
瘤胃微生物调控牛奶品质的研究进展
2021-01-29 06:17刘凯珍张养东黄国欣王加启
饲料工业 2021年1期
刘凯珍 张养东 黄国欣 郑 楠 王加启
(中国农业科学院北京畜牧兽医研究所,动物营养学国家重点实验室,北京100193)
牛奶是人类理想的食物之一,因为其不仅富有全面的营养成分,而且含有多种活性物质[1-2]。社会日益进步,人类生活水平不断提高,以及近年来二胎政策的开放使得人们对牛奶的需求量随之增加,尤其是对优质乳源的需求更加急切。因此,提升牛奶品质是全球奶业科研工作者的重要任务之一。
以往多项研究表明,在奶牛将日粮转化为牛奶的过程中,多种因素导致牛奶质量层次不齐,如奶牛基因、饲养环境和饲喂日粮等[3-7]。目前,研究最多的是通过调控日粮,从而改变牛奶营养成分的含量,虽然在一定程度上提升了乳蛋白、乳脂肪或功能性脂肪酸,然而这种途径常常顾此失彼,增加了生产成本或者是以牺牲其他生产性能为代价[8-9]。
近年来,有研究发现遗传背景、饲喂管理和饲养环境均一致的情况下,牛奶营养成分不同的奶牛其瘤胃微生物存在明显的差异,这表明动物机体将饲料中的养分转化为牛奶营养成分的过程中,微生物扮演着重要的角色,主要在乳蛋白和乳脂肪前体物的形成过程中发挥作用[10-11]。因此,瘤胃微生物也许是调控牛奶品质的一条有价值的途径。
1 牛奶营养成分的形成及其调控
奶牛能够将饲草转变为牛奶,这主要依赖于其独特的消化器官——瘤胃[12]。日粮被食入瘤胃以后,含氮物质在微生物的作用下被分解为肽和氨基酸,其中一部分又被特定的微生物利用,合成微生物蛋白(MCP),过瘤胃蛋白(UDP)和MCP一起进入小肠中被降解为氨基酸,经血液运输至乳腺,用于合成乳蛋白。以往对于牛奶中乳蛋白合成的研究大多集中于乳腺上[13],鲜有从瘤胃微生物利用含氮物质的效率层面研究乳蛋白形成的报道。
瘤胃中纤维分解菌能够将食入的粗纤维分解为挥发性脂肪酸(VFA,其中浓度最高的是乙酸、丙酸和丁酸),丁酸以β-羟丁酸的形式和乙酸在乳腺中用于合成乳脂防酸,主要是C4~C14脂肪酸;另外饲料中包含的脂类物质在微生物作用下首先分解为甘油和脂肪酸,然后C16以上的长链脂肪酸以非脂化脂肪酸的形式被血液运输至乳腺。脂肪酸、甘油、葡萄糖在乳腺中合成三酰甘油,构成乳脂。
葡萄糖是乳糖合成的重要前体物质,在二磷酸尿嘧啶葡萄糖异构酶的作用下一分子葡萄糖异构为一分子半乳糖,然后在乳糖合成酶的作用下一分子半乳糖和一分子葡萄糖经β-1,4 糖苷键连接形成乳糖[12]。乳腺中的葡萄糖主要来源于瘤胃中淀粉的分解,以及纤维分解产生的丙酸经体内三羧酸循环转化而成[14](图1)。乳蛋白、乳糖和乳脂是牛奶总固形物的主要组成成分,并且瘤胃微生物在这些前体物的形成中发挥了关键作用,因此瘤胃微生物对于牛奶品质也有着重要的影响[15-16]。
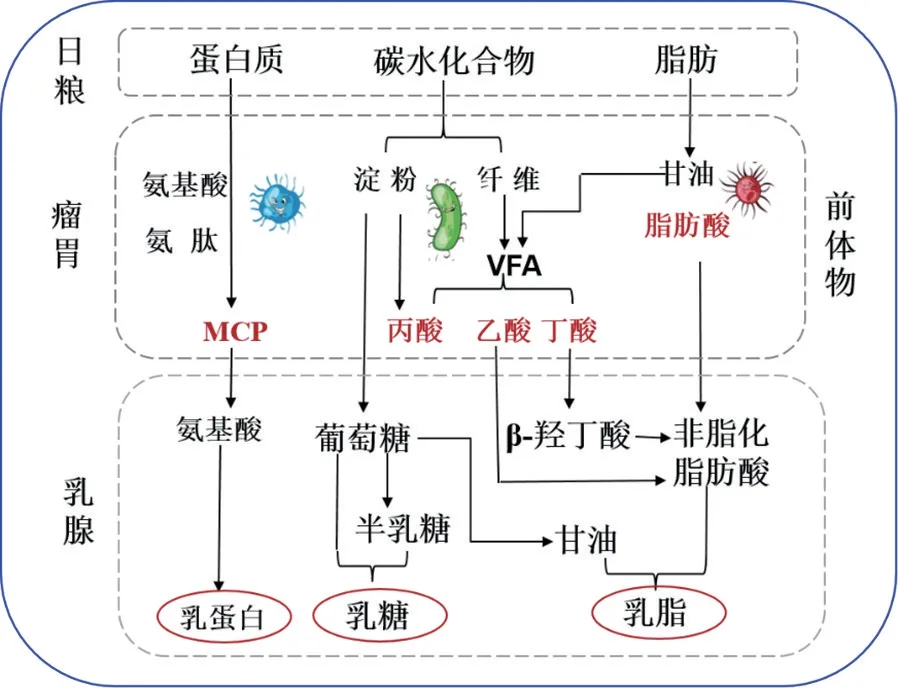
图1 乳蛋白、乳脂和乳糖在体内形成的主要过程
2 影响牛奶品质的因素
牛奶中含有丰富且均衡的营养物质,而且含有多种功能性脂肪酸和活性物质,如二十二碳五烯酸(EPA)、二十二碳六烯酸(DHA)、乳铁蛋白、生长因子等。针对牛奶品质的调控,以往集中研究了提高乳蛋白和乳脂肪产量的方法,以及开发功能性牛奶,如富含ω-3多不饱和脂肪酸的牛奶。研究发现,牛奶中易受影响的成分主要是乳蛋白和乳脂肪,乳糖含量的变化范围较小[8]。
2.1 遗传因素
遗传因素是决定奶牛产奶量、乳蛋白和乳脂产量,及其合成效率的内在因素,不同物种和不同品种奶牛的遗传性能存在较大的差异。荷斯坦奶牛的产奶量高于娟珊牛,但是牛奶总固形物含量和乳脂率均低于娟珊牛[17]。荷斯坦奶牛和瑞士褐牛的杂交奶牛,乳脂和乳蛋白产量高于纯种荷斯坦奶牛[3]。另外,有研究还发现奶牛的乳蛋白率与繁殖性能的遗传因素呈正相关[18]。Pegolo 等[13]针对1 011 头瑞士褐牛进行了全基因组相关研究(GWAS),关联分析结果发现GFI1B、ZNF407 和NR5A1 可能是乳蛋白合成的主要调节因子。多项研究结果显示,培育乳蛋白和乳脂肪产量高的奶牛是提高牛奶品质的一种有效手段[4]。但是由于奶牛的繁殖周期长达一年、单胎产犊只有一个等特点,给育种工作带来了很大的难题,因此通过育种改善牛奶品质是一个长期的过程。
2.2 日粮因素
多年研究结果和生产实践显示,乳糖产量受外界因素的影响较小,因此大多数科研工作者将提升牛奶品质的重点工作聚焦在提高乳蛋白和乳脂产量,以及牛奶中功能性脂肪酸(EPA、DHA、CLA)和活性物质的含量上[8,19]。
日粮中适当比例的粗饲料是保证乳脂率的关键,因为粗饲料的发酵产物乙酸和丁酸是乳腺合成乳脂的重要前体物质[20]。当精料比例过高时,瘤胃中丙酸产量增多,发酵模式发生改变,导致乳脂率下降[21-22]。然而不同奶牛的耐受性存在很大差异,饲喂高精粮日粮(粗料∶精料=35∶65)的泌乳中期荷斯坦奶牛,乳脂率均有所下降,但是耐瘤胃酸中毒的个体下降程度较小[23]。研究发现,日粮中适当补充脂肪或缓冲剂能够防止乳脂降低[23-24]。并且日粮中添加脂肪能够在一定程度上影响脂肪酸的组成,进而影响牛奶中功能性物质的含量,日粮中添加粗甘油,乳脂中奇数链脂肪酸和共轭亚油酸有增加的趋势[9],日粮中添加牧草和植物油料种子以及微藻,增加了牛奶中ω-3多不饱和脂肪酸和顺式-9,反式-11 共轭亚油酸,降低了饱和脂肪酸的含量[19]。
以往的研究发现,日粮中补充粗饲料对乳蛋白的影响并不大[24],因为乳腺合成乳蛋白的原料主要是氨基酸,而蛋白在瘤胃内的转化过程较复杂。许多学者通过添加过瘤胃保护氨基酸、胆碱等营养物质来提高奶牛的乳蛋白产量[25]。另外,日粮中补充叶酸,可提高代谢效率、泌乳性能和牛奶的营养物质含量,特别是在泌乳早期日粮中添加[26]。综上,通过改善日粮以提高牛奶品质是最有效的途径,然而也存在诸多弊端,因为在提高养分含量的同时提高了饲养成本,并且可能降低饲料利用效率。
2.3 环境因素
对奶牛的产奶量和乳成分影响较大的环境因素主要是温度和湿度[5-6]。Hill 等[5]分析了苏格兰地区1 369 头奶牛21 年来的温湿度指数(THI)、奶产量和乳成分等数据,结果发现在室内和室外饲养的奶牛,蛋白质含量均随着THI的增加而降低,并且在室外饲养比室内饲养降低的速度更大。THI 的改变不仅受饲养环境的影响,另外一个最主要的原因是随季节的改变而变动。研究发现乳脂肪、乳蛋白、总固形物和非脂固形物的含量在夏季最低,而在冬季达到最高[6]。造成这种现象的原因可能是由于夏季温湿度较高,容易造成热应激,而机体发生热应激时产奶性能将会受到严重的影响[27]。虽然环境对于牛奶品质有一定影响,但是随着生产水平的提高,养殖环境越来越完善,各项养殖设施也得到了优化,大大降低了环境造成的经济损失。
除日粮、遗传和环境因素外,机体的生理因素,如奶牛的胎次、泌乳阶段和疾病等[28-29],生产上的管理因素,如挤奶频率和饲喂方式等,也会对牛奶的品质产生一定的影响[25]。综上所述,任何因素发生变化都会在一定程度上影响机体对养分的吸收和利用,并且影响转化到牛奶中的养分,进而改变牛奶品质。
3 瘤胃微生物与牛奶品质
奶牛瘤胃中栖息着大量的微生物,如细菌、原虫、古生菌和真菌[30],在分解饲料和利用养分的过程中行使着各自的功能。奶牛体内约70%能量和50%蛋白质通过瘤胃微生物发酵提供[31]。牛奶的重要营养成分,如乳蛋白、乳脂肪和乳糖的前体物质大部分都是在瘤胃微生物的作用下产成的,而前体物的产量直接影响牛奶相应的养分含量。因此,瘤胃微生物与牛奶品质有着密不可分的关系。曾有研究发现,瘤胃微生物与奶牛的生产性状(如饲料利用效率、产奶量和乳成分)之间存在着紧密的相关性[10,32]。产奶量是衡量奶牛生产性能的重要指标,直接影响各种乳成分的产量,有研究发现,产奶量低的奶牛,其瘤胃乳酸菌如Streptococcus、unclassifiedEnterobacteriaceae、Ruminobacter、Treponema和unclassifiedBacteroidaceae含量较高[27]。
3.1 瘤胃微生物对乳蛋白的影响
乳蛋白的前体物主要是氨基酸,而氨基酸很大部分来源于瘤胃中合成的MCP 和直接进入小肠的UDP。UDP 的含量主要取决于饲料的质量[33],而MCP的产量与瘤胃微生物的结构和组成,以及原虫数量有直接关系[34]。有学者提出瘤胃中主要的蛋白降解菌有Bacteroides、Butyrivibrio、Clostridium、Eubacterium、Streptococcus和Selenomonas[35-36]。另外,瘤胃原虫,如纤毛虫能够吞噬细菌以满足自身的营养需要,因此原虫的存在降低了饲料的能量利用效率[37-38]。产甲烷古菌同样也能够降低机体对能量的利用,并且产甲烷古菌的丰度高更利于甲烷的产生[8],造成温室效应。瘤胃MCP 产生的过程需要能量的供应,因此原虫和产甲烷古菌能够通过影响供能来影响MCP的产量。
研究发现,饲喂同样日粮的奶牛,乳蛋白产量高(HH)的个体,其瘤胃微生物的丰富度小于乳蛋白产量低(LL)的个体,其中Proteobacteria在HH 组的丰度比LL 组高1.36 倍,属水平上Succinivibrio的相对丰度在HH 组中显著低于LL 组,Clostridium的相对丰度有降低的趋势,而HH 组中Sharpea的相对丰度则是LL奶牛的2.28 倍[11]。在其他奶畜中也发现了相似的现象,然而由于物种的差异,结果可能出现一定的差异。Zou等[39]针对广西乳水牛与乳蛋白和总固形物含量进行相关性分析,结果显示Acinetobacter与乳蛋白平均产量(R=0.56;P=0.03)、乳总固体平均产量(R=0.60;P=0.02)和标准乳产量(R=0.57;P=0.03)呈正相关(见表1)。
3.2 瘤胃微生物对乳脂的影响
牛奶中的乳脂肪来源于优良的饲草饲料,乳脂肪产量取决于奶牛对饲料的转化效率。乙酸、β-羟基丁酸和游离脂肪酸是主要的乳脂肪前体物,其中乙酸和丁酸主要由瘤胃微生物发酵粗饲料产生,然后经血液转运到乳腺用于合成三酰甘油。在此过程中瘤胃微生物对纤维的分解作用是决定乳脂前体物的关键环节。研究发现,Prevotella 的OTU 与产奶量和乳脂产量呈负相关[10]。水牛瘤胃中丁酸弧菌与乳脂肪的平均产量(R=0.55;P=0.03)、总固形物平均产量(R=0.56;P=0.03)和标准乳产量(R=0.52;P=0.05)呈正相关[39]。奶牛发生低乳综合征时其瘤胃Prevotella 的相对丰度显著增加,而unclassifiedLachnospiraceae、Oribacterium、unclassifiedVeillonellaceae和Pseudobutyrivibrio的相对丰度显著降低,并且乙酸和丁酸浓度均有降低的趋势[40]。研究还发现,瘤胃微生物群对荷斯坦牛乳中奇数链脂肪酸和C18多不饱和脂肪酸含量有显著影响,对短链和中链脂肪酸含量的影响较小[41]。部分瘤胃微生物能够通过生物氢化将不饱和脂肪酸变为饱和脂肪酸,从而改变牛奶脂肪酸的组成[42]。因此,由于瘤胃生物氢化菌的存在,通过日粮中添加不饱和脂肪酸来改善牛奶脂肪酸组成效果不明显。然而也有多项研究发现,unclassifiedClostridiales和Ruminococcaceae对生物氢化有显著影响[40,43]。研究发现饲喂同样日粮,直接添加PropionibacteriumP63能够增加牛奶中十六碳烯酸三酰甘油(cis-9 16∶1)的含量[44](见表1)。
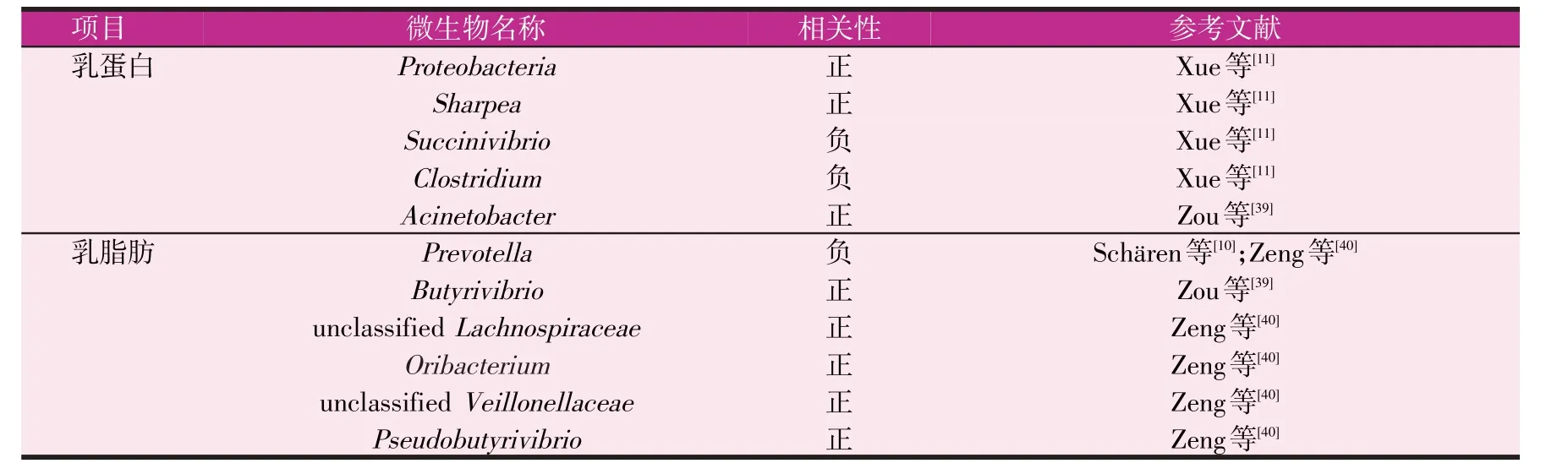
表1 瘤胃微生物与乳脂肪和乳蛋白产量的关系
总之,乳脂产量和乳脂肪酸组成的变化通常是由瘤胃发酵和生物氢化途径的改变引起的,而瘤胃微生物无论在瘤胃发酵还是生物氢化的过程中都是必不可少的,因此调控瘤胃微生物也许是改善乳脂降低综合征和增加牛奶功能性脂肪酸的重要方式。
3.3 瘤胃微生物对功能性物质的影响
牛奶中丰富的活性蛋白,如乳免疫球蛋白G(IgG)、乳铁蛋白(LTF)、溶菌酶(LYS)和乳过氧化物酶(LP)等,通常也受到广泛的关注。改善瘤胃微生物环境能够显著提高牛奶中活性蛋白的含量。研究发现日粮中添加益生菌(Lactobacillus casei和Lactobacillus plantarumP-8)虽然对乳脂、蛋白质、乳糖含量影响无显著性差异(P>0.05),但是能够显著提高产奶量和IgG、LTF、LYS 和LP 的含量,同时降低了体细胞计数(P<0.01)[45]。研究还发现,瘤胃中益生菌与瘤胃发酵菌,如Bacteroides、Roseburia、Ruminococcus、Clostridium、Coprococcus和Dorea等存在正相关关系,而与Bacillus cereus、Cronobacter sakazakii和Alkaliphilus oremlandii等条件致病菌呈负相关关系。
4 结论与展望
不同的研究表明,瘤胃微生物与奶牛的产奶量和乳成分之间存在着紧密的相关性。因此越来越多的学者着眼于通过瘤胃微生物手段改善牛奶的品质。许多学者还试图通过交换瘤胃内容物来改变瘤胃微生物。Weimer 等[46]将高低产奶性能的奶牛瘤胃内容物进行了交换,结果发现低产奶牛瘤胃中乳脂的前体物乙酸和丁酸有所增加,但是1 d 后又恢复到原有水平,并且交换内容物后10 d内恢复到交换前的细菌群落。成年反刍动物瘤胃微生物已经达到稳定状态,具有一定适应变化的能力,因此,通过改变成年动物瘤胃微生物群落以提高产奶性能效果并不理想。但是,幼龄反刍动物的瘤胃微生物具有较强的可塑性,断奶前瘤胃微生物群的初步定植对于后期牛奶的产量和养分的含量有重要的影响[47]。立足当前的研究基础,未来应该重点探索影响牛奶品质的特定微生物,接下来从调控断奶前后的犊牛瘤胃微生物入手,从而达到提高牛奶品质的最终目的。目前,量化瘤胃中特定的微生物所起的作用还比较困难,也是未来需要关注的因素。
在过去的20 年里,新一代测序技术的应用使得人们对瘤胃微生物群落有了新的认识。宏基因组和宏转录组技术为特定瘤胃微生物的功能揭开了神秘的面纱。目前人们了解更多的是,瘤胃微生物对于纤维和非纤维碳水化合物的消化、微生物蛋白质的合成和生物氢化的作用。然而,瘤胃微生物与宿主的相互作用及其在体内的代谢通路和因果关系还不太清楚,值得进一步研究。未来应重点从微生物的特定基因和微生物基因的转录活性入手,探索微生物影响产奶量和乳成分的代谢通路,为提高牛奶品质的技术开发提供理论基础。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
动物营养学报(2022年9期)2022-10-12
中国饲料(2022年5期)2022-04-26
湖南饲料(2021年4期)2021-10-13
湖南饲料(2021年3期)2021-07-28
中国畜牧杂志(2021年1期)2021-01-20
河南畜牧兽医(2020年1期)2020-01-11
中国奶牛(2019年12期)2020-01-08
分析化学(2018年4期)2018-11-02
现代畜牧科技(2018年3期)2018-05-14
