
雷州半岛东部近岸海域大型底栖甲壳动物群落结构及其影响因子
2021-02-03 10:18陈志劼吕少梁陆丽仪曾嘉维王学锋
广东海洋大学学报 2021年1期
陈志劼,吕少梁,陆丽仪,张 静,曾嘉维,王学锋
(1.南方海洋科学与工程广东省实验室(湛江),广东 湛江 524025;2.广东海洋大学水产学院,广东 湛江 524088)
预测物种的资源密度及其分布格局,是生态学研究的主要内容之一。作为海洋生态系统的重要组成部分,虾、蟹、虾蛄类甲壳动物种类多、数量大,是底栖动物的优势类群[1],其资源变动对海洋生态系统的稳定会产生重要影响[2-3]。随着近海渔业资源的衰退和鱼类等捕捞对象减少以及人们对海洋产品需求的增加,海洋甲壳动物因繁殖力强、生活史较短等特征在捕捞业和渔业经济中的地位不断提高,自20 世纪80 年代以来成为主要的捕捞对象之一[4]。根据1990―2016 年《中国渔业统计年鉴》,甲壳动物海洋捕捞总产量1990 年88.13 万t,至2016年升至239.63 万t,整体呈波动上升趋势[5]。因此,对甲壳动物资源开展定期调查与评估,是了解其资源动态及可持续开发利用的基础。
雷州半岛东部近岸海域潮汐特征复杂,几乎不受入海河流影响,海水盐度较稳定,为海洋生物提供了适宜的生境[6-7]。该海域不仅是多种海洋经济动物产卵、育幼及索饵的场所,也是国家一级保护动物中华白海豚 (Sousa chinensis) 主要栖息场所之一[8-9]。目前,关于雷州半岛东部近岸海域生物群落结构研究主要集中在鱼类[9-10]和浮游生物[11-12];在甲壳类方面,仅见肖汉洪等[13]硇洲岛潮间带蟹类组成及其分布的研究和张旭丰等[14]虾拖网副渔获种类组成的研究。硇洲岛海域国家级海洋牧场示范区为本项目研究海域的一部分,研究该海域甲壳动物群落结构及生态因子的调控作用,可为海洋牧场建设效果评估、增殖种类选择、增殖容量研究等提供基础数据。
1 材料与方法
1.1 数据来源
2016 年4 月、8 月、11 月和2017 年2 月于雷州半岛东部近岸海域(110°30′―110°45′E,20°40′―21°05′N)进行渔业资源底拖网调查,具体站位见图1。依据《海洋渔业资源调查规范》(SC/T 9403―2012)[15],选取选择性能小的单船型底拖网作为调查网具,网囊网目长度25 mm。受海况等因素的影响,各站位的实际拖速、拖网时间有偏差,因此在数据分析前先做标准化处理,即统一换算成拖速3 n mile/h 情况下每小时的渔获量(kg/h)。
采集拖网渔获物中的甲壳动物样品,参照文献[16-18]完成种类鉴定,统计其数量和质量(0.01 g)等。所有站位同步采集环境数据:利用SM-5A 测深仪测量水深(d),YSI 水质参数仪测定溶解氧(DO)、pH、盐度(S)和水温(t)。各站位采集500 mL 水样,利用分光光度法测定叶绿素a(Chl a)浓度。
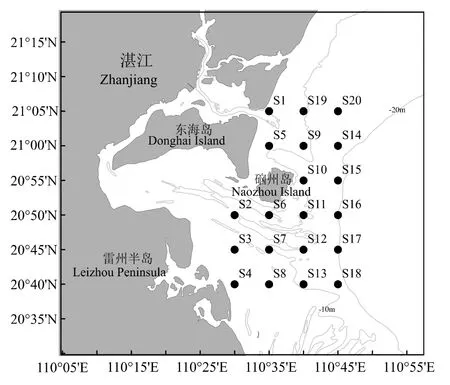
图1 雷州半岛东部近岸海域调查站位Fig.1 Survey stations in the east coastal of Leizhou Peninsula
1.2 数据处理与分析
1.2.1 质量资源密度及环境数据的处理 甲壳动物质量资源密度用扫海面积法计算:
Pi=Ci/(ai q)。
式中,Pi指第i站的甲壳动物资源密度 (质量密度单位:kg/km2);Ci指第i站甲壳动物的每小时渔获质量(kg/h);ai指第i站拖网的每小时扫海面积(km2/h);q指捕获率,本研究取0.5,原因有:1)捕捞系数是基于拖网渔具选择性而得,与渔具规格,海区底质类型、海流等有关;2)为与鱼类渔获物保持一致,增加数据可比性,便于后期对整体渔获物的比较分析;3)捕获样品以小规格虾蟹类居多。
为平衡群落中优势种极化作用及偶见种的影响,分析前对数据进行标准化处理。即剔除出现频率低于10%的种类,四次方根转换各站位甲壳动物的质量资源密度,环境数据除pH 外做lg (x+1)处理[4,9]。
1.2.2 优势种分析 利用Pinkas[19]相对重要性指数(IRI)确定甲壳动物优势种:
IRI=(N+W)×F×10 000,
式中,N为某一甲壳动物的数量比例 (%),W为某一甲壳动物的质量比例 (%),F为某一甲壳动物某站位的出现频率 (%)。将IRI>1000 的物种定为优势种。
利用种类更替率 (T) 研究优势种的季节变化:
T(%)=(a+b-2c)/(a+b-c),
式中,a、b为两个相邻季节的优势种种类数;c为两个相邻季节的共有优势种种类数。
1.2.3 多样性分析 采用Margalef物种丰富度指数(D)、Shannon-Wiener 多样性指数 (H′)、Pielou 均匀度指数 (J′) 分析甲壳动物的多样性,计算公式:
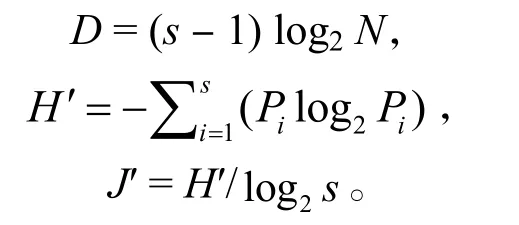
式中,s为甲壳动物种类数,Pi指第i种物种占甲壳动物总物种数的比例,N为甲壳动物总数。
1.2.4 群落结构分析 利用Primer 5.0 软件进行聚类分析 (Cluster) 和参数多维标序排序 (MDS),分析研究海域甲壳动物的群落聚集性。采用协强系数(Stress) 衡量MDS 二维排序图结果:当Stress<0.05时,图形代表性较佳;0.05<Stress<0.1 时,排序较佳;0.1<Stress<0.2 时,图形有一定解释意义。
采用单因子相似性分析 (Anosim test) 检验甲壳动物群落在不同站位、不同季节间的差异显著性;采用相似性百分比分析 (Simper test) 计算各季节群落组内相似性和组间相异性的平均贡献率,并列出贡献率前五的物种。
1.2.5 群落特征与环境因子的冗余分析 利用R统计软件[20](Vegan package) 分析甲壳动物群落分布与环境因子的关系。首先,对各站位甲壳动物资源密度矩阵 (站位×各甲壳动物的质量密度) 进行去趋势对应分析 (DCA),分析结果中,如梯度长度(Lengths of gradient)最大值超过4,则使用单峰模型(CCA)排序;若小于3,则选择线性模型(RDA)排序;若介于3~ 4,二者均可使用。之后采用蒙特卡洛置换检验 (Monte Carlo permutation test) 分析环境因子对甲壳动物分布影响的显著性。
2 结果与分析
2.1 种类组成
渔获物样品共鉴定出甲壳动物98 种 (含未定种1 种),隶属2 目24 科43 属(表1)。各季节种类数依次是夏季(74 种)>冬季(62 种)>秋季(59 种)>春季(50 种)。其中,对虾科 (Penaeidae)和梭子蟹科 (Portunidae) 的种类数较多,分别为23种和27 种,是调查海域甲壳动物的主要类群。四季共有种为变态蟳 (Charybdisvariegata)、断脊小口虾 蛄(Oratosquillinainterrupta)、哈氏仿对虾(Parapenaeopsishardwickii) 等30 种,占甲壳动物种类数的30.6%。
2.2 优势种
根据相对重要性指数 (IRI) 分析,四季均出现的优势种为变态蟳、哈氏仿对虾、须赤虾(Metapenaeopsis barbata) 等10 种(表2)。优势种的季节变化显著(P<0.05),春―夏季、夏―秋季和秋―冬季间优势种更替率分别为50.6%、29.5%和38.7%。
2.3 生物多样性
甲壳动物的多样性指数(H’)、均匀度指数(J’)和丰富度指数(D)均为夏季最高,冬、春季次之,秋季最低(表3)。
2.4 聚类分析
四季甲壳动物群落结构的协强系数 (Stress) 为0.10~0.18,较好地反映了群落间的相似度(图2)。其中,春季的S2 站位和秋季的S17 站位与同季其他站位相似性较低,暂不纳入群落分组。相关原因有待进一步研究。
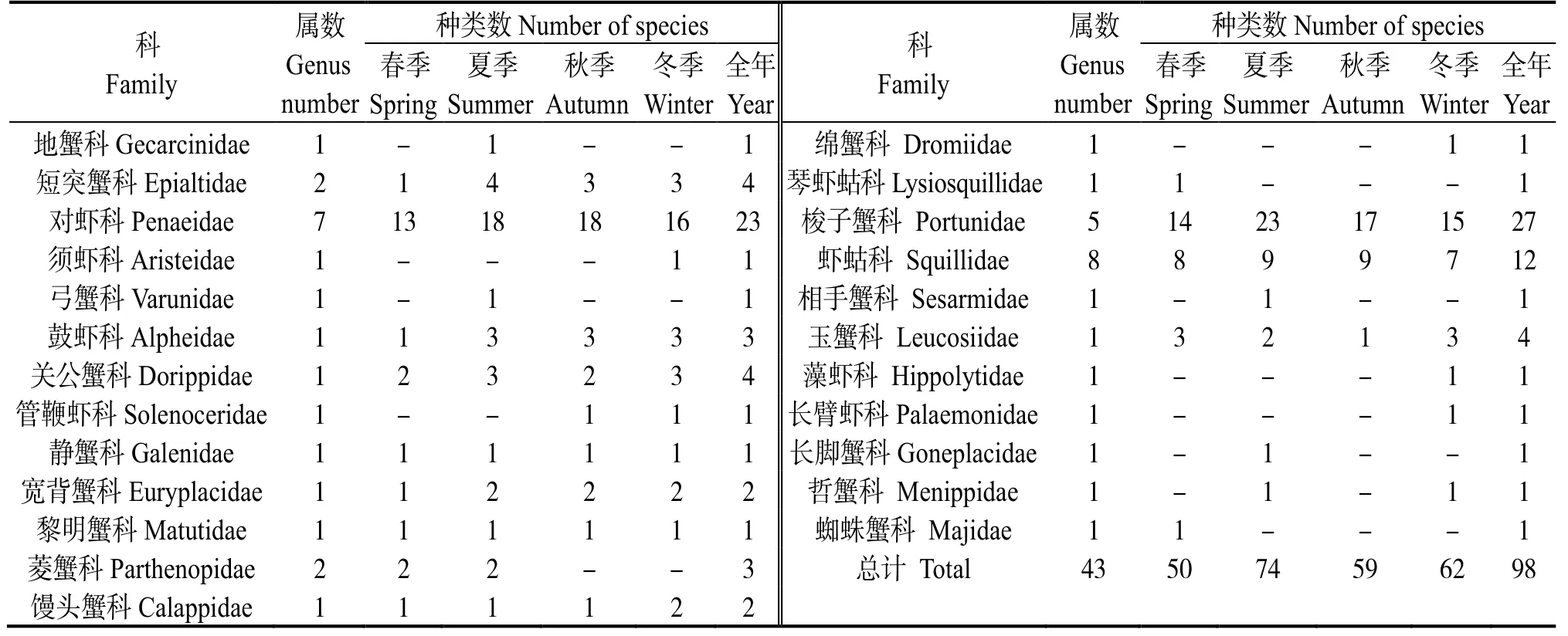
表1 雷州半岛东部近岸海域甲壳动物的科、属和种Table 1 Composition of crustaceans in the east coastal of Leizhou Peninsula
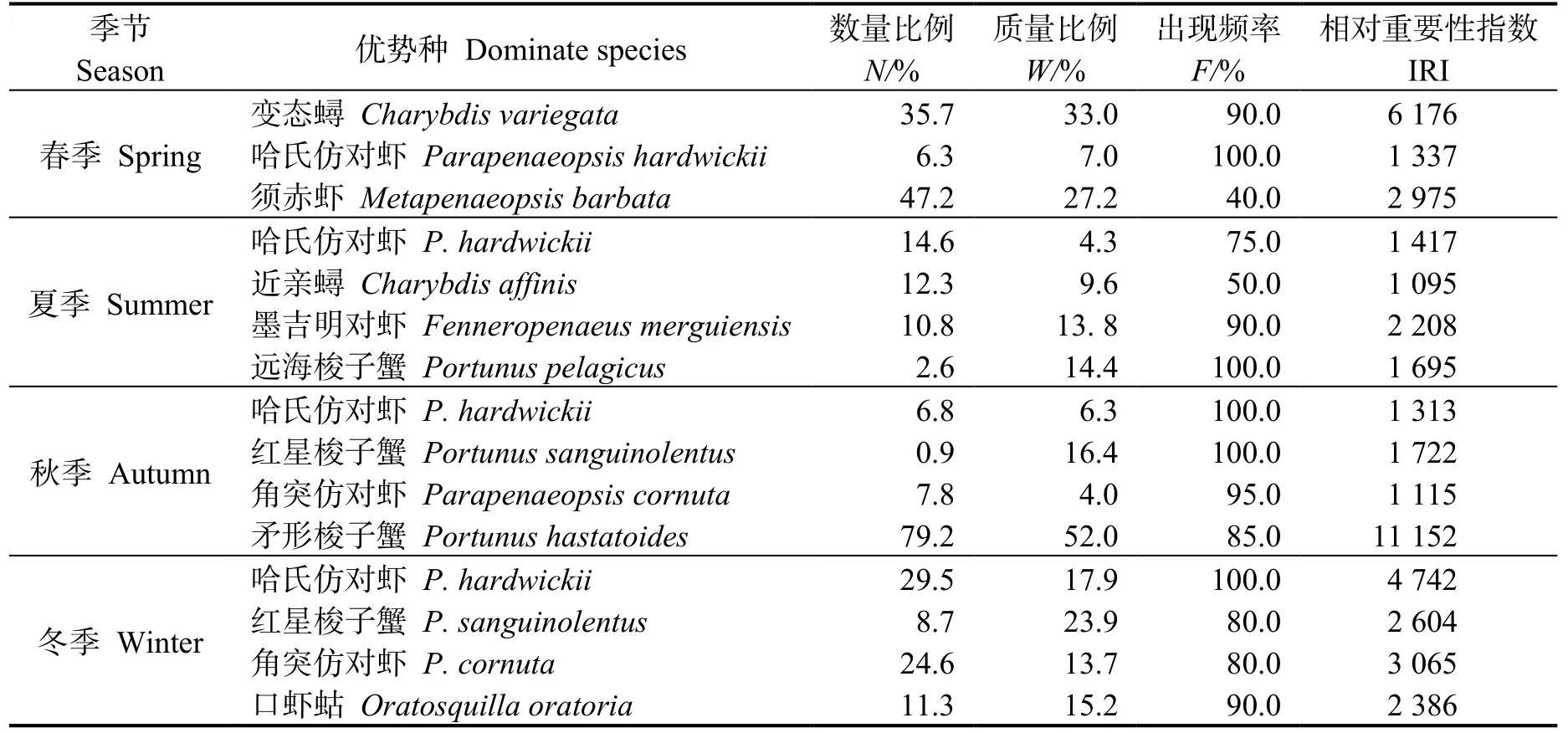
表2 各不同季节优势种的相对重要性指数(IRI)Table 2 IRI index of dominant species in different season in the east coastal of Leizhou Peninsula

表3 生物多样性指数的季节变化Table 3 Seasonal change of the diversity index in the east coastal of Leizhou Peninsula
四个季节甲壳动物均可划分为2个群落(图2),春季群落Ⅰ主要分布在10 m 等深线附近,以哈氏仿对虾、远海梭子蟹(Portunus pelagicus)、红星梭子蟹等为主;群落Ⅱ主要分布在20 m 等深线附近,以须赤虾、口虾蛄、伍氏平虾蛄(Erugosquilla woodmasoni)等为主;夏季群落Ⅰ位于该海域的西南一侧,靠近雷州半岛,以近亲蟳、远海梭子蟹、墨吉明对虾等为主;群落Ⅱ位于东侧及北部,以猛虾蛄(Harpiosquilla harpax)、远海梭子蟹、墨吉明对虾等为主;秋季群落Ⅰ位于西南一角的浅滩范围,以红星梭子蟹(P.sanguinolentus)、远海梭子蟹、哈氏仿对虾等为主;群落Ⅱ以矛形梭子蟹、红星梭子蟹、哈氏仿对虾等为主;冬季群落Ⅰ主要位于南部,以红星梭子蟹、角突仿对虾、哈氏仿对虾等为主;群落Ⅱ主要位于20 m 等深线附近,以口虾蛄、伍氏平虾蛄、哈氏仿对虾等为主。
2.5 群落相似性与相异性
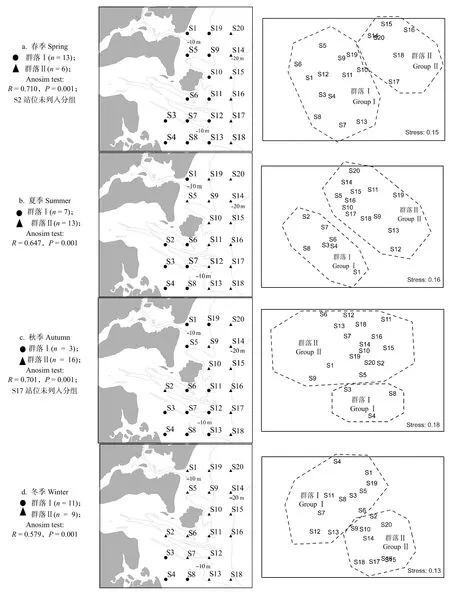
图2 不同季节甲壳动物的聚类分析和MDS 排序Fig.2 Cluster analysis and MDS sequence diagram of crustaceans in different seasons in the east coastal of Leizhou Peninsula
表4 表明,春季群落I、II 的组内相似性分别为44.5%、54.8%,群落Ⅰ主要特征种为哈氏仿对虾等,群落Ⅱ主要特征种为须赤虾等。夏季群落Ⅰ、Ⅱ的组内相似性分别为54.4%、53.8%,群落Ⅰ主要特征种为近亲蟳等,群落Ⅱ主要特征种为远海梭子蟹等。秋季群落Ⅰ、Ⅱ的组内相似性分别为62.7%、59.8%,群落Ⅰ主要特征种为哈氏仿对虾等,群落Ⅱ主要特征种为红星梭子蟹等。冬季群落Ⅰ、Ⅱ的组内相似性分别为47.3%、53.8%,群落Ⅰ主要特征种为哈氏仿对虾等,群落Ⅱ主要特征种为哈氏仿对虾等,贡献率前5 位的物种见表4。
春、夏、秋、冬四季群落Ⅰ与群落Ⅱ的相异性(表5)分别为70.6%、58.1%、50.2%和63.0%。春季群落间的主要分歧种为须赤虾等,夏季为近亲蟳等,秋季为猛虾蛄等,冬季为伍氏平虾蛄等。贡献率前5 位的物种见表5。
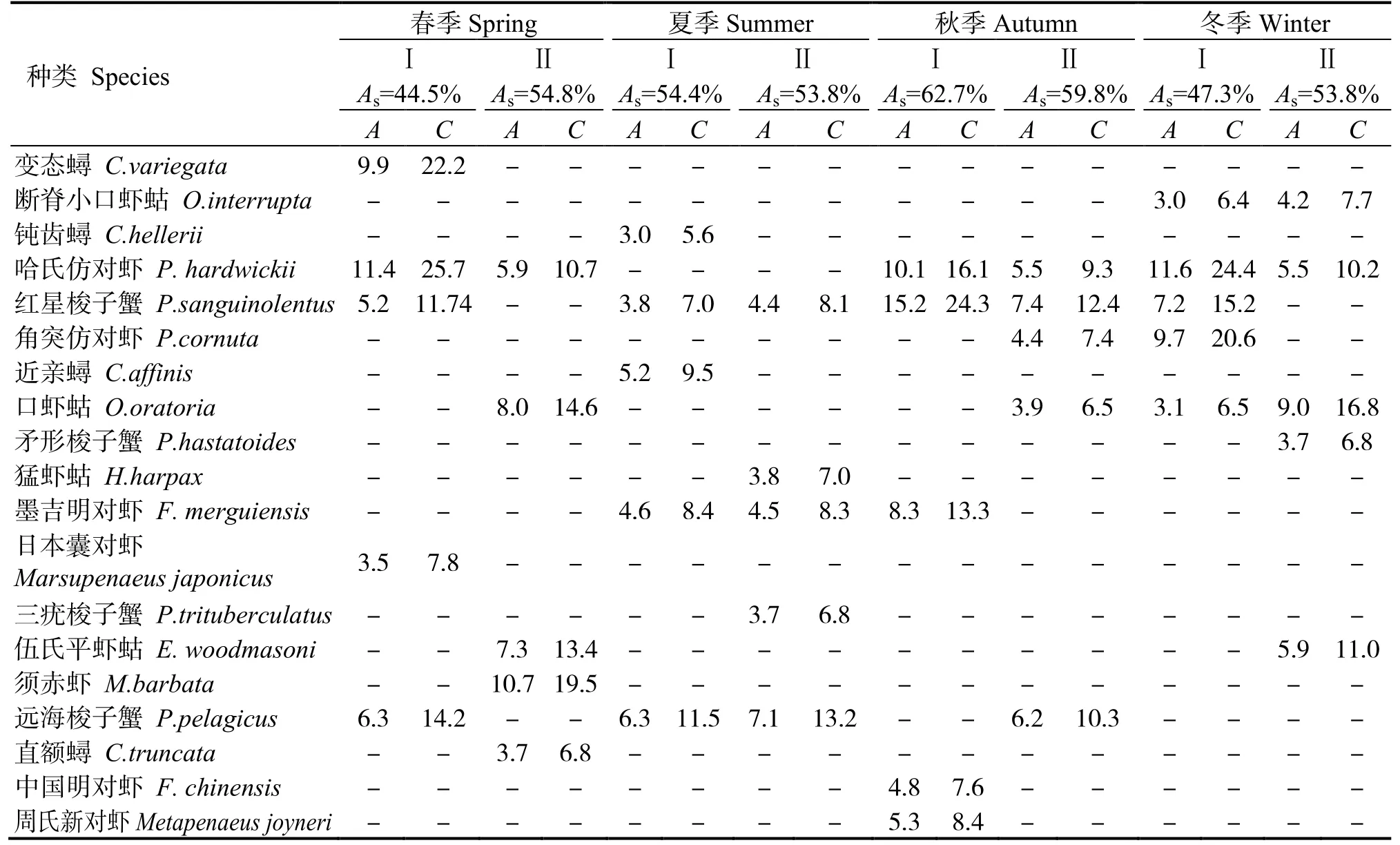
表4 不同季节群落平均相似性及其贡献率前5 位的种类Table 4 Top five representative species and their contributions to the average within-group similarity in different seasons in the east coastal of Leizhou Peninsula %
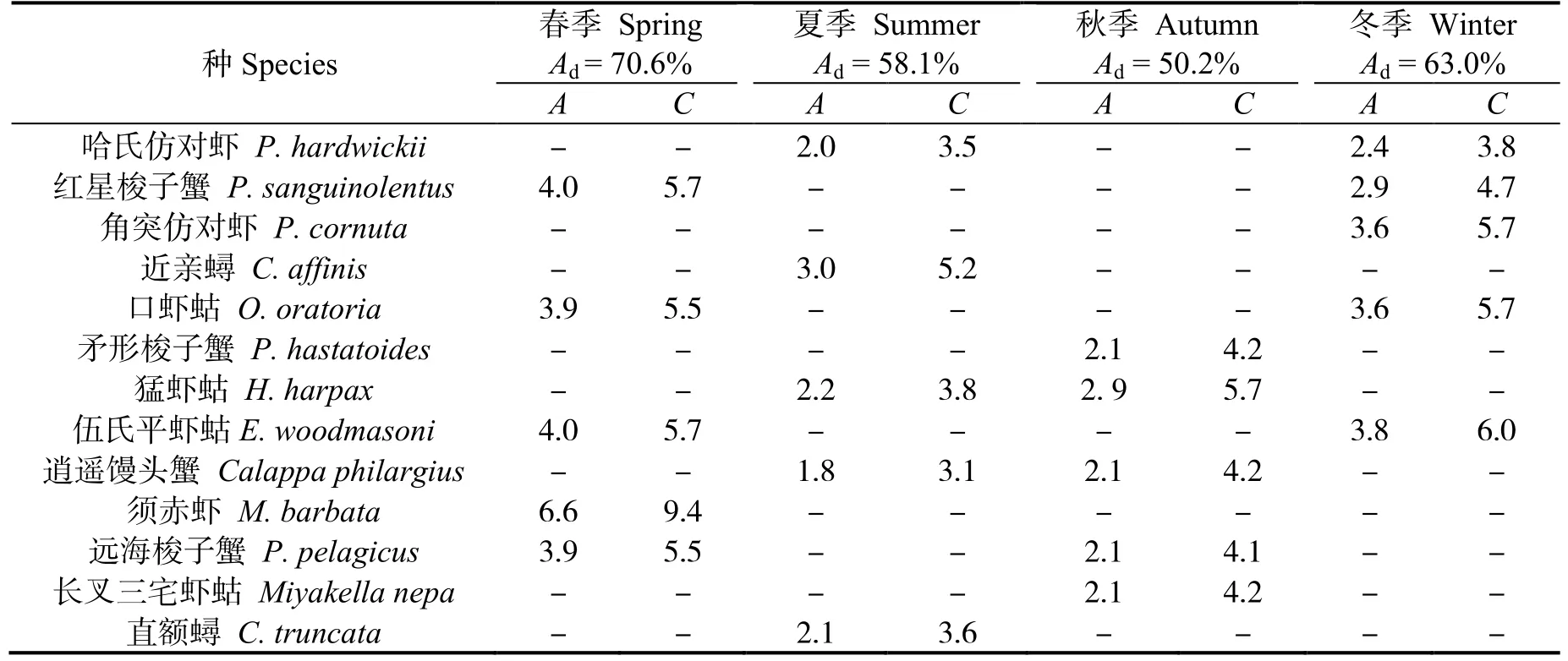
表5 各季节群落间甲壳动物平均相异性及其贡献率前5 的种类Table 5 Top five representative species and their contributions to the average between-group dissimilarity in different seasons in the east coastal of Leizhou Peninsula %
2.6 群落结构与环境因子的关系
DCA 分析表明,梯度长度的最大值均小于3,因此选用RDA 排序方法。当各季节甲壳动物群落-环境多元回归线性关系的R2经Ezekiel 公式[21]校正后的值无限接近于零或为负数时,环境因子对甲壳动物群落的解释能力不能优于初始时随机生成正态分布的解释变量[22],因此采用初始RDA 分析结果(表6)。
甲壳动物不同季节物种-环境相关性在RDA1轴(特征值e=1.7223~4.0220)、RDA2 轴(特征值e=0.8675~ 1.9030)分别为44.3%~ 51.3%、15.8%~ 26.2%(图3)。
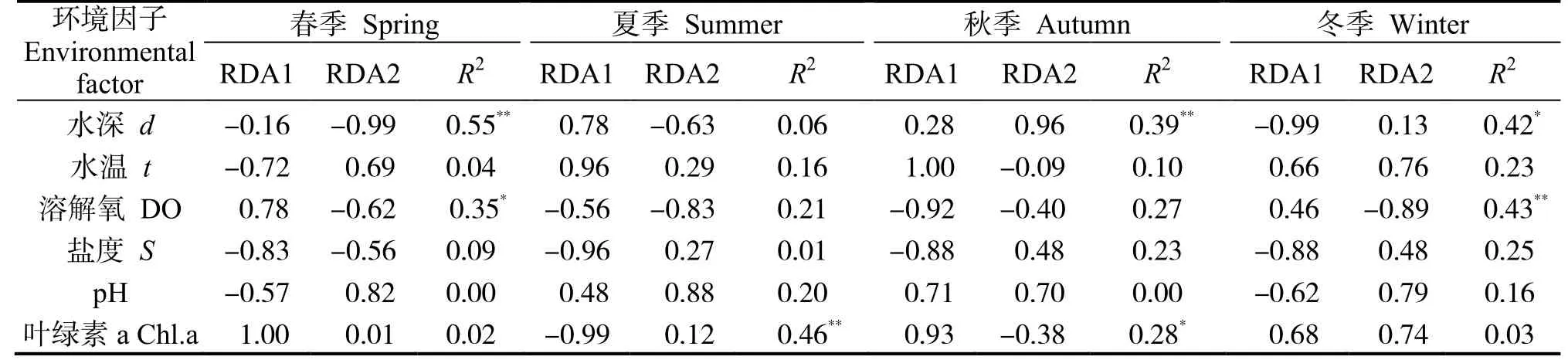
表6 环境因子的相关显著性检验Table 6 Significance test of environmental factors

图3 群落分布与环境因子的RDA 排序Fig.3 RDA triplots of crustaceans community and environmental factors
春季各环境因子对各站位甲壳动物群落结构(种类组成与密度)的贡献率为32.8%,其中水深与溶解氧是主要影响因子。水深对群落Ⅱ中S14、S15 和S16 站位的甲壳动物分布有极显著影响,呈正相关,对群落Ⅰ靠近雷州湾及湛江港出港口位置甲壳动物分布的影响呈正相关。溶解氧对群落Ⅰ南侧站位的甲壳动物分布影响较大,呈正相关,对群落Ⅱ除S17 外其他站位的影响呈正相关。
夏季各环境因子对甲壳动物群落结构的贡献率为31.6%,其中,叶绿素a 是主要影响因子。叶绿素a 对群落Ⅱ甲壳动物分布有极显著的影响,呈正相关,对群落Ⅰ东侧站位的影响呈正相关。
秋季各环境因子对甲壳动物群落结构的贡献率为30.8%,其中,水深与叶绿素a 是主要影响因子。叶绿素a 对群落Ⅰ甲壳动物分布有极显著的影响,呈正相关,对群落Ⅱ S1、S9、S2、S6、S7 和S12 站位的影响呈正相关。水深对群落Ⅰ甲壳动物分布的影响呈负相关,对群落Ⅱ S2、S5、S10、S13和东侧站位的影响呈正相关。
冬季各环境因子对甲壳动物群落结构的贡献率为39.5%,其中,溶解氧与水深是主要影响因子。溶解氧对群落Ⅰ S1、S6、S12、S13 站位甲壳动物分布的影响呈正相关,对群落Ⅱ S14、S15、S16、S17 和S18 站位有极显著的影响,呈正相关。水深对群落Ⅱ的影响呈正相关,对群落Ⅰ除S13 外其他站位的影响呈正相关。
3 讨论
3.1 甲壳动物群落结构特征
3.1.1 种类组成 本研究发现甲壳动物的种类数(98 种)与邻近纬度地区的珠江口附近海域[23](54种)、大亚湾口海域[24](52 种)和柘林湾海域[25](53种)以及中纬度地区的乐清湾海域[26](43 种)、苍南近海海域[27](45 种)和舟山海域[4](42 种)相比,更具优势。一般认为,低纬度地区的生物多样性水平高于高纬度,且捕捞强度是重要因素。研究海域处于北回归线以南的低纬度地区,受热带和亚热带季风气候影响,温度、盐度稳定,日照时间长,饵料生物资源充足,因此种类数亦较丰富[9-12]。此外,亦与研究海域的面积、海底地形和航次等因素有关。
雷州半岛东部近岸海域全年优势种有10 种,包括生态类型为暖水广盐性的变态蟳、近亲蟳、红星梭子蟹、墨吉明对虾、远海梭子蟹和矛形梭子蟹,广温广盐性的哈氏仿对虾、口虾蛄、须赤虾和角突仿对虾。优势种季节更替方面,春-夏季变化较显著,更替率达50.6%,其他相邻季节变化也较明显。可能是春-夏季交替期间,温度回升,甲壳动物活动频繁,且正值多数甲壳动物的产卵高峰期,如哈氏仿对虾主要汛期为2―5 月和8―11 月,矛形梭子蟹5―6 月常见抱卵。其中,哈氏仿对虾在各个季节均为优势物种,可能与其生命周期短、繁殖能力强和生长迅速等特性有关[28]。
3.1.2 生物多样性 多样性指数 (H′) 常作为评价水域生态环境健康的指标之一,也用于评价生物群落受自然因素和人为因素共同影响的程度(0<H′<1 为重度影响;1<H′<3 为中度影响)[29]。本研究中,甲壳动物多样性指数 (H′) 均值为2.37,说明其群落分布受到中等程度的自然、人为因素影响;丰富度指数 (D) 季节均值为4.77,比柘林湾[25](D=2.45)、乐清湾[26](D=1.64)和舟山海域[4](D=1.37)等物种丰富程度高,可能与该海域的自然环境优越、生物群落稳定性高和人类选择性捕捞鱼类过度等因素有关。四季中以夏季的生物多样性指数均为最高,而秋季的生物多样性水平却最低。夏季正值休渔季节,海洋生物受人类干扰程度最小,生物多样性水平较高;秋季正值伏季休渔结束,开渔期人们在巨大利益驱动下对海洋生物资源影响最大,致使该季节生物多样性水平较差。这充分说明伏季休渔制度对甲壳动物资源具有重要养护作用,雷州半岛东部近岸海域甲壳动物资源及生态环境的跟踪监测与休渔结束后的渔业管理政策需要加强,以促进休渔效果的累积和资源的合理开发。
3.1.3 群落划分 雷州半岛东部近岸海域甲壳动物群落结构四季变化明显,四个季节均可划分为两个群落,MDS 二维标序图、Anosim test 和Simper test结果均表明群落划分的合理性。四季中,除春季群落组间相异性远高于组内相似性外,其余三季节差距均较小。春季群落Ⅱ以须赤虾等为主,主要特征种亦是须赤虾,且群落组内分歧种为须赤虾,表明须赤虾对春季群落划分直接相关。
3.2 甲壳动物群落结构与环境因子的关系
RDA 将对应分析与多元回归分析相结合,用以显示物种群落与环境因素之间关系[29]。本研究RDA 排序结果表明,水深、溶解氧和叶绿素a 是不同季节影响该海域甲壳动物群落分布的主要环境因素,且以水深的影响较大。
水深与溶解氧是春季和冬季甲壳动物群落的主要影响因子。冬季温度相对较低,海水溶解氧随温度的降低而升高,甲壳动物开始从沿岸浅滩往深水区迁徙;春季气候回暖,海水溶解氧开始降低,且实测数据显示,冬、春两季溶解氧含量相差较大,促使甲壳动物往沿岸浅水区域洄游。甲壳动物营底栖生活,随所处水深的环境不同,海洋的理化性质(温度、盐度等)亦会产生较大的变化[9],而溶解氧限制其生理耐受性[31],影响了甲壳动物栖息地的选择与生活方式,导致不同地域甲壳动物的生物量和群落分布有较大差异。受限于实测数据,本研究仅有表层水温、盐度数据,可能造成结果有一定误差,但考虑到同一断面的综合影响,亦有相近研究将表层温、盐度数据纳入影响范围,故未将其剔除,后续调查应增加底层环境因子的监测。
叶绿素a 是影响夏、秋季甲壳动物群落分布的主要环境因素。雷州半岛东部近岸海域三面环陆,海底地形复杂、地质稳定,这为底栖生物提供良好的孕育场[32],研究海域日照时间充足,夏、秋两季日照时长尤其明显,且受到近年来气候的变化和环境污染(工、农业发展、海水养殖)等因素影响,初级生产力资源丰富甚至达到水体富营养化水平,进而驱动着次级生产力的生物量变化[9,11,33],最终对甲壳动物的资源变动、群落分布产生影响。
猜你喜欢
当代水产(2022年8期)2022-09-20
中国农业科技导报(2022年4期)2022-06-14
当代水产(2022年5期)2022-06-05
科技视界(2021年21期)2021-08-24
当代水产(2021年4期)2021-07-20
科学与信息化(2020年11期)2020-06-19
意林·少年版(2020年5期)2020-04-14
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
漫画月刊·哈版(2017年7期)2018-01-29
