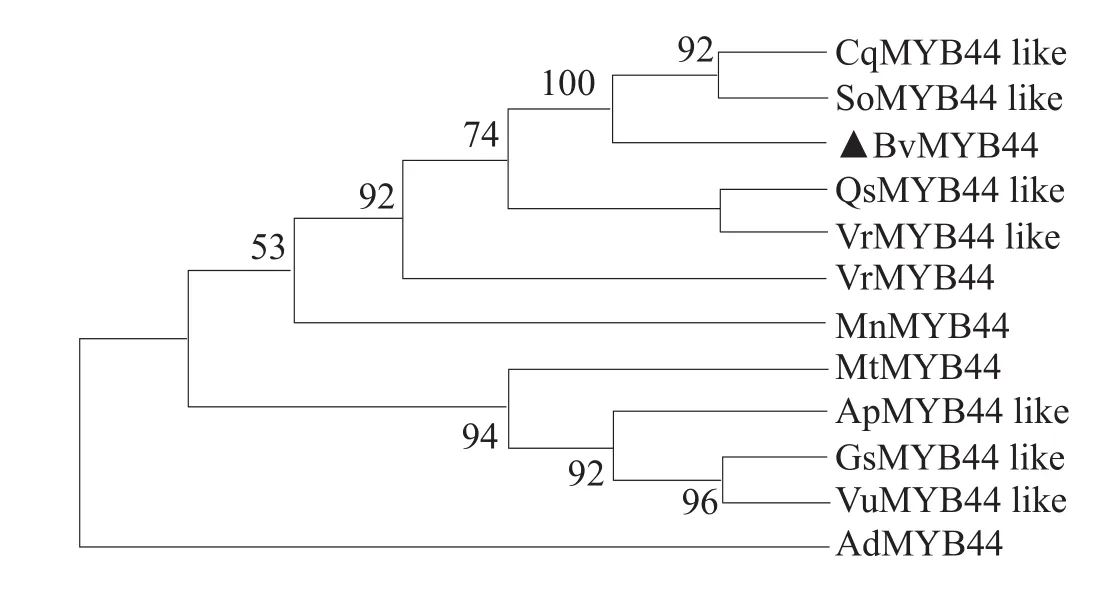
图8 BvMYB44和其他植物MYB蛋白系统进化树构建
2.8 BvMYB44蛋白的磷酸化位点及亚细胞定位预测
NetPhos 3.1 Server磷酸化位点预测BvMYB44结果表明,该蛋白共有39个磷酸化位点,32个丝氨酸磷酸化位点,6个苏氨酸磷酸化位点,1个酪氨酸磷酸化位点。
为研究BvMYB44转录因子的功能机制,利用BaCelLo预测其亚细胞定位,定位结果表明该蛋白定位于细胞核中,这有利于BvMYB44在细胞核内行使其转录调控功能。
2.9 BvMYB44基因启动子区顺式作用元件分析
为探究BvMYB44基因的调控机制,通过NCBI获取该基因起始密码子上游2000 bp的启动子区域,通过PlantCARE来预测其顺式作用元件。预测结果显示(表1)该区域中含有53个CAAT-box和51个TATA-box启动子区基本组成元件,说明该区域是典型的真核生物核心启动子区,且该启动子的转录活性较高。
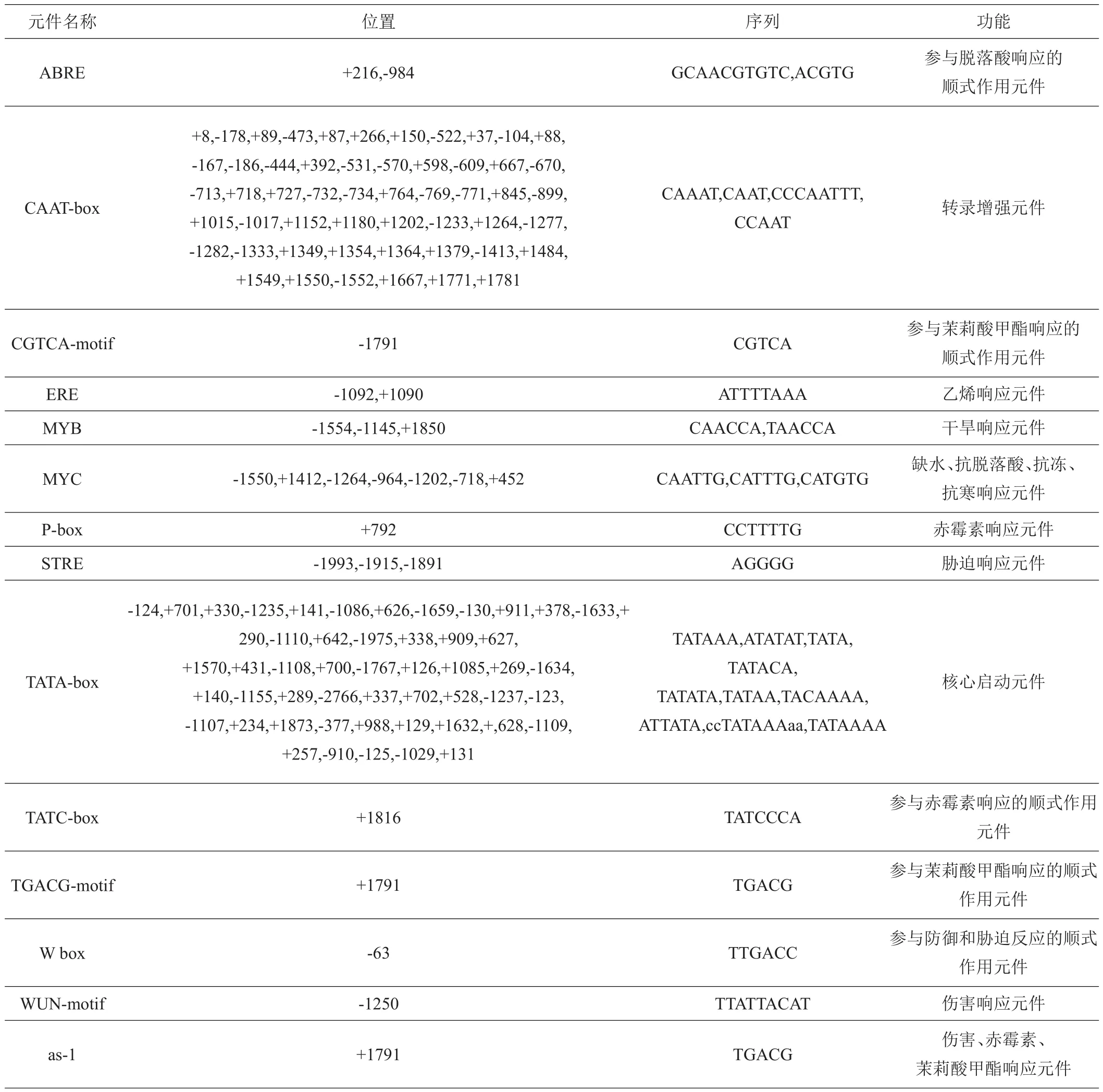
表1 BvMYB44基因启动子区顺式作用元件分析
在BvMYB44启动子序列中,发现了许多应激诱导元件,包括ABRE(脱落酸响应元件)、CGTCA-motif(茉莉酸甲酯响应元件)、ERE(乙烯响应元件)、MYB(干旱响应元件)、MYC(缺水、抗脱落酸、抗冻、抗寒响应元件)、P-box(赤霉素响应元件)、STRE(胁迫响应元件)、TATC-box(赤霉素响应元件)、TGACG-motif(茉莉酸甲酯响应元件)、W box(防御和胁迫响应元件)、WUN-motif(伤害响应元件)、as-1(伤害、赤霉素、茉莉酸甲酯响应元件)。这些结果表明,BvMYB44的转录可能是由激素胁迫和非生物胁迫诱导的。
2.10 BvMYB44基因响应镉胁迫的表达分析
为进一步探究BvMYB44基因应对非生物镉胁迫的响应,对能源甜菜幼苗用0.5 mmol/L浓度的CdCl2进行镉胁迫处理,选取0、6、12、24、48 h的叶和根材料进行qRT-PCR检测。荧光定量PCR结果表明,与未进行镉胁迫处理的对照相比,在镉胁迫处理下,叶和根中BvMYB44基因的表达量均上调,随着镉胁迫处理时间的增加,BvMYB44基因的相对表达量均呈现先上调后下调的趋势(图9)。
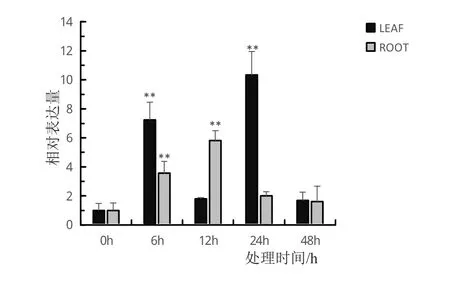
图9 BvMYB44基因在镉胁迫处理下叶和根的相对表达量
在能源甜菜叶组织中,当镉胁迫处理时间为6 h和24 h时,BvMYB44基因的相对表达量上升极为显著,在24 h时达到最大相对表达量10.34,处理时间为12 h和48 h时相对表达量虽相比对照上升但差异不显著。在能源甜菜根组织中,当镉胁迫处理时间为6 h和12 h时,BvMYB44基因的相对表达量上升极为显著,在12 h时达到最大相对表达量5.81,处理时间为24 h和48 h时相对表达量上升差异不显著。从整体来看,除镉胁迫处理时间为12 h时,BvMYB44基因在根组织中的相对表达量大于叶组织外,在镉胁迫处理时间为6 h、24 h和48 h时,BvMYB44基因在叶组织中的相对表达量均大于根组织。该实验结果说明BvMYB44基因的表达量受镉胁迫的响应。
3 讨论与结论
从《2015研究前沿》报告中可以看出,近年来机制性研究正在逐步深入,从基本的功能基因的研究向调控因子上过渡[21]。本研究转录因子BvMYB44基因对镉胁迫的响应,首次证实了调控因子在能源甜菜应对重金属逆境胁迫的重要作用,为能源甜菜在转录调控层面的功能研究提供了理论基础,也为抗重金属能源甜菜作为植物修复优良目标物种的种质创新提供了基因资源。
在被子植物中,R2R3-MYB家族基因作为MYB类转录因子最大的亚家族,该亚家族的大量扩展被认为是因为植物为抵御环境变化而进化复杂性的增加有关[20]。因此R2R3-MYB亚家族可能参与了植物的特异性调节过程,涉及到生长发育、初次生代谢、应答生物和非生物胁迫反应等[10,22],如小麦、玉米、水稻、拟南芥中大量的R2R3-MYB成员直接或间接参与了非生物胁迫响应途径[15,23]。AtMYB44通过调节ABA介导的气孔关闭以响应非生物胁迫,NtMYB44b也参与响应多种激素诱导和干旱胁迫[24]。本研究显示BvMYB44也属于R2R3-MYB亚家族成员,其应对非生物逆境镉胁迫的响应情况符合该家族典型的功能特性。
BvMYB44蛋白的二级和三级结构预测表明,该蛋白富含α-螺旋和无规卷曲结构,含有MYB蛋白的标志结构HTH结构[20,24],保守位点分析也推测出高度保守的色氨酸残基和其他氨基酸残基共同维持了该结构的形成,有利于BvMYB44蛋白功能的行使。磷酸化与蛋白质活性变化密切相关,BvMYB44转录因子共含有39个磷酸化位点,表示该蛋白的调控方式可能由多种翻译后修饰表达的蛋白参与作用,使BvMYB44转录因子参与调控响应多种环境胁迫。多序列比对表明多种不同物种中的MYB转录因子具有较高保守性,与BvMYB44进化关系最近的是藜麦和菠菜且相似度高,SoMYBs有R2R3-MYB成员被分析可能参与非生物胁迫[25],据此推测BvMYB44在进化过程中功能相对保守。
亚细胞定位预测BvMYB44转录因子定位于细胞核中,大多数研究显示转录因子定位在细胞核,或定位在细胞核和细胞质中[18,26],该预测符合转录因子的典型特征,有助于其转录调控功能的行使,后续可通过实验来进一步验证其行使转录调控功能的具体定位。
启动子区顺式作用元件的预测为基因的进一步功能分析提供了基础[27],本研究在BvMYB44基因启动子区发现了脱落酸、茉莉酸甲酯、乙烯、赤霉素多种激素相关元件和MYB、MYC、P-box、STRE、W box、WUN-motif、as-1应激诱导元件。以上顺式作用元件被证明与胁迫反应密切相关,除防御和胁迫响应元件外,赤霉素、脱落酸、茉莉酸甲酯也被证明在非生物胁迫中发挥作用[28-29]。这表明BvMYB44转录因子可能通过调节激素和应激诱导来转导信号通路参与非生物胁迫,后续可通过进一步研究BvMYB44特定结合基序探究其结合活性和调控机制。
前人研究表明,MYB转录因子能够被镉胁迫显著诱导。SbMYB15在重金属镉和镍胁迫处理下上调表达,其异源过表达显著增强了转基因烟草的抗重金属胁迫的能力[30]。镉胁迫反应中,BnMYB2的转录水平显著上调,在拟南芥中过表达该基因增强了转基因拟南芥的镉的耐受性和积累[31]。本研究检测了BvMYB44基因在响应镉胁迫下的相对表达量,该表达模式显示了转录因子BvMYB44基因在应激反应中的积极作用,同时也揭示了BvMYB44可能在植物应对环境胁迫的防御作用中发挥着重要作用,但其所介导的一系列调控机制还有待后续的进一步研究。
总之,MYB转录因子虽已在多种植物中得以广泛研究,已被证明响应非生物胁迫反应,如干旱胁迫、盐胁迫和高温胁迫为主要研究对象,但对重金属胁迫相关的功能探究相对较少,且目前其在能源甜菜中的功能研究鲜有报道。本研究中BvMYB44基因全长942 bp,编码313个氨基酸,属R2R3-MYB亚家族,定位于细胞核,启动子区预测到多个应激诱导元件,qRT-PCR揭示其响应镉胁迫逆境,为镉胁迫的积极响应提供了一些理论基础,可作为能源甜菜镉逆境候选基因进行深入研究。
猜你喜欢
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:04
小哥白尼(趣味科学)(2021年12期)2021-03-16 05:40:44
广州大学学报(自然科学版)(2019年1期)2019-05-07 01:33:26
中国糖料(2016年1期)2016-12-01 06:49:04
现代工业经济和信息化(2016年2期)2016-05-17 05:34:16
天津科技大学学报(2016年1期)2016-02-28 16:59:45
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10 08:41:53
电子工业专用设备(2015年4期)2015-05-26 09:10:40
汽车维修与保养(2015年8期)2015-04-17 03:33:01
现代检验医学杂志(2015年2期)2015-02-06 02:01:01
