
江西省番茄绵疫病菌对嘧菌酯的敏感性检测及抗性风险分析
2021-02-05 09:38何烈干李湘民黄瑞荣马辉刚
农药学学报 2021年1期
何烈干, 邹 芬, 李湘民, 黄瑞荣, 马辉刚
(江西省农业科学院 植物保护研究所,南昌 330200)
番茄绵疫病,又称褐色腐败病、番茄掉蛋,是由寄生疫霉Phytophthora parasitica Dast、辣椒疫霉Phytophthora capsici Leonian 或茄疫霉Phytophthora melongenae Sawada 引起的一种卵菌病害[1-2]。该病在世界各地均有分布,一般田块可导致减产30%~40%,重病田可达60%以上,甚至绝收,经济损失严重[3-5]。番茄为江西主栽蔬菜之一,近年来种植面积大幅增加,但在每年5—6 月份的丰水季节,绵疫病常暴发流行,发生面积数万公顷,产量损失达35%以上 (文献数据来源于地方农业局)。因生产上缺乏有效抗病品种,目前化学防治仍是防治该病的最主要措施[6]。长期以来,甲霜灵一直是防治绵疫病等卵菌病害的特效药剂,但疫霉菌等易对其产生抗药性,部分地区疫霉菌已对该药产生了抗药性[7-10],因此,近年来转而使用其他类型的杀菌剂来防治该病害。嘧菌酯是先正达公司根据天然产物strobilurins A 仿生合成的一种甲氧基丙烯酸酯类杀菌剂,其作用机制是通过抑制细胞色素b 和c1 之间的电子传递而阻止细胞的ATP 合成,从而抑制其线粒体呼吸而发挥杀菌作用[11]。嘧菌酯杀菌活性高、杀菌谱广、持效期长、高效低毒,对卵菌病害具有良好的杀菌活性[12-14],但由于其作用位点单一,因此,病原菌对其易产生抗性[15]。目前国内外尚未见有关番茄绵疫病菌对嘧菌酯的敏感性报道。为掌握番茄绵疫病菌对该药抗性现状,本研究测定了番绵疫病菌对嘧菌酯的敏感性,并对其抗性风险进行分析,以指导田间科学用药和杀菌剂抗性管理。
1 材料与方法
1.1 供试材料
1.1.1 供试培养基
10% V8 培养基:将100 mL V8营养汁、0.2 g CaCO3、900 mL 去离子水和18 g琼脂粉置于三角瓶中,摇匀,在121 ℃高压蒸汽灭菌锅中灭菌20 min。待冷却至60 ℃时倒入培养皿中。
选择性培养基:待上述培养基冷却至50 ℃左右时,加入利福平20 mg、氨苄青霉素200 mg 和制霉菌素100 mg,混匀,倒入平皿中。
1.1.2 供试菌株 于2018—2019 年在江西于都、安义、万载、瑞昌、余江、贵溪和乐平等番茄主产区的绵疫病重发田块采集典型病果,先用自来水冲洗3 min,再用灭菌水冲洗。在病健交界处切取0.5 cm × 0.5 cm 方形小块,经75%酒精和3%次氯酸钠消毒后置于选择性培养基上。待形成微小菌落后,在其边缘切取菌丝块,移入不含抗菌素的10% V8 培养基中培养。如此重复3 次,得到纯培养物。共分离纯化得到58 株菌株,根据形态学特征和ITS 测序结果,鉴定病原菌均为辣椒疫霉Phytophthoracapsici Leonian。将菌株转接至试管斜面上,加入灭菌石蜡油覆盖菌面,在10 ℃左右避光保存备用。菌株命名采用地名汉语拼音首字母+序号,如YX1 表示永新地区1 号菌株。
1.1.3 试验药剂 96% 嘧菌酯 (azoxystrobin)原药,由先正达 (苏州) 作物保护有限公司提供。试验时先用少许丙酮溶解并配制成质量浓度为100 μg/mL 的母液,待10% V8 培养基溶化后,并冷却至50 ℃时,加入适量母液,充分振荡摇匀后倒入培养皿中,即制得含梯度浓度药剂的平板。
1.2 试验方法
1.2.1 辣椒疫霉对嘧菌酯的敏感性检测 采用菌丝生长速率法[16]测定番茄辣椒疫霉对嘧菌酯的敏感性。先进行预备试验,将供试菌株活化后转接到含 0、0.1、1、10、100 μg/mL 嘧菌酯的 10%V8 培养基中,25 ℃黑暗培养7 d 后,根据菌丝生长情况将嘧菌酯质量浓度梯度设置为0.05、0.15、0.45、1.35 和4.05 μg/mL,以不加农药作为对照。在菌落边缘打取直径5 mm 的菌饼,菌面朝下置于含系列质量浓度嘧菌酯的培养基平板中央。每处理重复3 次。在生化培养箱中于25 ℃黑暗培养5~7 d,当对照中菌落直径达到7 cm 以上时,用十字交叉法测量各处理的菌落增长直径,计算抑制率。将抑制率转化成几率值,求出毒力回归方程y = a + bx,根据毒力回归方程计算有效抑制中浓度 (EC50) 和相关系数 (r)。
1.2.2 抗性突变体的获得及抗性水平的测定 将敏感菌株YD5 在10% V8 培养基上于25 ℃下培养7 d 后,在黑暗条件下,将菌落置于距紫外灯 (20 W、波长254 nm) 下方约20 cm 处照射120 s,保持菌落处于黑暗环境中60 min, 以降低光修复。用直径为5 mm 的打孔器在菌落边缘迅速打取菌饼,转接至含10 μg/mL 嘧菌酯的10% V8 培养基上,立即置于恒温培养箱中于25 ℃下黑暗培养,能够迅速生长的为疑似突变体。将这些疑似突变体转移到不含嘧菌酯的10% V8 培养基上活化培养3 代,转接至含 10 μg/mL 嘧菌酯的 10% V8 培养基上验证其抗药性。将各抗性突变体转接至含不同质量浓度嘧菌酯的10% V8 培养基上培养,计算药剂对菌丝生长的抑制率 (%),并求出毒力回归方程y = a + bx 和 EC50值 (μg/mL)。
根据各突变体的EC50值与其亲本菌株 (YD5)的比值计算抗性倍数,根据抗性倍数将各突变体的抗性水平划分为敏感、低抗、中抗和高抗[17],其中:抗性倍数≤3 的为敏感菌株 (S);3<抗性倍数≤10 的为低抗菌株 (LR);10<抗性倍数≤100 的为中抗菌株 (MR);100<抗性倍数的为高抗菌株(HR)。
1.2.3 突变体的抗性遗传稳定性 将抗性突变体转接到无药10% V8 培养基上连续转代培养8 代后,分别计算各突变体第2 代、第4 代、第6 代和第8 代对嘧菌酯的抗性倍数,根据其抗性倍数的变化情况,分析抗药性状在无药剂选择压力下的遗传稳定性。
1.2.4 抗性突变体的生物学性状
致病力:待抗性突变体和敏感菌株在10%V8 培养基上黑暗培养7 d 后,收集游动孢子制成每毫升含104个游动孢子的悬浮液,采用叶片喷雾法于番茄苗期5~6 叶时接种。单个菌株接种10 株,重复3 次。接种后24 h 内保持相对湿度100%、黑暗和25 ℃;此后保持相对湿度75%以上、光暗交替和25~28 ℃。7~9 d后调查病情,比较敏感菌株和突变体的致病力差异。
生长速率及产孢量:在抗性突变体和敏感菌株的菌落边缘打取直径5 mm 的菌饼,菌面朝下置于10% V8 培养基上于25 ℃下黑暗培养,每隔2 d 测量菌落直径,计算菌株平均生长速率。培养7 d 后用无菌水洗下孢子囊,置于4 ℃冰箱中预冷30 min,促使游动孢子充分释放,用血球计数板测定各菌株的游动孢子量。
孢子萌发率:将相同浓度的抗性突变体和敏感菌株的游动孢子悬浮液 (1 × 104个/mL) 100 μL均匀涂布于水琼脂培养基上,23 ℃培养24 h 后,在显微镜下观测每个处理的休止孢子,共观察10 个视野,计算孢子萌发率。
1.2.5 交互抗药性测定 采用菌丝生长速率法[18],测定敏感菌株YD5 及其突变体对烯酰吗啉和甲霜灵的敏感性 (EC50),分析嘧菌酯抗性突变体对烯酰吗啉和甲霜灵是否有交互抗性。
2 结果与分析
2.1 病原菌对嘧菌酯的敏感性
测定了2018—2019 年从江西各地采集分离到的58 个辣椒疫霉菌株对嘧菌酯的敏感性。结果(表1) 表明:嘧菌酯对辣椒疫霉菌丝生长有较强的抑制作用,其 EC50值介于 0.186 7~1.623 9 μg/mL之间,平均 EC50值为 (0.860 6 ± 0.331 8) μg/mL,EC50值最高值与最低值相差8.7 倍,菌株群体对嘧菌酯的敏感性差异不显著。从EC50值地理分布看,万载菌株平均EC50值最高,达1.022 2 μg/mL,其次为安义,平均EC50值为0.924 0 μg/mL,瑞昌和于都的最低,平均 EC50值分别为0.739 8 μg/mL和0.737 4 μg/mL,地区间差异较小。
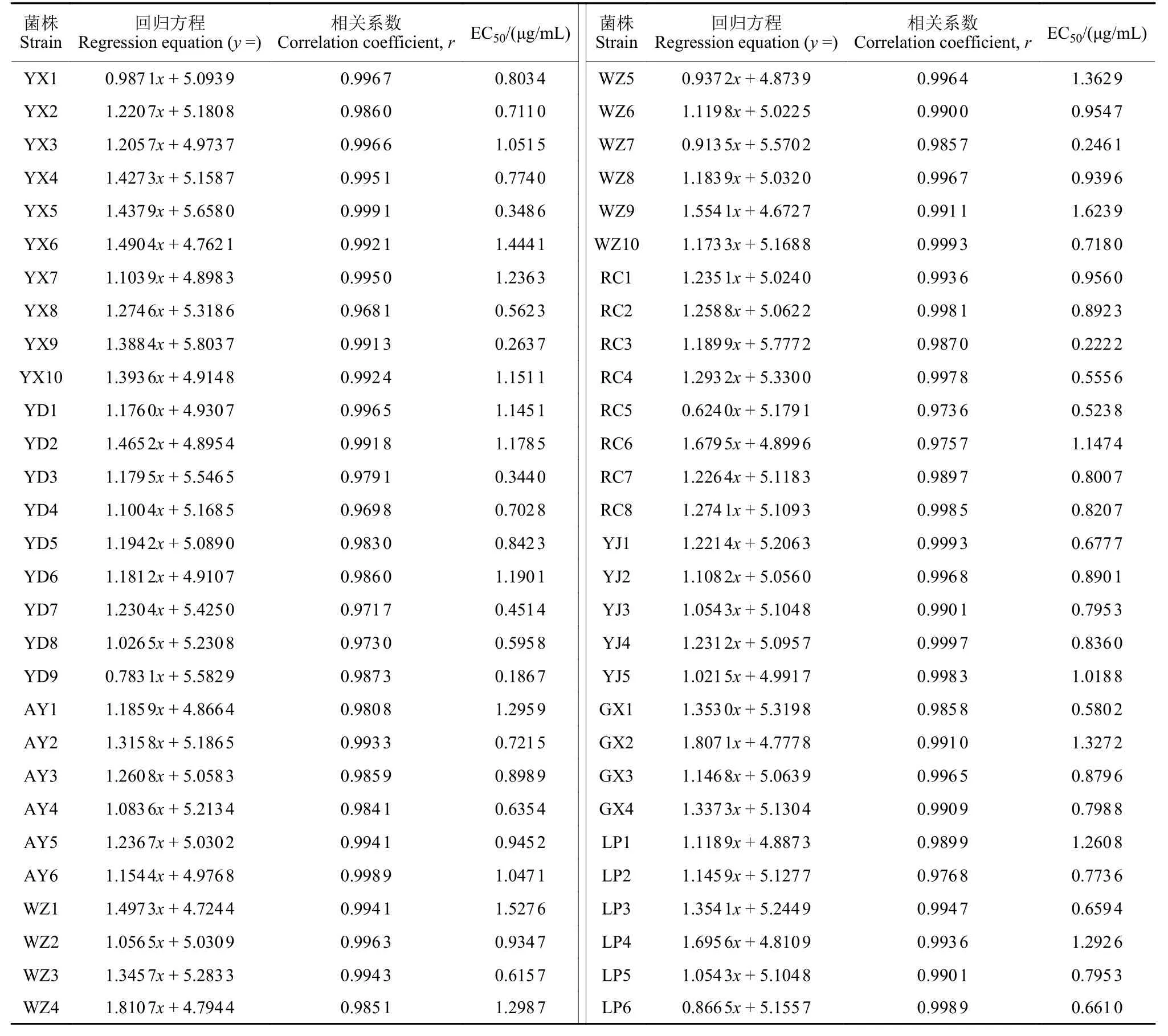
表1 江西辣椒疫霉对嘧菌酯的敏感性Table 1 Sensitivity to azoxystrobin of Phytophthora capsici in Jiangxi Province
对所测58 个菌株的EC50值进行阶段分布划分,统计每个阶段内菌株分布的数目及频率。结果显示 (图 1),EC50值在 0.15~0.45、>0.45~0.75、>0.75~1.05、>1.05~1.35 和>1.35~1.65 μg/mL 的菌株数分别为 6 株、15 株、21 株、12 株和4 株,分别占样本总数的10.34%、25.86%、36.21%、20.69% 和6.90%。从图中可以看出,58 个菌株对嘧菌酯的敏感性频率分布呈单峰曲线,接近正态分布,未出现抗药性亚群体,因此,本次测定的病原菌对嘧菌酯的平均EC50值(0.860 6 ± 0.331 8) μg/mL,可作为江西省辣椒疫霉对嘧菌酯的敏感性基线。
2.2 抗性突变体的获得及抗性水平
通过紫外线照射敏感菌株YD5 (EC50值接近敏感基线) 120 s 后,从20 000 个菌饼中获得4 个抗性突变体 (YD5-1、YD5-2、YD5-3 和 YD5-4),突变频率仅为0.02%,说明江西辣椒疫霉对嘧菌酯产生抗性变异的频率较低。
菌丝生长速率法测定的抗性突变体在含系列质量浓度 (0、1、2、5、20、10 和 20 μg/mL) 嘧菌酯V8培养基上的生长量结果见表2。4 个突变体的 EC50值在 4.452 1~7.812 1 μg/mL 之间,抗性倍数介于5.3~9.3 之间,均属于低抗水平。

表2 辣椒疫霉抗性突变体对嘧菌酯的敏感性Table 2 Sensitivity to azoxystrobin of resistant mutants of Phytophthora capsici
2.3 突变体的抗性遗传稳定性
经紫外线照射得到的4 个低抗性菌株YD5-1、YD5-2、YD5-3 和YD5-4,在无药培养基上连续转代培养8 代后发现,各抗性菌株的抗性水平变化不显著,抗性倍数均介于3~10 之间,仍为低抗菌株 (表3),说明上述4 个突变体的抗性能稳定遗传。
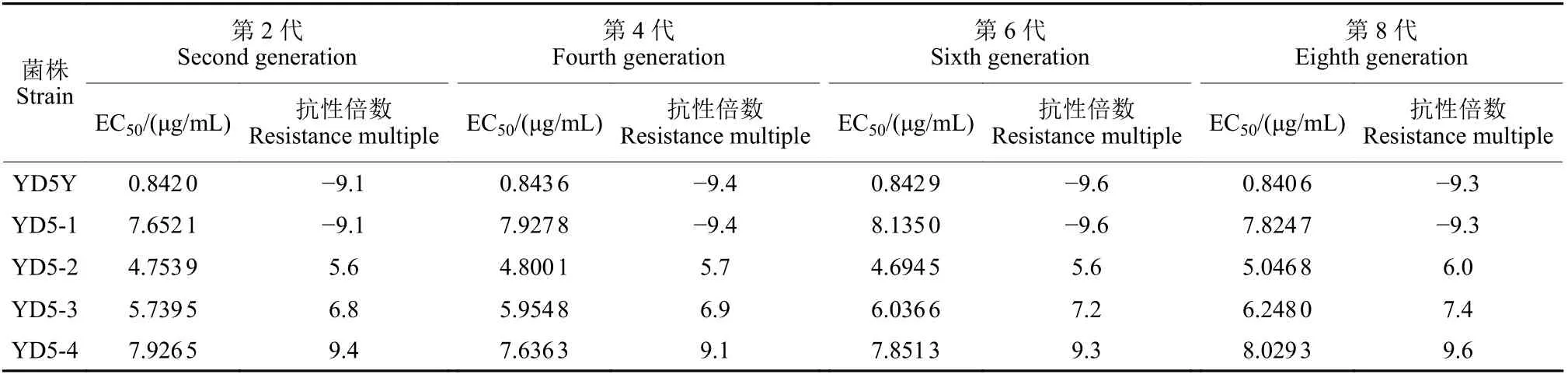
表3 番茄辣椒疫霉突变体对嘧菌酯抗性遗传稳定性Table 3 Genetic stability of resistance to azoxystrobin in the mutants of Phytophthora capsici
2.4 抗性突变体的生物学性状
从表4 可以看出:除突变体YD5-4 的致病力低于亲本菌株外,其余突变体与亲本菌株YD5 的致病力间不存在显著性差异;突变体YD5-2 的生长速率与亲本菌株相当,其余3 个突变体均显著下降;在产孢量方面,突变体YD5-3 与亲本菌株不存在显著性差异,而突变体YD5-1、YD5-2 和YD5-4 的产孢量显著低于亲本菌株,说明抗药性产生后其无性繁殖能力有明显变化;亲本菌株与所有抗性突变体的孢子萌发率不存在显著差异性。

表4 抗药突变体与亲本菌株间生物学性状比较Table 4 Comparison of biological traits between resistant mutants and their parent
2.5 交互抗药性
采用菌丝生长速率法测定了敏感菌株YD5及其4 个嘧菌酯抗性突变体对烯酰吗啉和甲霜灵的敏感性。结果表明,4 个抗性突变体对烯酰吗啉的抗性倍数为0.8~1.4,对甲霜灵的抗性倍数为0.8~1.3。4 个突变体对烯酰吗啉和甲霜灵的敏感性表现一定程度的降低或增加,说明嘧菌酯与烯酰吗啉和甲霜灵之间均不存在交互抗性 (表5)。

表5 突变体和敏感菌株YD5 对烯酰吗啉和甲霜灵的敏感性比较Table 5 Sensitivity comparison of resistant mutants and susceptible strain YD5 to dimethomorph and metalaxyl
3 结论与讨论
本研究结果表明:2018—2019 年从江西省番茄主产区采集分离得到的58 个辣椒疫霉菌株对嘧菌酯仍有较高的敏感性,其对嘧菌酯的敏感性基线为 (0.860 6 ± 0.331 8) μg/mL。58 个菌株的菌丝生长对嘧菌酯的敏感性频率呈正态分布,田间未出现抗性菌株,这与Emil 等[19]、Qin 等[20]和姜彦全等[21]关于疫霉菌的研究结果相符。鉴于此,目前江西地区仍可使用嘧菌酯防治绵疫病。但辣椒疫霉具有易变异性等特点,长期使用单一药剂有产生抗药性的风险,应定期加强田间菌株的抗药性监测,掌握病原菌的敏感性变化动态,适时制定防治策略。
为评估辣椒疫霉对嘧菌酯的抗性风险,本研究通过紫外线照射敏感菌株,成功诱导出4 个低抗菌株,未获得中、高抗菌株,抗性突变率为0.02%。对突变体的抗药性进行遗传稳定性分析,发现经过8 次转代培养后,其抗性水平变化不显著,抗性能够稳定遗传,且突变体的致病力与亲本菌株不存在显著性差异,多数突变体的生长速率与产孢量均显著低于亲本敏感菌株,而在孢子萌发率方面两者不存在显著性差异。综上可知,虽然目前生产上江西辣椒疫霉尚未出现抗嘧菌酯菌株,但存在抗性突变体产生的可能性,且这种抗性遗传很稳定,不易丧失,突变体致病力也不减弱,由此推断,病菌对嘧菌酯的抗性风险为中等,因此,生产上应该注意对嘧菌酯抗性的管理,应注意轮用或换用作用机理不同杀菌剂。
针对目前生产上常见的交互抗药性问题,本研究选用生产上防治番茄绵疫病的常规药剂烯酰吗啉和甲霜灵与嘧菌酯进行交互抗药性分析,证实嘧菌酯与烯酰吗啉和甲霜灵间均不存在交互抗性。郭梁等[22]和李成斌等[23]研究认为,疫霉菌对甲霜灵和嘧菌酯不存在交互抗性;袁善奎等[18]研究表明,疫霉菌对嘧菌酯和烯酰吗啉无交互抗性。张海良等[24]测定了江西辣椒疫霉菌对甲霜灵的敏感性,结果发现,108 个菌株均为敏感菌株,未出现抗药性病原菌亚群体,且在生产中甲霜灵对卵菌病害防效较为显著。因此,建议生产上可将该药剂与烯酰吗啉、甲霜灵以及其他作用机理不同的杀菌剂混用或交替使用,以缓解杀菌剂对抗药性的选择压力,延长药剂的使用期限。
猜你喜欢
作物学报(2022年2期)2022-11-06
亚热带农业研究(2022年1期)2022-08-08
农药学学报(2022年3期)2022-06-14
作物学报(2022年8期)2022-05-29
农业工程(2022年1期)2022-04-11
蔬菜(2021年7期)2021-11-27
新农民(2020年5期)2020-12-10
农民致富之友(2020年18期)2020-06-19
安徽农学通报(2019年14期)2019-09-03
农民致富之友(2019年14期)2019-05-24
