
九万山常绿阔叶林物种组成及空间结构特征分析
2021-01-29 08:18彭玉华申文辉何琴飞欧芷阳
中南林业科技大学学报 2020年12期
彭玉华,曾 健,申文辉,何 峰,郑 威,何琴飞,欧芷阳
(1.广西壮族自治区林业科学研究院,广西 南宁 530002;2.广西九万山国家级自然保护区管理中心,广西 融水 545300)
森林空间结构是树木在林地的分布格局及属性在空间上的排列方式,表达的是树种、林木大小、分布等空间关系,是群落树木之间的竞争势及其空间生态位的决定因素,也在很大程度上对森林林分的稳定性、发展的可能性和经营空间的大小具有决定性影响,对林分空间结构进行定量分析,将有利于提高森林林分经营决策的准确性[1-4]。随着人类对环境与发展问题认识的不断加深,森林经营理念已转变至可持续发展,森林可持续经营对森林精确信息需求的不断增加,使得林分空间结构的研究越发重要[5]。林分空间结构作为森林信息重要部分,是森林空间结构的最直观表现因子,已逐渐成为国内外研究森林结构时的热点[6-13]。以林分相邻木为单元构建林分空间结构的混交度、大小比数、角尺度三个参数,分别描述了林分树种空间隔离程度、林木大小分化程度和林木水平分布格局,提高了林分结构的解析性,能准确地描述林分所处的状态,对创建与维护最佳的森林结构有着重要意义,因此,被学者们广泛应用于描述和研究林分空间结构特征,有较好的参考和运用价值[14-17]。
天然阔叶林林分结构稳定、物种繁多、组成复杂,对保护生物多样性、发挥森林功能和维持区域生态平衡等具有不可替代的作用,对环境保护、维持全球碳平衡和可持续发展等都具有重要作用[15]。广西九万山国家级自然保护区是我国南部常绿阔叶林及其垂直带森林生态系统保存较好的地区之一,是中国亚热带常绿阔叶林中的苗岭山地常绿阔叶林的重要组成部分,在中国中亚热带地区中具有相当高的典型性和代表性,同时也是全球同纬度地区生物多样性保护的关键地区[18]。九万山保护区常绿阔叶林生长茂盛,生物资源异常丰富,特有种类繁多,林分结构组成较为复杂。森林经营的基础是林分结构的研究,通过研究天然林林分空间结构,让人们更清楚地了解森林群落规律和森林结构特征,及其了解优势种群的数量组成和空间分布格局的特征对林分结构所产生的影响,从而培育出健康稳定的森林。本研究基于九万山常绿阔叶林森林动态监测样地为研究平台,开展九万山保护区林分结构研究,分析描述林分数量特征的同时,利用空间结构参数对林分空间结构特征进行全面描述与分析,以期为人们更好地认识和了解该区域植物群落的特征,旨在为九万山自然保护区政策制定提供有效参考,对该区域常绿阔叶林的生物多样性保护和可持续经营管理具有重要意义。
1 研究区概况
广西九万山国家级自然保护区地处广西北部(108°33′32″E—108°48′49″E,25°01′55″N—25°19′54″N),地跨环江、罗城与融水三县,地处广西与贵州交界处,属苗岭山脉南缘。保护区林地面积24 872.7 hm2,其中天然林面积20 721.8 hm2,占林地总面积的83.31%,森林覆盖率 95.7%,活立木蓄积量2.610 46×106m3。山峰海拔普遍在1 000 m 以上,谷底约400 m 左右,最高峰海拔1 693 m;保护区属中亚热带季风气候区,多年平均日照时数1 000~1 600 h;多年平均气温12.0~17.1℃,最冷月(1月)平均气温4~6℃,极端低温-8℃,最热月(7月)平均气温 22~25℃,极端高温37℃;年均降水量1 600~2 100 mm,降水量多集中在4—8月,占全年降水量的70%左右,相对湿度为85%~90%;全年无霜期300 d。土壤为酸性土,肥力较高。保护区属中亚热带常绿阔叶针叶林区,树种以阔叶树和杉木为主;植被类型多,结构复杂,在国内同纬度地区甚为少见。调查样地为典型的中亚热带常绿阔叶林,主要的树种有:刨花润楠Machilus pauhoi、建润楠Machilus oreophila、柳叶润楠Machilus salicina、润楠Machilus pingii、桂南木莲Manglietia chingii、新木姜子Neolitsea aurata、黄丹木姜子Litsea elongata、多花杜鹃Rhododendron cavaleriei、红花荷Rhodoleia championii、木荷Schima superba、硬毛山香圆Turpinia affinis、两广杨桐Adinandra glischroloma、水青冈Fagus longipetiolata等等。
2 研究方法
2.1 样地设置与调查
2017年3月—2018年10月,在广西九万山国家级自然保护区的杨梅坳片区,经全面踏查并结合地形和林分特征基础上,利用全站仪(南方测绘,NTS-372R10)确定样方范围和方向,设置相隔3 km 的2 块面积为100 m×100 m 的永久性标准固定样方,分别标记为样方a、样方b。利用GPS 进行定位,测定样地的经纬度、海拔等立地因子,并进行植被群落学调查。样方a 为108°38′54″E,25°12′58″N,海拔为1 243 m,坡向为东南109°,坡度25°,坡位中上,郁闭度0.9;样方b为108°33′38″E,25°12′17″N,海拔为1 210 m,坡向为西北280°,坡度25°,坡位中上,郁闭度0.9。采用相邻网格法将每个样方划分25 个20 m×20 m正方形小样方网格,以网格为单元,进行乔木层调查,以样地西南角为坐标原点,东西向为横轴(x),南北向为纵轴(y),对样地内胸径≥2 cm 的独立木本植物分别用网格进行全林定位,记录所研究林分每个个体的相对坐标、树种、胸径(DBH)、树高(H)、冠幅等。在处理数据时把相对坐标做全局化处理。
2.2 重要值计算方法
植物种群重要值参照李语晨等[1]计算方法:乔木重要值=(相对频度+相对优势度(胸径断面积)+相对多度)/3。
2.3 林分非空间结构分析
以2 cm(胸径至28 cm 后以4 cm)为标准径阶、2 m 为标准高度分别作直方图。
2.4 空间结构分析
为全面描述并分析九万山国家级自然保护区内单个林木和整个林分的林木空间分布格局、林木大小分化程度(生长优势状态)以及树种的空间隔离程度,本研究采用惠刚盈等[19-20]的森林空间结构量化分析方法分析空间结构特征数量表达的参数(混交度、大小比数、角尺度)。为能明显地表达出混交度等结构参数的林学意义,本研究采用参照树周围4 株相邻木组成一个结构框架,以每株树木(i)为对象,根据该对象与样地4 条边的最近距离以及与周围树木间的距离选取4 株最近相邻木。为了消除处于林分边缘树木的系统影响,设置了5 m 缓冲区,缓冲区内林木只做相邻木处理。
混交度、大小比数、角尺度参照惠刚盈等[19-20]的公式计算。
林分空间结构指数(FSSI):林分空间结构指数能够从整体上来把握林分的空间结构特征[21-22]。其表达式为:

式(1)中:FSSI、M、U、W分别为林分空间结构指数、混交度、大小比数、角尺度,它们的取值范围均为[0,100]。林分空间结构指数越大,林分结构越理想,即当林分空间结构指数最大[FSSI=100,M=100、U=0、W=50]时林分是最理想状态。
林分空间结构距离(FSSD):现实林分的空间结构向理想结构点逼近或者远离的趋势可以通过不同林分条件下林分空间结构点到理想结构点的距离来表示,即用林分空间结构距离(FSSD)来定量化表达[21-22]。其表达式为:
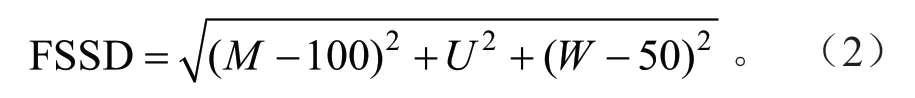
式(2)中:FSSD为林分空间结构距离,FSSD、M、U、W的取值范围均为[0,100];当M=100、U=0、W=50 时林分空间结构距离最小,即从现实林分到达理想林分距离最短;林分空间结构距离值越小,林分结构越理想。
3 结果与分析
3.1 九万山群落木本植物树种组成与结构特征
九万山群落的调查样地内胸径(DBH)≥2 cm的木本植物中,样方a 有135 种隶属于44 科76 属,属、种数占优势的科主要有山茶科(6 属14 种)、樟科(5 属15 种)、蔷薇科(5 属8 种)、壳斗科(4属13 种);样方b 有147 种隶属于47 科82 属,属、种数占优势的科主要有山茶科(8 属19 种)、樟科(5 属17 种)、蔷薇科(6 属8 种)、壳斗科(4属10 种)、杜鹃花科(2 属7 种);有90 个种隶属于38 个科59 个属在2 个样方中均有分布,综合来说,九万山群落杨梅坳区域观测到的木本植物有53 科99 属192 种。从表1可见,样方a 林分密度为4 517 株·hm-2,林木个体较多的有刨花润楠、建润楠、桂南木莲等,林木个体≤10株达83种,其中偶见种[23]有57 种,稀有种26 种;刨花润楠是数量上最多的树种,相对多度达13.03%,而赤杨叶相对优势度最大为13.07%,这两个树种的重要值也较大。样方b 林分密度为4 417 株·hm-2,林木个体较多的有多花杜鹃、红花荷、硬毛山香圆等,林木个体≤10 株达86 种,其中偶见种有47 种,稀有种39 种;重要值在前2 位的树种是多花杜鹃和红花荷,它们是数量上较多的树种,相对多度达13.03%和8.04%;综合2 个样方统计,有偶见种64 种,稀有种45 种。2 个样方没有重要值超过10%的树种,样方a 最大的刨花润楠也只有7.96%,样方b 最大的多花杜鹃为8.16%,说明该群落中没有绝对占优势的优势种,是较多的优势种形成了群落的共建种。
表1和表2中可以直观群落乔木树高在垂直方向上的空间分布变化,两块样方a、b 的大部分林木处在群落的下层(≤10 m),分别占总个体数的72.86%和78.90%,林木个体随着高度增加急剧减少,10~20 m 的中层木分别占总个体数25.08%和20.29%,≥20 m 以上的上层木非常少,分别只林木总个体数2.06%和0.82%。图1进一步展示了2 块样方林木个体树高分布的相似性,均呈左侧分布(单峰型),样方a、b 的林木个体树高多数集中分布在4~8 m 之间。

表1 a 样方林分及主要树种的数量特征Table 1 Quantitative characteristics of the stands and main tree species of plot a

表2 b 样方林分及主要树种的数量特征Table 2 Quantitative characteristics of the stands and main tree species of plot b
群落径级分布范围较大,样方a、b 分别在2~48.8 cm 和2~52.8 cm 径阶范围内呈连续分布,它们的林木胸径、树种数分布均呈单峰型(图1),林木个体胸径多数(51.10%、52.25%)集中分布在4~6 cm 径级,林木个体随着径级的增大急剧减少,胸径超过30 cm 的大径木样方a、b 分别只有64 株和41 株,占总个体数1.42%和0.93%;样方a 中6 cm 径级的树种分布最多达84 个,而样方b 中4 cm 径级的树种分布最多达106 个。在群落生长发育过程中,林木受树种生物学特性、竞争、环境及其在群落中占据的生态位不同,长成优势木的树种逐渐减少,所以样方a、b 中树种数总体分布趋势是随着径级增大树种数逐步降低,40 cm以上径级树种数分布样方a、b 分别有7 个和11 个,占树种总数的5.19%和7.48%;林分中没有个体在每个径级都有分布的树种。

图1 胸径、树高分布Fig.1 Distributions of DBH and tree height of the stands
3.2 九万山群落木本植物空间结构特征
3.2.1 林分空间分布格局

图2 林分的混交度、大小比数、角尺度分布Fig.2 Distributions of mingling degree,neighbourhood comparison and uniform angle of the stands
从图2可知,九万山群落样方a、b 林分中角尺度取值0.5 的林木个体所占比例分别为55.76%和55.32%,表明随机分布是九万山群落林分中大多数林木个体的主要分布方式;林分中不均匀分布和团状分布频率分别为26.81%和26.36%,均匀分布和绝对均匀分布频率占17.43%和18.33%,不均匀和团状分布的结构单元多于均匀分布,表明林分中局部出现的团状结构带动林分整体呈轻度聚集分布(样方a、b 林分平均角尺度分别为0.544和0.547)。从样方a、b 中重要值在前面的8 个种群的平均角尺度(表3~4)来看,它们的平均角尺度均在0.533~0.594 之间,均呈轻度聚集分布;从各种群分布频率看,除了水青冈种群随机分布林木个体比例为48%外,其它种群随机分布的林木个体均在50%以上,表明随机分布也是各个种群大多数林木个体的主要分布方式,各个种群都有10%左右林木团状分布,而只有0~2%的林木绝对均匀分布。

表3 a 样地主要树种的混交度、大小比数、角尺度分布Table 3 Distributions of mingling degree,neighbourhood comparison and uniform angle of the main tree species in plot a
3.2.2 林分个体大小分化程度
从图2中看出,群落内林木的各等级(从优势至绝对劣势)所占比例相差不大,林木优劣分化程度不大,个体大小分布较为均匀,表明林分整体相对稳定;样方a、b 林分平均大小比数分别为0.514 和0.533,说明在空间结构单元内,大部分个体分化程度较高,不同径级林木在空间结构单元内分布较均匀,但绝对劣势的林木个体比例稍大于优势木个体,导致整个林分处于中庸偏劣势过渡状态。从表3~4 中看出,各树种平均大小比数取值范围为0.099~0.727,表明林分内树种间的大小分化存在很大差异,其中,赤杨叶以优势木为主,占林木个体比例达77%,只有6%的中庸木和5%的劣势木,没有绝对劣势林木;建润楠、海南新木姜子以绝对劣势木为主,分别占林木个体比例达41%和30%,分别还有27%和18%的劣势木,只有4%和14%的优势木;木荷在2个样方均以优势木为主,分别占林木个体比例达36%和35%,同时分别还有29%和33%的亚优势木,分别只有7%和8%绝对劣势木,表明同一树种在生境差异不大时的生长状况基本一致;红花荷、马尾树、水青冈、罗浮锥以优势木为主,分别占林木个体比例为28%、38%、32%和31%;杉木以亚优势木为主,占林木个体比例为35%;刨花润楠、桂南木莲、润楠、新木姜子、硬毛山香圆各等级(从优势至绝对劣势)所占比例相差不大。
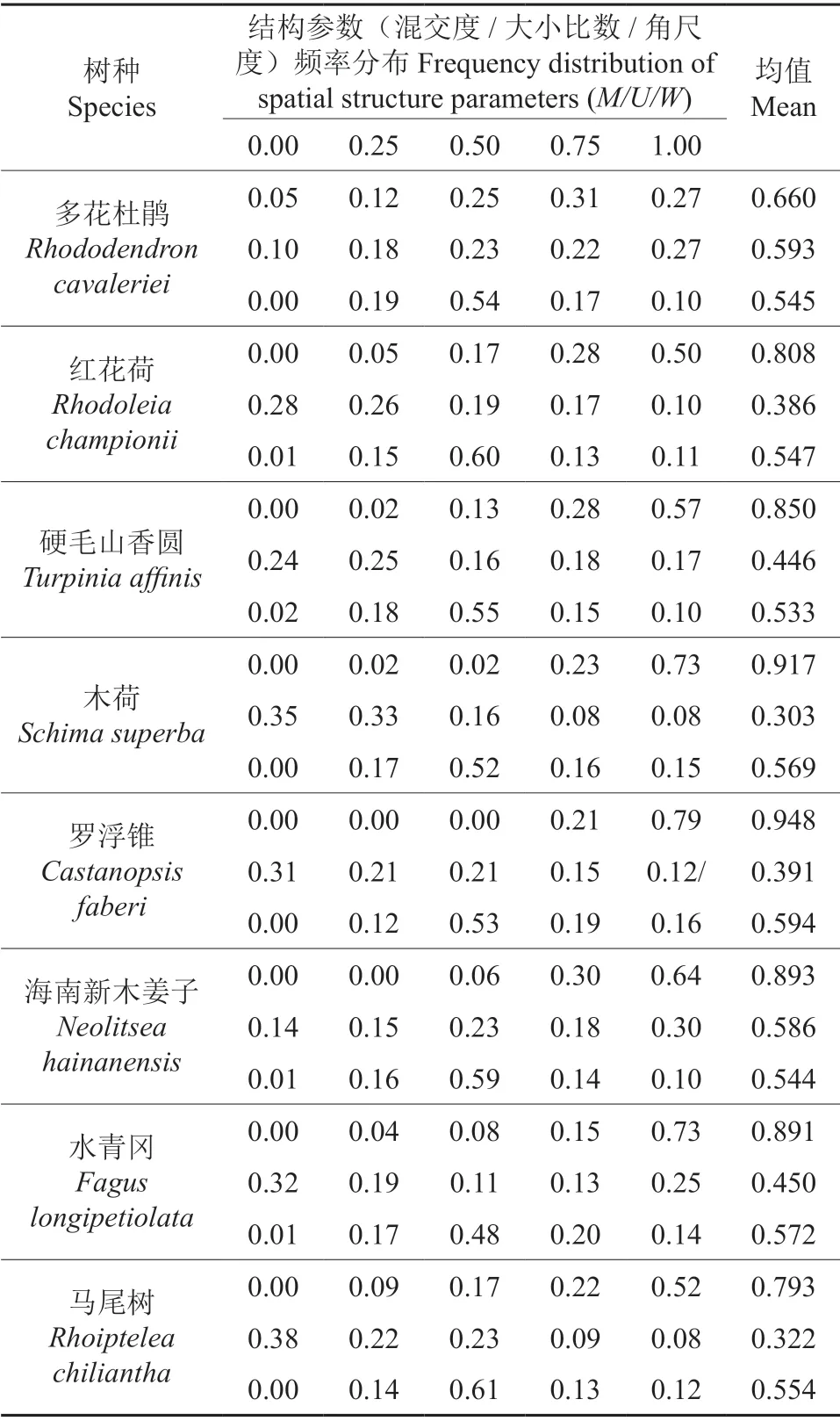
表4 b 样地主要树种的混交度、大小比数、角尺度分布Table 4 Distributions of mingling degree,neighbourhood comparison and uniform angle of the main tree species in plot b
3.2.3 林分树种空间隔离度
由图2可知,九万山群落样方a、b 林分混交频率由小到大逐渐递增,极强度混交频率分别占60.39%和62.37%,而零度和弱度混交频率之和分别只占4.34%和6.82%,林分混交度分别为0.851和0.842,已超过强度混交值0.101 和0.092,说明群落树种之间的隔离程度很高,林分混交状况良好。从样方a、b 中重要值在前面的8 个树种的平均混交度频率(表3~4)来看,所有树种的强度混交和极强度混交频率均大于零度和弱度混交,特别是赤杨叶和罗浮锥没有中度、弱度和零度混交的林木,故这两个树种的平均混交度频率接近了极强度混交;润楠和多花杜鹃的中度混交频率超过20%,带动了它两树种平均混交度频率低于强度混交值,处于中度混交向强度混交过渡的状态;其余树种均以强度、极强度混交为主,充分说明了整个群落林分中,同树种聚集在一起的情况不多,多数树种不与本树种为伴,大多数林木均为混交生长,林分群落结构相对稳定。
3.3 九万山群落木本植物空间结构指数与距离
林分空间结构指数(FSSI)能够客观、全面地反映不同林分空间结构的差异和同一林分不同时间尺度上空间结构的变化程度,林分空间结构距离(FSSD)是与林分空间结构指数(FSSI)相对应的一个指标。九万山群落两个样方的林分空间结构指数和林分空间结构距离分别为76.36、75.15 和53.66、55.80,两个样方相差不大,表明在这一区域范围内林分空间结构基本一致,表明研究区域的常绿阔叶林具有较好的空间结构,但与理想的空间结构有一定的差距(表5)。

表5 林分空间结构指数与距离Table 5 Stand spatial structure index and spatial structure distance
4 结论与讨论
群落结构和物种组成不仅反映群落中物种间的关系,也反映环境对物种生存和生长的影响,他们是群落最基本的特征和生态学研究的基础[24]。九万山群落杨梅坳区域样地内观测到的木本植物有53 科99 属192 种,常绿阔叶树占绝对优势,反映出广西雨水充沛、光照足的气候特征;群落中重要值没有超过10 的树种,表明群落优势树种不明显,结果与武夷山典型常绿阔叶林群落[23]、哀牢山常绿阔叶林群落[25]和尖峰岭群落热带山地雨林[26]相一致,均由较多的优势种形成群落的共建种,这也是很多天然阔叶林群落的特征;群落具有稀有种45 种,占总树种数的23.44%,稀有种比例高于武夷山群落[23]16.5%(23 种)和尖峰岭群落[26]20.7%(60 种),低于弄岗群落[27]33.63%(75 种),可能是受树种特性、土壤性质、样地大小等影响有关;大部分林木处在群落的下层,树高≤10 m 的林木占比超过70%;群落总径级结构呈倒“J”形,小径级树木占绝对优势,样地DBH 2~6 cm 的个体数占总个体数的51.68%,与武夷山[23]样地DBH 1~6 cm 的个体数占总个体数的73.89%、弄岗[27]样地DBH 1~3 cm 的个体数占总个体数的53.59%和尖峰岭[26]样地DBH 1~5 cm 的个体数占总个体数的76.50%基本一致。
群落的空间结构特征与群落形成的演替阶段有密切关系,演替至相对稳定阶段,物种通过竞争都占据着不同的空间,资源利用更为充分有效,水平分布格局为随机分布[14-15]。九万山群落主要树种和林分整体分布格局虽然是聚集分布,但角尺度只略大于0.517,较接近随机分布,表明群落处于演替中后期向顶极演替的阶段[28]。九万山群落林分中大小比数分别为0.514 和0.533,林木大小分化不明显且频率分布较均匀,整个林分处于中庸偏劣势过渡状态,这种状态表明树种的空间大小分化存在较大的差异,林木个体数量较多的种群大多数处于林冠中下层,对资源和空间的竞争势较弱,趋于劣势,致使林分总体大小比数分布较为均匀[29]。九万山群落林分混交度超过0.8,表明树种隔离程度较高,达到了较高混交程度,主要是群落林分中物种丰富,偶见种和稀有种较多且散生于群落中,促使群落中极强度和强度混交林木比例超过80%,说明该群落形成了由多树种混交的较为复杂的结构,各树种占据不同的生态位,形成协调的种间关系,促使群落向稳定的顶极群落演替。综合来说,九万山群落的空间分布为具有较高的混合结构、大小分化明显且呈轻度聚集分布特征,林分未达到稳定状态,这种状态在林分空间结构指数和林分空间结构距离分析中得到进一步的验证。
理想的林分空间结构表现出林分树种混交程度高,林分整体空间分布格局为随机分布,林木分化程度不明显[30],这也是林分空间结构优化的目标。用理想林分结构来衡量九万山群落林分结构,林分树种混交状况良好、隔离程度较高,林木分化属于中庸向劣态过渡状态,树种水平分布格局不够合理,与自然顶极群落有一定的差距。作为中国亚热带常绿阔叶林中的苗岭山地常绿阔叶林的重要组成部分的九万山自然保护区,合理的经营和保护是实现其可持续发展的关键。因此,针对九万山群落林分空间结构现状,制定以大小比数、角尺度为主要调整目标的林分空间结构调整方案,在保证原有混交度不降低的前提下,尽量选择角尺度大于0.75 以上且处于劣势或绝对劣势的非目的树种个体进行调整,提高九万山群落林分质量,促进其林分的顺行演替。
本研究只是针对九万山自然保护区杨梅坳区域内的空间结构所做的初步探讨,今后有待扩大调查研究区域范围,寻找被认为是九万山群落阔叶林分的标准林分,分析其标准林分的空间结构参数,使之得到更有效的保护和管理,达到稳定的群落状态。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
民间故事选刊·下(2021年5期)2021-07-08
民间故事选刊(2021年10期)2021-05-23
岷峨诗稿(2020年2期)2020-11-19
湛江文学(2017年1期)2017-11-24
