
柑橘果实枯水的生理及分子机制研究进展
2021-02-02 05:44:14吴黎明宋放王策王志静何利刚彭抒昂蒋迎春刘继红
华中农业大学学报(自然科学版) 2021年1期
吴黎明,宋放,王策,王志静,何利刚,彭抒昂,蒋迎春,刘继红
1.湖北省农业科学院果树茶叶研究所,武汉 430064; 2.华中农业大学园艺林学学院,武汉 430070
柑橘枯水是柑橘类果肉汁胞的生理失调症,是一种生理性病害,常常发生在果实成熟和采后贮藏过程中,导致果实品质和果实贮藏性能严重下降。柑橘枯水主要分为汁胞粒化型枯水和汁胞萎缩型枯水,粒化是枯水的表型之一,一般在甜橙和宽皮柑橘中称作“枯水”,而在柚中称为“粒化”。枯水的果实出现含水量减少、果实质量明显减轻、果实“发泡”等特征,初期果肉表现出汁胞变硬、变干、粒化且颜色变浅,囊瓣增厚等现象,后期严重时果汁减少、糖酸物质丧失、营养物质流失、果实风味寡淡,甚至失去食用价值和商品价值[1-2]。
柑橘枯水于1934年首次由Barthofomew等报道。其后,一些柑橘生产大国如澳大利亚、巴西、中国、印度、以色列、日本等的研究者也作了相关报道[3-8]。世界上几乎所有柑橘种类和栽培品种果实都会在一定程度上出现枯水现象,甜橙类品种如‘凤梨橙’‘华盛顿脐橙’‘哈姆林甜橙’‘血橙’‘伦晚脐橙’等以及宽皮柑橘如‘丹西橘’等,发生果实枯水严重一些,而其他柑橘种类如‘葡萄柚’‘酸柚’‘柠檬’‘莱檬’‘探戈橘’等发生较轻[5,7]。目前柑橘果实枯水发生的生理和分子机制仍不清楚,有效防治措施也尚未解决。本文从细胞学、品质营养、激素水平、酶活性和基因表达等方面对柑橘果实枯水发生的生理变化和分子机制进行综述,旨在为后期进一步深入开展枯水机制和防控措施的研究奠定基础。
1 柑橘果实发生枯水的细胞形态学变化
正常的柑橘果实果皮与果肉通过囊瓣膜(囊衣)紧连,果心间隙小,囊瓣密合,汁胞汁液充足;而枯水后果皮和果肉之间、囊瓣之间以及汁胞形态发生了明显变化,果皮发育失衡,白皮层内侧细胞停止生长,而外侧的细胞则继续生长,进而导致白皮层组织疏松,空隙增大增多,果皮和果肉组织分离[9]。果实枯水初期,细胞组织结构松散;随枯水程度增加,细胞壁增厚,细胞数量增多;在果实枯水后期,汁胞营养物质大量消耗,内含物不断减少,细胞原生质收缩,薄壁细胞崩塌,出现较大空隙[10]。柑橘果实在贮藏过程中,果皮细胞角质层和蜡质层发育不良或果皮细胞组织疏松,引起果皮组织衰老,呼吸代谢加强,果实失水加快,发生采后果实枯水[11-13]。
刘芳等[14]通过对温州蜜柑进行解剖学研究,认为粗皮大果易发生枯水,是由于粗皮果果皮凹凸不平、果皮细胞组织排列疏松、细胞层数增长快,导致果皮变厚。枯水果中果皮细胞壁有较大的细胞间隙,胞间层发生了明显的解体,从而导致细胞间连接松动和细胞壁肿胀,细胞壁的纤维物质发生了明显分离,细胞间隙变大[15]。Burdon等[16]对温州蜜柑浮皮枯水果或正常果皮的磁振显影技术图像分析表明,枯水果实中果皮与囊瓣之间存在较大的间隙,间隙面积约占果实总横切面积的5%。王少峰[17]通过电镜观察发现,芦柑枯水果的果皮油胞层细胞增大,同时细胞内的液泡增大,引起细胞质总量的减少,从而造成果皮发泡。温明霞等[18]采用石蜡切片方法观察温州蜜柑贮藏期间枯水果,发现其细胞排列不规则、结构松散和紊乱,细胞壁增厚,严重时原生质收缩,薄壁细胞崩溃,砂囊中汁胞与汁胞之间出现了一些较大的空隙。甜橙类果实枯水主要发生在中心柱或者果皮附近的汁胞中,汁胞萎缩失水,密度下降,导致中心柱空隙增大,但是果皮无明显变化,很难从外观上进行判断[19]。Wu等[20]通过组织切片观察,发现伦晚脐橙枯水后果实中心柱附近的汁胞皱缩,汁胞间出现空隙,且随着枯水程度的加深,汁胞间的空隙也增大,汁胞空瘪、颜色变浅;Wu等[20]还通过Periodic Acid-Schiff染色发现伦晚脐橙枯水果实的细胞壁纤维素和木质素增多、细胞壁增厚,细胞内含物减少。柚类果实发生枯水时,汁胞膨大、变硬、木质化,果汁显著减少,后期汁胞严重枯水、干瘪,通过扫描电镜观察发现汁胞粒化症状从顶部开始出现,汁胞表面横纹逐渐消失;潘东明等[21]通过透射电镜观察发现,粒化汁胞细胞壁加厚明显,纤维素、木质素、半纤维素类物质等填充细胞壁中胶层,细胞内可见线粒体和高尔基体,同时观察到多个核仁,严重粒化时可见原生质收缩、质壁分离等现象。在果实发育成熟后期和贮藏过程中观察到,柚类果实汁胞粒化发展过程经历正常、凝胶化、粒化及萎缩阶段,粒化的原因是大多数细胞的细胞壁木质化加厚形成厚壁组织[22]。
2 柑橘果实枯水产生的生理生化变化
2.1 果实水分含量变化
柑橘正常成熟果中水分含量约为85%~90%[23],发生枯水时果实中水分严重散失、果实质量减轻、果肉营养物质大量消耗,食用时呈现粗糙、干枯、不化渣等口感[9]。柑橘果实汁胞发生枯水时,水分束缚在果胶中,在Ca2+的参与下形成凝胶,导致自由水含量显著减少,果汁率降低[24]。柑橘果实囊瓣之间、果皮和果肉之间水分相互流通,在贮藏期或受逆境胁迫发生枯水时,水分从果肉流向果皮转移加快,以维持果皮生理活性,水分胁迫诱导了相关激素信号、基因表达、转录因子调控以及相关生理生化反应[25]。果皮中的水分会进一步散逸到空气中,且水分参与的一些生化反应会生成难溶性物质,从而引起水分含量下降[26-27]。
2.2 果实营养物质代谢变化
柑橘果实中含有丰富的营养物质,包括碳水化合物、有机酸、维生素、矿质营养、抗氧化物质等,其中糖酸是柑橘的核心品质,果实成熟时其含量和类型决定柑橘果实的风味[28]。糖酸主要指可溶性糖和有机酸,可溶性糖主要包括蔗糖、葡萄糖及果糖;有机酸以柠檬酸为主,占总含量的70%~90%;还含有少量苹果酸、奎宁酸、琥珀酸、乌头酸、延胡索酸等[29-30]。在糖酸代谢途径中,柑橘糖主要通过蔗糖代谢途径中几种关键酶如蔗糖合酶、蔗糖磷酸合成酶和转化酶降解[31]。蔗糖的降解首先转化为己糖,然后作为呼吸底物通过糖酵解途径进一步合成其他碳水化合物[32-33]。柑橘果实中的主要有机酸是柠檬酸,在果实发育过程中不断增加,而在成熟期和采后过程中逐渐减少[25,34-35]。柠檬酸的减少很大程度上是由于它被用于呼吸底物或合成氨基酸[32,36]。柠檬酸的降解由一系列酶进行催化,例如乌头酸酶(ACO)、磷酸烯醇丙酮酸羧激酶(PEPCK)和异柠檬酸脱氢酶(IDH)[37-38],经由三羧酸循环(TCA)、γ-氨基丁酸(GABA)、谷氨酰胺合成、乙酰辅酶A合成等多个降解途径[34-35,37]。
柑橘果实枯水与糖酸物质代谢直接相关。枯水加速了包括柚、脐橙和椪柑等[39-41]柑橘果实中糖酸的降解。柑橘枯水不仅导致果实中可溶性固形物含量急剧减少,不溶性蛋白质、中性糖(包括鼠李糖、甘露糖、阿拉伯糖和半乳糖等)和可溶性糖(包括葡萄糖、果糖和蔗糖等)含量也降低[41]。在柑橘枯水果实中淀粉或水溶性碳水化合物(可溶性固形物和单糖)转化为聚合多糖等不溶性物质,细胞壁合成加快导致细胞壁加厚[42]。果实汁胞的枯水与可滴定酸呈负相关关系[43-44],甜橙酸含量下降和果汁pH值升高会引起果实枯水的发生,在发生枯水的汁胞中,抗坏血酸的含量出现先下降后增加的趋势[42]。柚类品种在贮藏过程中果实主要品质可溶性固形物、可滴定酸等呈下降趋势,粒化指数增加,含水量显著下降[45]。琯溪蜜柚果实粒化汁胞柠檬酸和异柠檬酸含量较正常果实低,其果汁总酸含量也随之降低,但发生粒化的柚其果汁中苹果酸含量较正常果汁含量高,通过测定发现有机酸代谢相关酶在转录水平上产生了调节,顺乌头酸酶活性的增强导致了柠檬酸的降解,磷酸烯醇丙酮酸羧化酶(PEPC)诱导了苹果酸的生化合成[40]。不同柑橘种类,包括宽皮橘、脐橙、柚等,当果实出现枯水时,糖酸营养物质降解较快,可溶性糖和有机酸的含量均显著下降,严重时降低至正常汁胞的25%~30%[29]。因此,柑橘果实枯水时,糖酸降解导致果实的营养风味发生劣变。还有研究表明,与正常果相比,在一些宽皮柑橘品种中检测到枯水果的呼吸速率增加,呼吸速率加强导致了枯水果营养物质大量消耗[46-47]。对葡萄柚粒化的研究中发现,呼吸速率增强为细胞壁生物合成、糖酸底物降解提供能量[48-49]。在柑橘采后贮藏过程中还观察到果皮的再生长[50],进一步合理解释了果肉发生枯水的原因[51]。
柑橘果实发生枯水时,果汁颜色变淡,汁胞变白,其中类胡萝卜素含量显著下降[52],抗坏血酸、多酚等营养物质含量也显著降低,最高下降50%左右[2]。香气物质是形成果实风味品质的重要因素,主要以萜烯类物质如单萜烯类、倍半萜烯类为主[53],还有酯类、醇类以及醛酮类等挥发性物质[54-55],枯水导致柑橘香气物质尤其萜烯类物质D-柠檬烯大量减少[56],这些物质的大量减少,导致柑橘感官品质中香气丧失,大大降低了果实的风味品质。
2.3 果实矿质营养状况变化
有较多的研究表明,柑橘果实枯水与植株自身营养状况有关。谢志南等[57]发现在琯溪蜜柚果实粒化汁胞中矿质营养元素尤其是N、P、K等大量元素显著增加,而叶片中Ca元素含量减少,并且随着粒化程度增加,土壤中有效Cu含量增加。在甜橙中发现,枯水果实枯水率越高,叶片和果肉中Ca、Zn、B含量越低,但叶片中N、K、Mg、S、Cu、Mn元素含量无显著性变化[42]。Munshi 等[58]研究发现,随着2个甜橙品种果实枯水的发生,叶片N、P、K、Mg、Zn、Cu等营养元素含量增加,而Ca、Fe、B含量减少。‘丹西橘’果皮中K、Mg及果肉中N、P、K、Ca含量水平增加与果实较高枯水发生率相关[59]。Kotsias[60]研究发现果实在枯水过程中,光合产物的转运受到了限制,但增加K和一些微量元素如B、Zn、Cu等有助于营养物质的转运和减少枯水的发生。Wang等[40]在琯溪蜜柚果实粒化发生研究中也发现粒化与植株矿质营养存在相关性,枯水汁胞中检测到矿质元素尤其是P、Mg、S、Zn、Cu等的积累。然而,Singh等[61]发现在‘金诺橘’中果实枯水与植株N、K、Mg、Zn、Cu、Fe含量没有明显关系,可能与柑橘不同品种在逆境胁迫(枯水)下对矿质元素吸收和利用存在差异有关。
2.4 果实酶活性变化
1)细胞壁代谢与水解酶类。柑橘果实枯水与汁胞次生壁形成和细胞壁加厚密切相关[47,62],细胞壁结构的变化主要是由于细胞壁组分包括木质素、纤维素、半纤维素和果胶含量的增加[27,61,63]。
佘文琴等[64]研究发现,红肉蜜柚果实粒化与细胞壁组分和水解酶活性密切相关。在粒化发生期间,汁胞内果胶甲酯酶(PME)、多聚半乳糖醛酸酶(PG)、纤维素酶(CL)等细胞壁降解酶活性较正常果低,而原果胶、纤维素和半纤维素等细胞壁组分物质含量较正常果高。同样,在发生枯水的脐橙突变体中,果实汁胞细胞壁组分物质如原果胶、纤维素、半纤维素以及木质素在整个发育过程中具有较高的含量[65]。在晚熟杂柑黄果柑枯水研究中发现,果实枯水使果皮细胞膜透性增大,细胞壁中木质素、纤维素含量增加,内源多胺物质精胺(Spd)、亚精胺(Spm)合成减少,腐胺(Put)含量降低,(Spd+Spm)/Put比值降低,枯水果抗逆性下降,同时果实中可溶性固形物、Vc、糖以及可食率降低,品质下降[66]。在椪柑果实采后枯水研究中,伴随椪柑果实枯水逐渐发展,参与糖酵解途径、柠檬酸降解途径以及果胶、纤维素、木质素等细胞壁组分的合成途径的相关酶基因表达增加。与此同时,枯水汁胞中参与细胞壁合成途径的酶活性增强,从而增加了纤维素和木质素含量[28]。
正常柑橘果实成熟时,细胞壁的果胶物质在果胶甲基酯酶(PME)的作用下去甲基化和脱酯化,随后被多聚半乳糖醛酸酶(PG)分解成可溶性果胶,细胞壁发生降解从而导致果实软化。由于柑橘果实枯水是一个复杂的生理病变过程,在果实成熟过程中,枯水果实的果胶代谢关键水解酶活性变化也存在差异。例如红桔果实发生枯水时,果皮果胶甲基酯酶(PME)活性升高,原果胶降解加快,果皮组织细胞结构变疏松[67]。特早熟温州蜜柑品种‘胁山’在采前果实浮皮枯水发生过程中,果皮和果肉中PG活性升高,果皮丙二醛(MDA)不断累积,细胞发生膜脂过氧化,从而引起果皮早衰[68]。Sharma等[2]对杂交宽皮柑橘‘金诺橘’果实枯水进行研究,发现枯水果实果胶甲基酯酶(PME)活性较正常果高。谭兴杰等[69]对椪柑采后枯水进行研究,发现果实细胞壁代谢相关的果胶甲酯酶和多聚半乳糖醛酸酶出现2个活性高峰,在高峰期间果实质量减轻,高峰过后,果实出现枯水症状。
2)自由基代谢酶类。正常情况下植物细胞内的活性氧处于一种累积与清除的动态平衡过程,一旦这种平衡被破坏将导致细胞内的生理紊乱,从而引起植物生理病变。Sharma 等[7]研究发现尽管多酚氧化酶(PPO)活性在不同柑橘品种中存在差异,但枯水果实中PPO的活性均低于正常果实。柚类不同品种采后贮藏期果皮和果肉的过氧化物酶(POD)和超氧化物歧化酶(SOD)活性变化存在差异,果皮POD先升后降,SOD先降后升,而果肉POD活性较低且变化无规律,SOD先升后降,呈与果皮相反的趋势;并且随着果皮中SOD活性的增加和果肉中SOD活性的下降,果实开始出现粒化[70]。琯溪蜜柚在果实成熟后期,活性氧H2O2代谢失调,在细胞中过量累积,膜脂过氧化;同时H2O2活性氧不断积累诱导相关清除酶类SOD、POD、CAT(过氧化氢酶)活性不断增强,促进细胞内木质素合成,从而果实发生粒化[71]。胡位荣等[52]选取红肉蜜柚正常汁胞、半枯汁胞、全枯汁胞为材料对比分析汁胞枯水氧化酶活性,发现随粒化发生和发展,汁胞中SOD活性显著下降,而POD活性显著上升,CAT活性先上升后下降,PPO氧化酶活性也显著降低,积累较多的活性氧H2O2,引起自由基代谢失调,细胞内丙二醛(MDA)含量增加,膜脂过氧化,且木质素不断累积,汁胞硬化、粗糙,呈现粒化较为典型的症状。
2.5 果实激素含量变化
植物激素和生长调节剂在柑橘果实发育和采后贮藏中起重要调控作用,与果实枯水发生存在相关性。在枯水过程中,果实内源激素含量会相应发生改变。椪柑果实在贮藏过程中,枯水果的果皮中脱落酸(ABA)含量较正常果高,而IAA和CTK含量则低于正常果,且果肉ABA含量在枯水过程中显著上升[72]。ABA的增加和积累加快了柑橘果实次生物质如木质素、纤维素以及果胶等的代谢,导致糖酸含量下降,细胞壁变厚,呈现枯水症状。自交琯溪蜜柚汁胞粒化比较严重,而通过杂交授粉的琯溪蜜柚其果实汁胞粒化较轻。前人研究发现,杂交果中含有较高含量的内源激素GA3,授粉后可使子房IAA、ABA等内源激素含量增加,促进果实发育,从而降低果实粒化的发生率[73]。另外,外源施加植物激素也能调控柑橘果实枯水的发生。例如,Inder等[74]通过外源施用GA、2,4-D、2,4,5-T和NAA 显著降低了‘丹西’红桔枯水的发生率和发生程度,并增加了单果质量、总糖和可溶性固形物。Shi等[75]通过外源激素持续处理离体培养的柚汁胞50 d,发现外源IAA和ABA可以显著抑制或者促进CgMYB58及其靶基因CgPAL1、CgPAL2、Cg4CL1和CgC3H的表达量,从而有效调控华农红柚汁胞粒化的发生。
3 柑橘果实枯水的分子生物学研究进展
近年来,随着分子生物学技术的发展,柑橘枯水的研究不再停留于形态、生理生化和细胞层面,转而从分子层面和基因层面对柑橘果实枯水的机制进行研究。佘文琴等[71]采用mRNA 差异显示技术对琯溪蜜柚汁胞响应粒化的基因进行鉴定,获得了25 个差异cDNA 片段,并进一步采用RACE技术克隆了相关基因cDNA全长。钟凤林[76]采用双向电泳技术和质谱技术对琯溪蜜柚汁胞不同发育时期响应粒化的蛋白质进行分析,鉴定到30个在粒化汁胞中差异表达的蛋白,包括与细胞壁伸展相关的Germin(草酸氧化酶)蛋白和汁胞发育相关的APX(抗坏血酸过氧化物酶)蛋白。杨明等[77]采用抑制性差减杂交(SSH)技术构建了椪柑果肉枯水差减文库,分离到52个椪柑枯水相关差异表达基因,其中8个参与衰老、抗逆防御、几丁质和细胞壁代谢、蛋白降解的基因均显著上调表达。Zhang等[63]利用生物信息学分析筛选到12个与甜橙枯水相关的候选miRNA,并且发现csi-miR397,N-miR828 可能通过分别调控Cs6g06890.1和Cs1g17590.1基因的表达来调控木质素的合成从而调控柑橘汁胞枯水。Yao等[29]采用转录组测序技术研究椪柑果实采后枯水机制,发现细胞壁果胶合成关键基因 UDP-葡萄糖醛酸-差向异构酶基因(GAE)和半乳糖醛酸转移酶基因(GAUT)表达量显著上调,而果胶降解关键基因果胶甲酯酶基因(PME)表达量显著下调,导致枯水汁胞中果胶积累。纤维素合酶基因和木质素合成基因如肉桂醇脱氢酶(CAD)基因表达量显著上调,导致细胞壁纤维素和木质素含量增加。Wu等[20]通过转录组测序从晚熟脐橙采前枯水果实中鉴定出903个差异表达基因,并且发现与果胶分解相关的PME基因上调表达,多聚半乳糖醛酸酶(PG)基因则下调表达;与木质素合成相关的苯丙氨酸酶(PAL)、CAD和POD上调表达,而参与纤维素分解的CL下调表达。Shi等[75]对‘华农红柚’‘HB柚’和‘高班柚’不同发育时期果实进行分析,发现‘华农红柚’最易发生枯水,同时其木质素含量也最高;随后对‘华农红柚’‘HB柚’和‘高班柚’不同发育时期汁胞进行转录组分析发现PAL、4-香豆酸酯CoA连接酶基因(4CL)、肉桂酸4-羟化酶基因(C4H)、咖啡酰CoA 3-O-甲基转移酶基因(CCoAOMT)等17个木质素合成相关基因在易枯水的华农红柚汁胞中高表达,说明木质素合成途径可能是影响柑橘果实枯水的关键因素。
转录因子在植物次生细胞壁生物合成和发育中起着关键的作用[78-79]。Yao等[29]在椪柑枯水汁胞中鉴定到一些响应枯水的转录因子如NAC、MYB等。前人研究表明,柑橘CsMYB330、CsMYB3308和CsMYB85可以精细调控木质素生物合成相关基因表达,进而调控次生细胞壁合成[80-81]。Shi等[75]发现将CgMYB58瞬时超量表达‘华农红柚’和‘高班柚’的白皮层或稳定超表达柑橘愈伤组织能够激活木质素合成关键基因CgPAL1、CgPAL2、Cg4CL1和CgC3H的表达,可以显著增加木质素的含量,说明MYB58能够通过调控木质素的合成而直接影响柚果实的粒化。
4 柑橘果实枯水发生的生理及分子机制
柑橘果实长期储存或受冬季低温胁迫的影响,导致汁胞中果胶分解相关的多聚半乳糖醛酸酶(PG)基因下调表达,PG酶活性较低,而果胶甲基酯酶(PME)基因显著上调表达,果胶被PME酶去甲基化和脱酯化,溶解性不断增加,但由于PG酶的低活性可能不足以完全解离胶质侧链并顺利分解果胶,从而导致高分子质量果胶的积累并增加了果汁的粘度。随后果胶物质与游离的Ca2+结合导致果实汁胞硬化[82]。当可溶性果胶积累达到一定水平时,可在Ca2+的作用下形成钙桥或被酯键固定化,从而形成凝胶,将水分束缚在凝胶中。同时,汁胞中纤维素酶(CL)基因的表达显著下调,纤维素酶活性低,降解过程受阻,导致纤维素含量的积累。此外,转录因子MYB58的表达上调,导致汁胞中PAL、4CL、CAD和POD等与木质素合成有关的基因显著上调表达,相应的酶活性升高,木质素大量合成。随着木质素合成的增加,分解速率低于合成速率,导致次生壁生长、增厚,呈现出枯水症状。上述过程同时受到植物细胞内源激素尤其是脱落酸(ABA)和生长素(IAA)的调控,ABA在发生枯水过程中一直保持着高水平,而IAA则保持较低水平。ABA和IAA能够显著抑制或者促进CgMYB58及其靶基因CgPAL1、CgPAL2、Cg4CL1和CgC3H的表达量,从而导致果实枯水的发生(图1)。
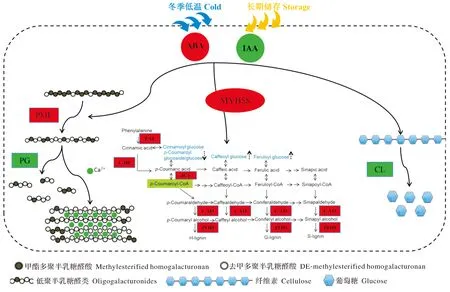
ABA:脱落酸,IAA:吲哚-3-乙酸,PME:果胶甲基酯酶,PG:多聚半乳糖醛酸酶,PAL:苯丙氨酸解氨酶,C4H:肉桂酸4-羟化酶,4CL:4-香豆酸酯CoA连接酶,CAD:肉桂醇脱氢酶,POD:过氧化物酶,CL:纤维素酶。红色表示在枯水的柑橘果实中上调,绿色表示下调。ABA:Abscisic acid,IAA:Indole-3-acetic acid,PME:Pectin methylesterase,PG:Polygalacturonase,PAL:Phenylalanine ammonia lyase,C4H:Cinnamate 4-hydroxylase,4CL:4-Coumarate CoA ligase,CAD:Ainnamyl alcohol dehydrogenase,POD:Peroxidase,CL:Cellulase. Red colour indicated up-regulation in granulated citrus fruits,green colour indicated down-regulation.
5 展 望
柑橘果实汁胞受内源激素、相关酶和基因的调控,发生糖酸物质代谢、营养物质消耗等一系列生理生化变化,汁胞水分和内含物质的不断转移和消耗,最终导致果实汁胞崩塌、变瘪、变干和枯水。近年来研究人员从细胞学、品质营养、激素水平、酶活性和分子生物学等方面对柑橘果实枯水发生的生理和分子机制进行了初步的探究,并在柚类果实粒化的转录调控机制研究中取得了突破性的进展。但关于柑橘果实枯水过程中代谢产物的变化和基因的生物学功能还有待深入研究。今后对柑橘枯水机制和防控的研究应集中于以下几个方面:(1)通过靶向/非靶向LC-MS对柑橘枯水果实中的代谢物进行检测,并结合转录组明确影响柑橘枯水的关键代谢物及其在转录水平的调控;(2)通过酵母文库深入挖掘调控柑橘果实枯水的关键转录因子,并通过柑橘遗传转化研究细胞壁代谢关键基因(PG、PME、PAL、POD、PAL、4CL等)及关键转录因子的生物学功能;(3)基于柑橘枯水生理生化和分子机制研究成果,在生产中开展柑橘枯水的防控技术措施研究,包括柑橘新品种区域化布局、柑橘园温湿度等环境调控、砧穗组合筛选、矿质营养和生长调节剂的应用、留树和采后保鲜药剂筛选等,从而预防和减轻柑橘果实枯水。
猜你喜欢
文萃报·周五版(2021年44期)2021-12-06 20:05:14
耕作与栽培(2021年3期)2021-12-01 02:38:13
食品与发酵工业(2020年18期)2020-01-06 22:43:12
广西农学报(2019年4期)2019-11-26 11:56:23
浙江工业大学学报(2017年5期)2018-01-22 02:03:42
广东农业科学(2017年5期)2017-08-29 10:37:54
长江科学院院报(2014年12期)2014-08-20 08:50:06
温州职业技术学院学报(2014年3期)2014-03-11 19:03:38
食品工业科技(2014年9期)2014-03-11 18:15:40
华东师范大学学报(自然科学版)(2014年3期)2014-03-11 16:18:22
