
绿潮迁移过程中环境要素与浒苔藻体生物学特征的相关性分析*
2021-02-03 01:44侯承宗唐学玺
海洋与湖沼 2021年1期
管 晨 郝 雅 侯承宗 唐学玺, 王 影,①
(1. 中国海洋大学海洋生命科学学院 青岛 266100; 2. 青岛海洋科学与技术试点国家实验室 海洋生态与环境科学功能实验室 青岛 266237)
绿潮(green tide)是一种常见的有害藻华(HABs),主要由石莼属(Ulva)等海藻的过度生长引起(Fletcher,1996; Anderson et al, 2002)。绿潮的暴发多与当地水体富营养化有关, 其暴发严重威胁当地的水产养殖和旅游业, 造成巨大的经济损失(Sun et al, 2008)。目前, 黄海已成为世界上绿潮规模最大的地区。黄海绿潮的主要优势种为浒苔(Ulva prolifera) (张晓雯等,2008; 丁兰平等, 2009; Gao et al, 2013)。自 2007 年开始, 我国黄海海域均出现大规模绿潮。目前最为科学家们广泛接受的观点为黄海南部的苏北浅滩筏架养殖区是黄海浒苔绿潮的起源地。每年4 月中旬至5 月初, 浒苔藻体脱离固着基, 形成小块漂浮浒苔并出现在黄海南部江苏沿海, 在风场和海流的作用下逐渐向北漂浮迁移, 生物量迅速增加; 在 5 月下旬, 聚集的漂浮浒苔形成大面积绿藻斑块; 6 月中旬, 漂浮浒苔逐渐在山东半岛堆积; 7 月浒苔生物量达到最大后开始下降, 并最终消亡于山东半岛海域(Hu et al,2010; Liu et al, 2013; Wang et al, 2015)。
南黄海海域环境复杂多变, 尤其是营养盐的浓度变化剧烈(王俊杰等, 2018)。黄海冷水团、黄海暖流等海流显著影响南黄海营养盐的来源、分布和输运(Wei et al, 2016)。射阳河、灌河、长江等河流输入的氮和磷为南黄海海域绿潮的暴发提供了物质基础(马洪瑞等, 2010)。南黄海营养盐的分布通常呈现出由近岸到远海逐渐降低的趋势, 其中在苏北近岸海域呈现出了较高的氮、磷含量(高嵩等, 2012)。南黄海海州湾附近的35°N 断面是南黄海最长的断面。35°N 北部海水透明度增加, 营养盐含量较低, 而 35°N 南部海域营养盐含量丰富。在南黄海, 春、夏季水温上升,温、盐跃层的出现使得水体稳定性增加, 营养盐含量呈现上层低、下层高的趋势, 环境条件适宜, 有利于绿潮的发展(韦钦胜等, 2012)。有研究表明, 在 35°N断面附近, 漂浮浒苔生物量具有明显增加的特征(孔凡洲等, 2018)。
与其他大型海藻相比, 浒苔生态幅较宽, 具有广盐、广温等较强适应环境的特征。浒苔对海水温度、盐度和光照强度的适应范围分别为 10—30°C、7.2—53.5 和1000—10000 lx; 最适范围分别为 15—25°C、20.2—26.9 和 5000—6000 lx (吴洪喜等, 2000)。浒苔具有较强的适应环境的能力, 其生物学特征受到环境变化的影响。在正常盐度条件下, 低温降低了浒苔的生长, 高温显著降低了浒苔的最大光化学量子产量Fv/Fm; 相对于温度变化, 盐度变化对浒苔Fv/Fm影响不显著(刘榆莎等, 2016)。在营养盐丰富的环境中, 浒苔吸收营养盐的能力显著高于其他多数海藻, 浒苔也可以迅速吸收多种形式的营养盐, 而随着环境中营养盐含量的增加, 浒苔藻体可以迅速吸收并储存大量的营养盐用于藻体快速生长(吴晓文等, 2010)。温度、光照和盐度均可以影响浒苔硝态氮的吸收能力,在 20°C 时, 浒苔吸收硝态氮的速率最大, 盐度与浒苔硝态氮的吸收呈正相关; 在低光条件下, 浒苔对硝态氮的吸收速率较低, 随着环境中营养盐含量的增大, 浒苔吸收硝态氮的速率显著增加(丛珊珊, 2011)。不同温度、盐度和光照强度可以影响浒苔孢子和配子的萌发和放散(王浩东等, 2012)。浒苔孢子囊的形成与光合活性有关(Gao et al, 2010)。通过对南黄海浒苔孢子囊形成的情况进行了研究, 发现在江苏沿海海域, 浒苔藻体呈现出较好的生理状态, 但呈现出较少的孢子囊, 而随着浒苔向北漂浮迁移, 藻体生理活性下降, 孢子囊形成比例升高(郑阵兵等, 2018)。
前期针对浒苔绿潮形成的环境驱动机制进行了大量的研究工作, 在暴发海区富营养化对浒苔绿潮形成的驱动作用已形成统一的共识(高嵩等, 2012;Wang et al, 2015; 白雨等, 2019)。浒苔具有很强的环境可塑性, 其自身存在诸多可以适应环境变化的生物学特征, 如光合活性的改变、营养盐吸收的改变、形态改变等。在浒苔绿潮暴发过程中, 尤其是数百公里的迁移过程中, 浒苔藻体时刻面临生态要素剧烈变化的影响。浒苔藻体的环境可塑性是否与生态要素存在相关性关系还需进一步明确。本文通过现场调查的方式探究了绿潮迁移过程中环境要素与浒苔藻体生物学特征的相关性以期甄别驱动浒苔绿潮形成的主要环境因子, 为阐明为什么黄海会暴发大规模浒苔绿潮提供理论依据。
1 材料与方法
1.1 站位布设与样品采集
浒苔样品采集于2018 年5 月25—30 日, 根据监测数据此时黄海绿潮从江苏近岸到青岛近岸呈长条状斑块分布, 早期迁移斑块已抵近青岛近岸, 但还未集聚(国家海洋局北海预报中心)。采集范围在(34.0°—36.0°N, 120.0°—121.5°E), 共设 6 条断面进行大面站调查, 其中断面 A、B 和 C 位于海州湾南部(SH), 断面 D、E 和 F 位于海州湾北部(NH), 采样站位如图1 所示。采集不同站位的绿藻, 通过无菌海水反复冲洗藻体, 同时用刷子清洗藻体表面并去除泥沙等杂物。随机挑选浒苔样品, 现场测量叶绿素荧光参数、NRA 和生殖分配率。

图1 浒苔采样站位点Fig.1 Map of sample collecting stations
1.2 环境参数的测量
温度、光照、盐度等环境参数在每天同一时段(10:00—11:00)进行测量。温度和盐度使用温盐深仪(conductivity-temperature-depth profiler, CTD, Seabird 25, Sea-Bird, USA)测量。光照强度使用照度计(TES1335, 泰仕电子工业有限公司, 中国台湾)测量(单位: lx), 然后依据Thimijan (Thimijan et al, 1983)所述方法将光照强度转化为 400—700 nm 的光合可利用辐照[photosynthetically active radiation, PAR, 单位: μmol photons/(m2·s)]。水样经过 GF/C 膜(Whatman,US)过滤, 储存于-80°C, 全自动连续流动分析仪(AutoAnalyzer3, Seal Analytical, Germany) 测量溶解无机氮(DIN)和磷酸盐(-P ), DIN 和-P 的测定方法参照《海洋监测规范》(GB17378. 4—2007)。
1.3 浒苔暴发海域藻体光合特征测定
采用Dual-PAM-100 荧光仪(Walz, Germany)测定浒苔样品的光合活性。实验开始前, 预先设置好仪器程序, 将浒苔暗适应 20 分钟。测定光合诱导曲线并记录 Fv/Fm和 Y(II)。Fv/Fm表示光系统 II (PSII)在藻类中的最大光化学量子产量(其中Fv是黑暗中最大可变叶绿素荧光, Fm是黑暗中最大叶绿素荧光), Y(II)表示光系统II (PSII)的实际光化学量子产量。各种参数均重复 5 次, 每次测量在同处理的不同藻体上进行,叶绿素荧光参数计算的基本步骤按照 Wang 等(2012)和Heinz Walz GmbH 操作手册进行。
1.4 浒苔暴发海域藻体硝酸还原酶活性的测定
硝酸还原酶活性(Nitrate reductase activity, NRA)的测定按照Young 等的方法(Young et al, 2005)。称取采集的浒苔样本于研钵中研磨并添加提取液聚乙烯吡咯烷酮和 1 mmol/L 二硫苏糖醇的磷酸缓冲液, 在4°C 条件下 5 000 r/min 离心 5 min 后, 加入 0 .1 mol/L的 KNO3溶液和 NADH 溶液, 混匀后再 25°C 水浴30 min, 随后加入醋酸锌和乙醇终止反应, 最后用萘乙二胺显色法在显色15 min 后再540 nm 的波长测定吸光值。NRA 单位为μmol/L/(mg·h)。
1.5 浒苔暴发海域藻体生殖特征测定
清洁后的浒苔悬浮于放有无菌海水样品盘中。将浒苔剪为 1 mm×1 mm 左右大小放置于载玻片上,用光学显微镜RVL-100-G (ECHO, USA)观察细胞并统计含有孢子或配子的生殖细胞和营养细胞数。生殖分配率(RA)=生殖细胞数/营养细胞数计算(Wang et al, 2019)。
1.6 数据处理与分析
本章所提供的数据以平均值±标准差(mean ± SD)表示。数据统计分析使用IBM SPSSStatistics22 (SPSS Inc, USA)中的独立样本 T 检验(Student’s t-test)和单因素方差分析(one-way ANOVA) 进行, 显著性水平设置为0.05。其中P 值小于0.05 表明差异具有统计学意义, P<0.05 认为差异显著, P<0.01 认为差异极显著。数据首先采用 Levene 检验数据的方差齐性(homogeneity), 数据正态性(normality) 使用Shapiro-Wilk 检验。当组间方差不齐时, 采用Dunnett’s T3 对数据进行计算, 其他情况下采用Duncan post-hoc test 进行处理。使用 Origin 软件(OriginLab, USA)和 Arcgis 软件(Esri, USA)作图。冗余分析(RDA)使用Canoco 5 作图。
2 结果与分析
2.1 海水表层部分环境要素测量
分别对6 个断面的海水表层的温度、盐度、光照强度、DIN、-P进行了测量, 图2 所示, 在浒苔绿潮漂浮迁移所经过的海域中, 温 度和光照 強度总体呈现南高北低、盐度北高南低的特征。DIN 和-P在A1、B1 和C1 时最高, 总体呈现南高北低和近岸高, 远海低的特征。通过对调查海域海水表层环境要素的测量发现, 35°N 断面附近的环境要素有较为明显的变化。表1 所示, SH 海水表层的温度、盐度、光照强度、DIN 和-P分别为(21.76±0.24)°C、(30.75±0.48)、(394.29±17.18) μmol photons/(m2·s)、(17.43±5.62) μmol/L 和(0.27±0.09) μmol/L。NH 海水表层的温度、盐度、光照强度、DIN 和-P分别为(20.72±0.39)°C、(31.18±0.54)、(355.11±59.9) μmol photons/(m2·s)、(5.16±0.63) μmol/L 和(0.10±0.02) μmol/L。SH 海水表层的温度、DIN 和-P都显著高于NH海水表层(Student’s t-test, P < 0.05), SH 和 N H 海水表层的盐度和光照强度没有显著差异(Student’s t-test, P> 0.05)。
2.2 浒苔暴发海域藻体光合特征测定
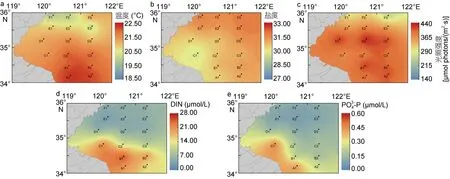
图2 浒苔迁移过程中海水表层部分环境要素Fig.2 Environmental factors in surface during the Ulva prolifera drifting
表1 海州湾南部(SH)和海州湾北部(NH)海水表层温度、盐度、光照强度、DIN 和 -P的测量Tab.1 Temperature, salinity, light intensity, DIN and -P in southern Haizhou Bay (SH) and northern Haizhou Bay (NH)
注: 同一列中不同上标字母表示数值之间有显著差异(P<0.05), 相同上标字母表示无显著差异(P>0.05)
海域 温度(°C) 盐度 光照强度[μmol photons/(m2·s)] DIN (µmol/L) PO34 --P (µmol/L)SH 21.76±0.24a 30.75±0.48a 394.29±17.18a 17.43±5.62a 0.27±0.09a NH 20.72±0.39b 31.18±0.54a 355.11±59.92a 5.16±0.63b 0.10±0.02b

图3 浒苔迁移过程中藻体的光合参数Fig.3 Photosynthetic parameters of Ulva prolifera during drifting
通过对采集的浒苔藻体进行光合活性的测定,藻体 Fv/Fm和 Y(II)的变化如图 3 所示, 浒苔藻体Fv/Fm和 Y(II)总体呈现南高北低的特征。断面 A、B和C 分布的浒苔藻体的Fv/Fm显著高于断面D、E 和F 的浒苔(图 3a, one-way ANOVA, P < 0.05)。断面 A、B 和 C 浒苔藻体的 Fv/Fm值在 0.55—0.7 之间, 表明漂浮初期浒苔藻体的光合活性较好, 而断面 D、E 和 F藻体的 Fv/Fm值在 0.45—0.60 之间, 表明漂浮一段距离后浒苔藻体光合活性较差, 受胁迫程度相对较高。浒苔藻体Y(II)呈现出与Fv/Fm相似的趋势。图3 表明,断面 A、B 和 C 藻体 Y(II)值在 0.4—0.55 之间, 而断面 D、E 和 F 藻体的 Y(II)值在 0.25—0.5 之间, 表明NH 海域的浒苔藻体的实际光合活性较差。在同一断面上, 不同站位的Fv/Fm和Y(II)值呈近岸高, 远海低的特征, 但站点间无显著差异。表明同一纬度、不同经度的浒苔光合活性存在差异, 整体趋势与营养盐浓度变化趋势类似。在向北迁移过程中, 浒苔藻体Fv/Fm和 Y(II)总体呈现下降趋势。SH 海域浒苔的Fv/Fm和 Y(II)显著高于 NH 浒苔(图 3c 和 3d, Student’s t-test, P <0.05), 在 SH, 藻体 Fv/Fm和 Y(II)的平均值分别为 0.63 和 0.52, 而在 NH, 藻体 Fv/Fm和 Y(II)的平均值分别为0.48 和0.39。
2.3 浒苔暴发海域藻体硝酸还原酶活性的测定
通过测定不同断面浒苔藻体的 NRA, 可以反映藻体对硝态氮的吸收和代谢水平。图4 表明, 断面A和B 的浒苔藻体NRA 显著高于断面D、E 和F 的藻体(图 4 a, one-way ANOVA, P < 0.05), 而断面 C 浒苔藻体的NRA 与其他断面浒苔NRA 的变化均不显著。浒苔NRA 总体呈现南高北低的特征, SH 海域浒苔藻体 N RA 显著高于 N H 藻体(图 4 b, Student’s t-test, P<0.05)。在 SH, 藻体 NRA 的平均值为 2.29 μmol/L/(mg·h), 而在 N H, 藻体 Fv/Fm和 Y (II)的平均值为 0 .66 μmol/L/(mg·h)。
2.4 浒苔暴发海域藻体生殖特征测定
本次实验统计了断面A、B、C、D、E 和F 漂浮浒苔的生殖分配率。结果表明, 浒苔藻体形成的生殖细胞数目呈先上升, 后下降的趋势, 在断面B 时生殖分配率最低, 断面 D 时生殖分配率最高。断面 D 的浒苔藻体生殖细胞数目显著高于断面 B 浒苔藻体生殖细胞数目(图5a, one-way ANOVA, P <0.05), 而其他断面之间均无显著性变化。浒苔藻体生殖分配率整体呈现上升趋势, SH 海域藻体生殖分配率显著低于NH 藻体(图 5b, Student’s t-test, P <0.05)。在 SH, 藻体生殖分配率的平均值为 27%, 而在 NH, 藻体生殖分配率平均值为38%。

图4 浒苔迁移过程中藻体的NRA 的测定Fig.4 NRA of Ulva prolifera during the drifting

图5 浒苔迁移过程中藻体的生殖分配率Fig.5 RA of Ulva prolifera during drifting
2.5 环境要素与浒苔生物学特征的关系
图 6 显示了不同断面浒苔的生理参数与环境要素的 RDA 分析图, 参与 R DA 分析的环境要素有温度、光照强度、盐度、DIN 和-P, 参与 P CA 分析的生物学特征有Fv/Fm、Y(II)、NRA 和生殖分配率。环境要素对浒苔生物学特征的总解释度为 85.4%,DIN、温度、-P、盐度和光照强度的解释度分别为80.1%、72.1%、61.0%、49.0%、26.1%和25.5%, 说明DIN 是驱动浒苔生物学特征改变的主要环境要素。表2 显示了SH 和NH 海域的环境要素与浒苔藻体的生理和生殖参数的相关性分析结果, 具体说明如下:
(1) SH 和NH 海域DIN 与纬度呈极显著负相关(R=-0.819, P<0.01), 说明浒苔绿潮向北迁移过程中海水表层的 D IN 逐渐降低; DIN 与温度和-P呈极显著正相关(P<0.01), DIN 与光照强度和盐度的相关性不显著(P>0.05); 温度、光照强度和-P与纬度呈极显著负相关(P<0.01), 温度与光照强度呈现出显著正相关(P<0.05), 与-P呈显著正相关(P<0.05),盐度与纬度呈显著正相关(P<0.05), 说明纬度越高,温度、光照强度和-P越低, 盐度越高。

图6 海州湾南部和北部海域浒苔的生理参数与环境要素的RDA 分析图Fig.6 Redundancy analysis (RDA) of Ulva prolifera samples and physiological parameters and environmental factors
(2) SH 和 NH 海域 DIN 与浒苔 Fv/Fm(R=0.853)和 Y(II) (R=0.814)呈极显著正相关(P<0.01), 结果表明海水表层 DIN 浓度越高, 浒苔的光合作用效率越高。藻体 Fv/Fm和 Y(II)与纬度呈极显著负相关(P<0.01), 与温度和光照强度呈极显著正相关(P<0.01),与-P 呈显著正相关(P<0.05), 而盐度与 Fv/Fm相关度不显著(P>0.05), 与Y(II)呈显著负相关(P<0.05)。
(3) SH 和 NH 海域 DIN 与浒苔 NRA 呈极显著正相关(R=0.897, P<0.01), 表明海水表层DIN 浓度越高,NRA 越高; 藻体NRA 与纬度呈极显著负相关(P<0.01),与温度呈极显著正相关(P<0.01), 与-P 和光照强度呈显著正相关(P<0.05), 与盐度呈显著负相关(P<0.05)。NRA 与 Fv/Fm和 Y(II)呈极显著正相关(P<0.01)。
表2 海州湾南部和北部温度、盐度、光照强度、DIN 和 -P与浒苔生理特征和生殖特征相关性分析Tab.2 Correlation analysis between temperature, salinity, light Intensity, DIN, -P, physiology and reproduction of Ulva prolifera in SH and NH

表2 海州湾南部和北部温度、盐度、光照强度、DIN 和 -P与浒苔生理特征和生殖特征相关性分析Tab.2 Correlation analysis between temperature, salinity, light Intensity, DIN, -P, physiology and reproduction of Ulva prolifera in SH and NH
注: *表示 P<0.05; **表示 P<0.01。
DIN 纬度 温度 光照强度 PO34 --P 盐度 Fv/Fm Y(II) NRA RA DIN 1 -0.819** 0.726** 0.366 0.703** -0.379 0.853** 0.814** 0.897** -0.717**纬度 1 -0.912** -0.702** -0.626** 0.516* -0.946** -0.895** -0.906** 0.420温度 1 0.553* 0.554* -0.473 0.824** 0.853** 0.852** -0.451*光照强度 1 0.173 -0.653** 0.655** 0.633** 0.506* -0.275 PO --P 1 -0.583* 0.589* 0.594* 0.702* -0.467盐度 1 -0.468 -0.597* -0.513* -0.220 Fv/Fm 1 0.923** 0.920** -0.443 Y(II) 1 0.905** -0.408 NRA 1 -0.542*RA 1 34
(4) SH 和NH 海域DIN 与浒苔生殖分配率呈极显著负相关(R=-0.717, P<0.01), 表明海水表层 D IN 浓度越高, 生殖分配率越低。藻体生殖分配率与温度和NRA 呈显著负相关(P<0.05), 与纬度、光照强度、-P和盐度相关性不显著(P>0.05)。
3 讨论
黄海浒苔绿潮是一种典型的跨区域海洋生态灾害, 在其长距离迁移过程中, 经历复杂的生态学过程,受多种环境要素驱动(Wang et al, 2015; Liu et al,2016)。本文分析了浒苔绿潮迁移路径上温度、光照强度、盐度、DIN、-P 等环境要素与浒苔藻体光合活性、NRA 及生殖分配率之间的相互关系, 探究了环境要素与浒苔光合、营养盐吸收和生殖等关键生物学特征的相关性, 为最终阐明为什么黄海会暴发大规模浒苔绿潮提供了理论依据。
浒苔绿潮迁移海区温度、盐度、营养盐等环境要素变化显著, 盐度略有升高且变化不显著。江苏近岸海域主要受到苏北沿岸流、黄海冷水团、黄海暖流和长江冲淡水等海流的影响, 苏北近岸海域营养盐处于较高浓度(Wei et al, 2016)。因此, 苏北近岸海域营养盐高值区与低盐区相对应, 营养盐浓度整体呈现为南高北低, 近岸高, 远海低的趋势, 盐度整体呈现南低北高的趋势。王俊杰等(2018)测量了苏北海域海水表层营养盐浓度, 发现 6—8 月 DIN 浓度上升至28 µmol/L 以上,-P 浓度上升至 0.6 µmol/L 以上。在夏季, 海水表层受热增温, 随着纬度的升高,温度和光照强度逐渐降低, 白雨等(2019)对2008—2017 年绿潮密集区域海水表层温度和光照强度分析也得出了类似的结果。
光合作用是植物能量的来源之一, 也是植物重要的生理过程。叶绿素荧光可以反映浒苔藻体的生理状态以及对环境的适应能力。本文发现, 浒苔在向北漂移过程中, 浒苔的光合活性略有上升, 随后显著下降。光合作用与 D IN 呈极显著正相关, 与-P呈显著正相关, 高浓度的 DIN 对浒苔光合活性的影响高于-P。浒苔生长过程中会消耗大量的营养盐,相比磷, 浒苔对氮的需求更高(吴晓文等, 2010)。环境中氮浓度的变化对浒苔有较大的影响, 而磷对浒苔的影响不如氮明显。在缺乏磷的条件下, 浒苔光合作用受到的影响较小(Villares et al, 2004)。温度可以影响浒苔的光合活性。过高或过低的温度均不利于正常的光合活性(张涛等, 2017)。本文发现, 温度对浒苔藻体光合活性的影响呈极显著正相关, 表明浒苔向北迁移, 纬度升高, 温度降低, 与光合作用相关的酶活性可能下降, 抑制了浒苔的光合活性。光照强度是影响浒苔体内叶绿素 a、b 含量的重要因素, 较弱的光照显著抑制浒苔藻体的生长和酶反应速率(丛珊珊,2011)。随着纬度的升高, 光照强度整体呈现下降趋势,光合活性降低浒苔藻体捕获的光能下降。盐度与浒苔藻体的 Y(II)值呈显著负相关, 表明在漂浮过程中,浒苔光合作用对盐度的变化也较为敏感。
硝酸还原酶是硝态氮转化为可利用氨氮的限速酶, 对氮同化起到主要调控。而硝酸还原酶容易受到外在因素影响。NRA 直接影响植物对环境中硝态氮的利用, 影响植物的生长。表2 的相关性分析结果表明, DIN、温度、-P、盐度和光照对 N RA 按影响大小排列的顺序为 DIN>温度>-P>盐度>光照。硝酸还原酶是一种底物诱导酶, 其活性受到环境中的氮、磷浓度尤其是硝态氮的影响。NRA 会随着氮磷比或氮浓度的升高而增大(王金花, 2008)。环境中温度和光照强度也可以影响NRA 的大小。硝酸还原酶是一种蛋白质, 对温度的变化较为敏感, 温度过高或过低均不利于硝酸还原酶保持较高活性(陈月红,2015)。光照强度可以通过光合磷酸化调控ATP, 影响浒苔营养盐的吸收, 从而影响NRA。
生殖分配是植物生活史的核心组成部分, 不同的生殖分配通常反映了植物在自然选择下的不同策略(Reekie et al, 2005)。生殖分配率的变化对浒苔适应环境变化起着关键作用。在本实验中, 生殖分配率结果表明在向北漂浮过程中, 浒苔藻体逐渐形成生殖细胞, 并释放孢子或配子, 为生物量的快速积累提供可能。DIN 对浒苔的生殖分配率的影响极显著, 温度与生殖分配率呈显著负相关, 而-P、盐度和光照强度对浒苔生殖分配率的影响未达到显著水平。温度可以影响浒苔孢子/配子的放散和萌发。浒苔孢子/配子放散的最适温度为 25°C, 浒苔孢子/配子萌发的最适温度为20°C (韩红宾等, 2015)。浒苔在向北迁移过程中, 环境温度逐渐降低, 接近藻体孢子/配子萌发的最适温度, 促进了生殖分配率的升高。浒苔在盐度为10 的条件下对配子的放散有极显著影响, 在40 盐度的条件下对孢子的放散有显著影响(王浩东等,2012)。盐度对浒苔藻体生殖分配率无显著的影响, 本文推测, 在向北漂浮迁移过程中, 盐度通过影响浒苔光合作用间接影响藻体生殖细胞的形成, 且浒苔主要放散的为孢子。
4 结论
猜你喜欢
中国畜牧业(2022年20期)2022-11-19
环境工程技术学报(2022年3期)2022-06-05
江苏农业科学(2019年20期)2019-12-23
中国集体经济(2017年11期)2017-04-24
绿色科技(2016年24期)2017-03-30
河北渔业(2017年1期)2017-02-21
新会计(2016年11期)2017-01-04
中学理科园地(2016年6期)2017-01-03
西部金融(2015年7期)2015-10-29
青岛科技大学学报(社会科学版)(2015年1期)2015-05-22
