
低镉水稻研究进展
2021-01-28 02:23:48胡婉茵吴殿星舒小丽
核农学报 2021年1期
胡婉茵 王 寅 吴殿星 舒小丽
(1浙江大学原子核农业科学研究所,浙江 杭州 310029;2浙江省农业科学院农村发展研究所,浙江 杭州 310021;3农业农村部创意农业重点实验室,浙江 杭州 310021)
镉(cadmium,Cd)是一种二价阳离子,通过人类活动释放到环境中,属于可溶性剧毒金属,因其无法降解且具有持久性,已成为广为人知的环境污染物。镉广泛存在于我国稻田土壤中,极易通过土壤-作物系统积累在植物体内,且这种积累不可逆,进而通过植物源性食物。人体内摄入的镉与蛋白质结合生成镉-蛋白质,不仅影响人体磷(P)、钙(Ca)等元素的吸收代谢,还会对人体免疫系统、骨骼系统、泌尿系统和肾脏等产生毒害作用,导致高血压、蛋白尿、葡萄糖尿和骨骼软化等病症,并有可能致癌和致畸[1]。20世纪50年代震惊全世界的日本“骨痛病”事件即属最严重的慢性镉中毒事件[2],引起了全球的广泛关注。
水稻(OryzasativaL.)在我国农业生产中占据着重要地位,每年我国水稻的消耗量占谷物消耗量的55%[3]。近年来中国稻米镉污染现象日益严重,王茂起等[4]在2003—2004年期间对全国16个省市及自治区的29类食品进行检测,结果发现送检大米的镉超标率达到5.33%;2007年,甄燕红等[5]随机抽检全国部分市场的市售大米,结果13%的样品镉含量超标;谢燕湘等[6]于2012年在南方某市随机采集市售大米进行镉含量测定,结果镉含量超标率高达46%;覃芸等[7]在2016—2017年期间对柳州市市售大米样品进行质量安全检测,结果约9%的样品镉含量超标。且自2013年媒体先后揭露了湖南、广东和江西等地出现的“镉大米”事件,引起了公众对镉米的恐慌。因此,减少镉毒素在稻米中的积累,特别是减少稻田受污染地区稻米镉积累迫在眉睫。
目前控制水稻镉污染主要有两种途径:一是通过化学或者生物学方法阻碍稻田土壤中的镉通过根系进入植物体内;二是采用适当的农艺措施与耕作制度来减少镉在水稻体内的积累。而通过遗传育种技术培育低镉水稻品种是最直接有效的方式,能从源头有效保证水稻的安全生产。本文通过对影响水稻镉吸收和运输的环境、遗传因素进行阐述,并结合目前国内外有关低镉水稻选育的最新进展,探讨选育低镉水稻的可能途径,旨在为无镉或低镉水稻筛选培育提供理论和实践依据。
1 镉与水稻的生长
镉是最普遍的非营养重金属元素,其不参与构成植物体的生物结构,也不参与植物体的代谢活动,在所有有害重金属中,镉的危害性最大。水稻极易吸收并积累镉,但具基因型差异。一定浓度的镉胁迫对水稻的生理生化和形态等方面有强烈的毒害作用,严重影响水稻的生长并导致其减产,稻米品质劣变。
1.1 镉对水稻生理特性的影响
水稻体内积累过量镉会导致严重的毒性效应,影响植株的光合色素含量、呼吸强度、蒸腾和光化学效率。镉能够强烈抑制原叶绿素酸酯还原酶的活性并阻碍氨基-γ-酮戊酸的合成,从而影响叶绿素的生物合成及与蛋白质的稳定结合,破坏光系统Ⅰ与光系统Ⅱ中的光合电子传递,阻碍光合磷酸化过程,使得ATP合成受阻,扰乱叶绿体进行正常的光合作用[8-9],且光系统Ⅱ比光系统Ⅰ更容易受到镉的影响[10]。镉对光合作用的影响还可通过形成Cd-叶绿素复合物来抑制和破坏叶绿素-蛋白复合物[9]。镉胁迫浓度增大,水稻幼苗叶片的叶绿素含量和叶片光合特性指标(净光合率、气孔导度、蒸腾速率、胞间CO2浓度)均呈下降趋势[11]。此外,镉的积累还会抑制水稻植株细胞中苹果酸脱氢酶、琥珀酸脱氢酶、葡萄糖-6-磷酸脱氢酶和6-磷酸葡萄糖酸脱氢酶等呼吸酶的活性,造成植株呼吸代谢紊乱[12]。
1.2 镉对水稻产量的影响
水稻产量关系国民经济和人们的生活,镉胁迫也会影响水稻产量。但不同的研究结果不尽相同。黄冬芬等[13]发现土壤镉处理对盆栽水稻结实率和千粒重影响不大;李玉清等[14]也发现镉含量不同的水灌溉对水稻结实率和千粒重的影响并不明显;丁园等[15]发现土培不同镉浓度对水稻的千粒重、每穗实粒数和分蘖率影响不显著,但在高镉土壤(20.84 mg·kg-1)中生长的水稻,经济产量降低10.65%。而陈娟[16]研究发现,在镉处理下,水稻产量构成因素各指标均显著低于对照,尤其是每穗粒数,表明镉胁迫主要通过影响水稻幼穂分化进而影响其产量;葛才林[17]研究发现镉处理对水稻穂数的抑制效果最明显,但对千粒重的抑制程度较小;刘春梅等[18]研究认为镉胁迫处理浓度上升,水稻每穗粒数和粒重均随之减少,但千粒重无明显变化。这些不一致的研究结果可能是与使用的水稻品种不同,且试验过程中镉处理的方式不一致有关。因此,镉对水稻产量的影响需要继续深入开展,以获得普遍性的结果。
1.3 镉对水稻稻米品质的影响
随着人们消费结构升级,对稻米品质的要求越来越高,然而安全、健康营养的稻米已成为最基本的需求。水稻籽粒中积累镉,会影响稻米的加工、营养、外观和卫生品质。其中,稻米的卫生品质指籽粒中重金属的含量根据国家粮食卫生标准(GB 2762-2005)[19]其镉含量必需低于0.2 mg·kg-1。据报道,受镉污染的稻米醇溶蛋白含量较多,而清蛋白和球蛋白等含量较少,赖氨酸、直链淀粉含量较低,营养品质较差[13,20]。受到高浓度镉胁迫的水稻,其稻米的淀粉粘滞谱特征值与对照组相比发生了显著变化,崩解值和最高黏度值明显变小,而消减值、最终粘度与热浆黏度明显增大,表明镉降低了稻米的食味品质[13]。李玉清等[14]发现在水稻生长发育期间,采用低于0.012 mg·L-1镉的水灌溉对稻米的营养成分不会产生明显影响,但会明显影响稻米的外观品质。丁园等[15]发现水稻生长在镉污染(20.84 mg·kg-1)土壤中,其糙米中直链淀粉和镉含量均明显增加,而在低浓度镉污染(4.48 mg·kg-1)的土壤中生长的水稻,其籽粒蛋白质含量显著降低。镉处理浓度越高,稻米中粗蛋白、粗淀粉和赖氨酸含量越低[17]。
2 水稻对镉吸收、转运和积累机制研究
水稻对镉的吸收和积累的途径主要有3种:1)根转运体从根际土壤吸收镉进入根细胞;2)木质部装载介导镉从根转移到茎;3)镉经韧皮部从茎转运到籽粒。
2.1 水稻根对镉的吸收
根系吸收镉被认为是植物体内镉积累最重要的一步,水稻根系最先接触到稻田土壤中的镉离子并通过质外体和共质体途径吸收土壤溶液中的镉。其中,质外体途径主要利用镉离子在细胞内外产生的电势差以自由扩散的方式通过质外体空间进入到细胞内,运输速度快,无需消耗能量,属于被动运输过程。而共质体途径主要依靠细胞膜上的载体蛋白进行选择性跨膜运输,运输速度相对较慢,属于主动运输过程,需要消耗能量才能将镉转移到根系细胞内,再通过胞间连丝实现根系细胞间的运输[21]。
2.2 镉在水稻地上部分中的转运
水稻根吸收镉后,由植物短距离运输到木质部中完成加载,而后向地上部分运输。水稻地上部分的镉经木质部再通过筛管转移到韧皮部,最后通过节点和穂输送到籽粒中。从木质部到韧皮部的转移是镉到达水稻籽粒的必要条件。
研究表明,镉离子经木质部从根系到茎部的转移是决定水稻茎鞘和籽粒镉积累的主要因素[22-23]。镉在水稻植株中通过筛管进行转运,稻米中91%~100%的镉是由韧皮部转运而来[24-25],水稻韧皮部汁液中的镉浓度可能是决定其籽粒镉含量的关键因素[26]。Fujimaki等[27]采用正电子发射示踪成像系统对水稻灌浆期茎中镉转运和分布进行示踪研究,表明镉从茎基部的木质部运输至韧皮部,水稻茎节是木质部中镉向韧皮部转移的重要组织并在镉向籽粒转运过程中起着关键作用。在水稻营养生长期,下部叶组织是主要的镉储存器官,越接近地面的叶片在水稻早期生长阶段,镉含量越高,但在完全成熟阶段几乎不含镉[28]。
2.3 镉吸收、转运和积累相关基因
矿质元素需要依靠转运子才能从根部转移到茎[29],再从茎转运至叶片、籽粒。近年来,许多调控水稻吸收、转运和积累镉的基因已被克隆(表1)。
OsIRT1和OsIRT2是水稻Fe2+转运蛋白基因,由缺铁诱导表达,在酵母中能转运镉,并且OsIRT1对镉的特异性更大,可能在镉的吸收过程中占主导地位[30]。过表达OsIRT1水稻对镉更敏感且积累更多的镉,表明OsIRT1确实具有转运镉的活性[31]。
水稻NRAMP蛋白(natural resistance-associated macrophage protein)与金属离子转运具有重要关系,水稻中共有7个Nramp基因,其中OsNramp1和OsNramp5被证实参与植株体内细胞对镉的吸收和转运。OsNramp5位于根外皮层和根内皮层的远端,在根部高度表达,介导镉从外部溶液进入到根细胞内[32]。Takahashi等[33]发现OsNramp1表达水平差异是导致Habataki(高镉积累)和Sasanishiki(低镉积累)两种水稻镉积累差异的主要因素;OsNramp1是水稻镉积累的重要蛋白,在质膜上充当镉流入转运体,在镉从根到茎的转运过程中起着关键作用[34]。OsNramp6蛋白能够在酵母中转运镉,其定位于胞质膜上,主要在水稻幼叶中表达,可能与叶片积累镉有一定的关系[35]。
重金属转运ATP酶(heavy metal transporting ATPase,HMA)蛋白被认为是重金属穿透细胞膜的外排转运蛋白[36],水稻中有9个HMA基因。其中OsHMA3在根中高度表达,位于根部液泡膜上,能与流入胞质溶胶中的镉螯合,以复合物的形式暂时储存,降低胞质镉迁移率,限制镉从根部向地上部位转移从而降低植株中镉积累,过表达OsHMA3可以显著降低籽粒镉含量且不影响其他微量元素[37]。OsHMA2定位在质膜上,在水稻中充当镉的内流转运体,其主要在根部和节间表达,参与镉的根-茎转移,将镉从薄壁组织重新装载到维管束的韧皮部中并介导镉在水稻植株中的分配[38-40]。Lee等[41]研究发现OsHMA9定位于质膜上,主要在维管束和花药中表达,并证实OsHMA9是一种镉外排转运体。
水稻低亲和阳离子转运蛋白OsLCT1是水稻韧皮部中镉转运的主要转运子,属于外排型镉转运体,其定位于质膜上,在水稻营养生长阶段的叶片和茎中高度表达[42]。OsLCT1RNAi干扰系籽粒镉含量大约是对照组的50%,且籽粒中矿物质营养成分以及植株生长和产量不受影响[42]。李曜魁等[43]以华占和籼稻93-11为材料,从中克隆了OsLCT2基因,该基因编码蛋白与小麦TaLCT1蛋白的同源性超过60%,由于小麦TaLCT1对镉具有高亲和力,介导镉流向细胞质内,因此推测水稻OsLCT2可能也有类似的镉转运功能。
水稻OsLCD与其他已知的镉转运吸收基因均不同源,其位于细胞核与细胞质中,主要在根部和叶片中表达,OsLCD的T-DNA插入突变体lcd(OsLCD的第一个内含子中插入T-DNA)籽粒镉积累量比野生型植株显著降低,而锌铁含量、干重和种子产量无明显变化,表明OsLCD是一个低镉水稻育种的候选基因[44]。
金属耐受性蛋白(metal tolerance proteins,MTPs)属于阳离子扩散促进剂转运蛋白家族,这类蛋白能够调节植物体内重金属的稳定状态[45]。水稻中已鉴定出10个MTP基因,其中OsMTP1属于Zn-CDF子群,主要分布在质膜上,能够有效转运镉,并且可以使植物体内离子维持稳定状态[46]。Yuan等[47]通过转基因获得OsMTP1 dsRNAi水稻植株,该转基因株系幼苗对镉的耐受性下降,而OsMTP1在野生型酵母中表达增强了酵母对镉的耐受性。刘宝秀[48]对水稻OsMTP2进行分析,结果表明该基因为MTP家族的一个膜蛋白基因,主要在叶片、茎中表达,可能在水稻营养生长期对金属离子转运起重要作用。
ABC转运蛋白(ATP-binding cassette protein)也参与植株金属离子的转运,目前水稻2个亚种基因组中已鉴定出121个该类蛋白质[49],大部分是由膜蛋白组成的,且能够通过消耗ATP产生的能量把有毒重金属排出细胞。其中OsPDR5和OsPDR9对镉可能有转运作用。Oda等[50]研究发现镉胁迫下水稻OsPDR5在根中大量表达,是对照的3.5倍,但在茎中的积累不受影响,说明水稻OsPDR5可能参与镉转运并增强水稻对镉的耐受。Moons[51]发现镉能够显著诱导OsPDR9在水稻幼苗根系中的表达,表明OsPDR9参与水稻镉胁迫响应。此外,CAL1蛋白能够特异性结合细胞质中的镉,驱使镉从原生质体中转移到细胞外侧,随后装载到木质部汁液中,促进镉从根部转运至茎节当中,调控镉在水稻中的积累[52]。
研究发现,长非编码核糖核酸(lncRNAs)和微小核糖核酸(miRNAs)也能够参与植物镉胁迫应答反应,是植物体细胞中的关键调节因子。Liang等[53]通过深度测序,确定了镉诱导水稻根中全基因组的lncRNA表达谱,结果表明,lncRNAs可能对镉胁迫起重要作用。Fei等[54]用镉处理水稻幼苗并对其根系进行深度RNA测序,序列分析结果表明,有9个lncRNA基因与镉胁迫反应高度相关,对这9个基因进行鉴定,发现转录后基因调节也可能参与镉胁迫反应。Ding等[55-56]鉴定出19个响应镉胁迫的miRNAs,在镉胁迫下有18个miRNAs表达下调,而miRNA528表达上调,后续研究发现镉胁迫显著诱导miRNA268的表达,且在镉胁迫下水稻体内miRNA268的靶基因(NRAMP3)表达量显著降低,而miRNA268过表达水稻植株幼苗镉含量比野生型高,表明miRNA268是水稻耐镉胁迫的负调节因子。研究发现,miRNA390的过表达水稻对镉胁迫的敏感性增强,其镉积累量比野生型高,且在镉胁迫下水稻体内的OsHMA2和OsNRAMP5表达水平上升,说明miRNA390也是水稻耐镉胁迫的负调节因子[57]。现有研究证实了miRNA166在水稻木质部的镉负载中起作用,其过表达降低了镉在水稻中的转运,使得镉从根向茎的转移减少,并且过表达植株的稻米镉含量显著降低,水稻体内miRNA166对镉的积累调节作用主要是通过调节其靶基因OsHB4实现的[58]。已鉴定的镉响应lncRNAs和miRNAs对于进一步了解水稻体内镉代谢的分子机制具有重要意义,目前对lncRNAs和miRNAs的研究还不够深入,大部分的lncRNAs和miRNAs生物学功能尚不清楚,未来需要加大研究力度,以便为系统了解水稻镉代谢的机制提供有价值的参考意见。
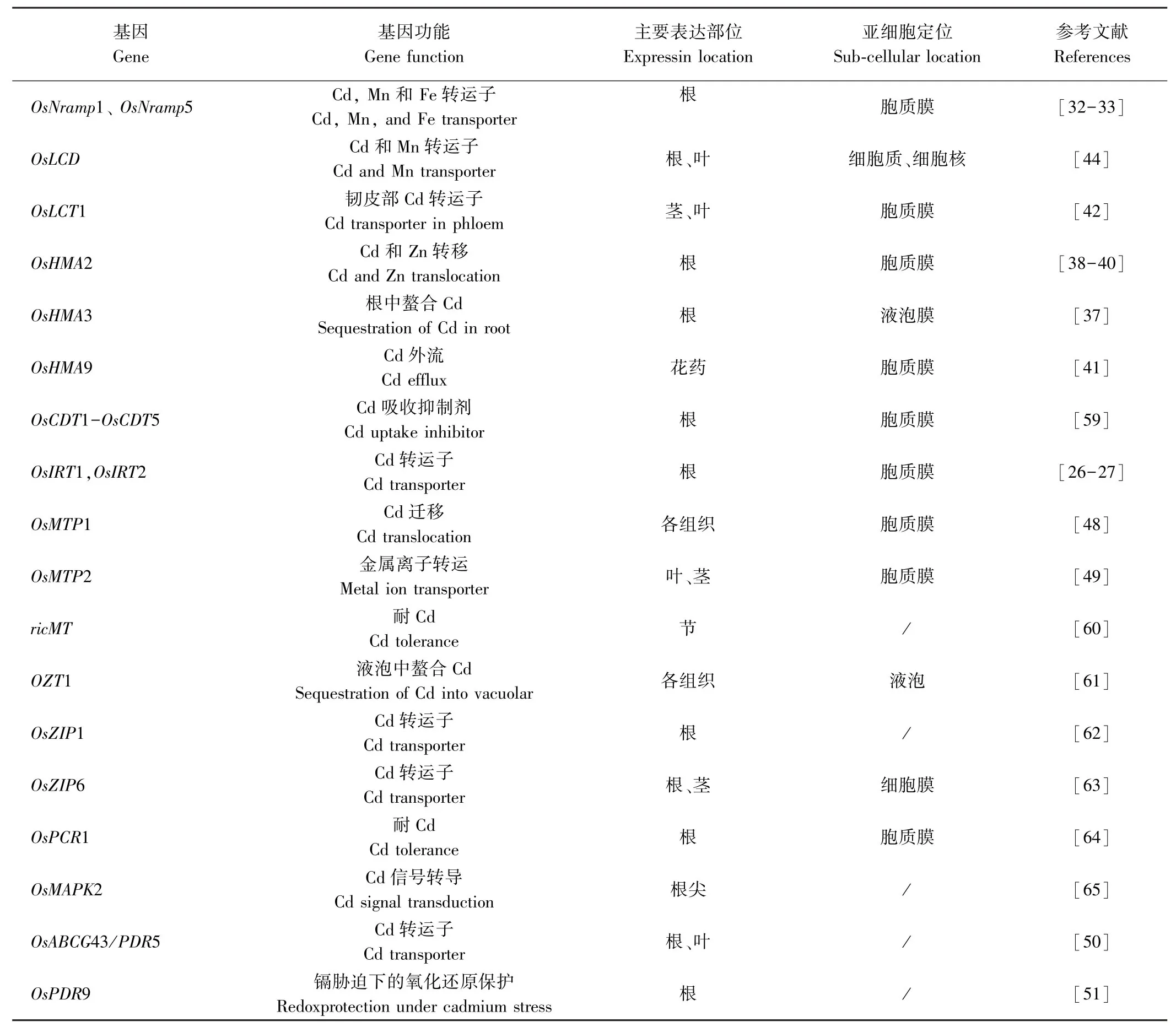
表1 水稻中已克隆的负责镉吸收与转运相关基因Table1 Cloned genes responsible for cadmium uptake and translocation in rice
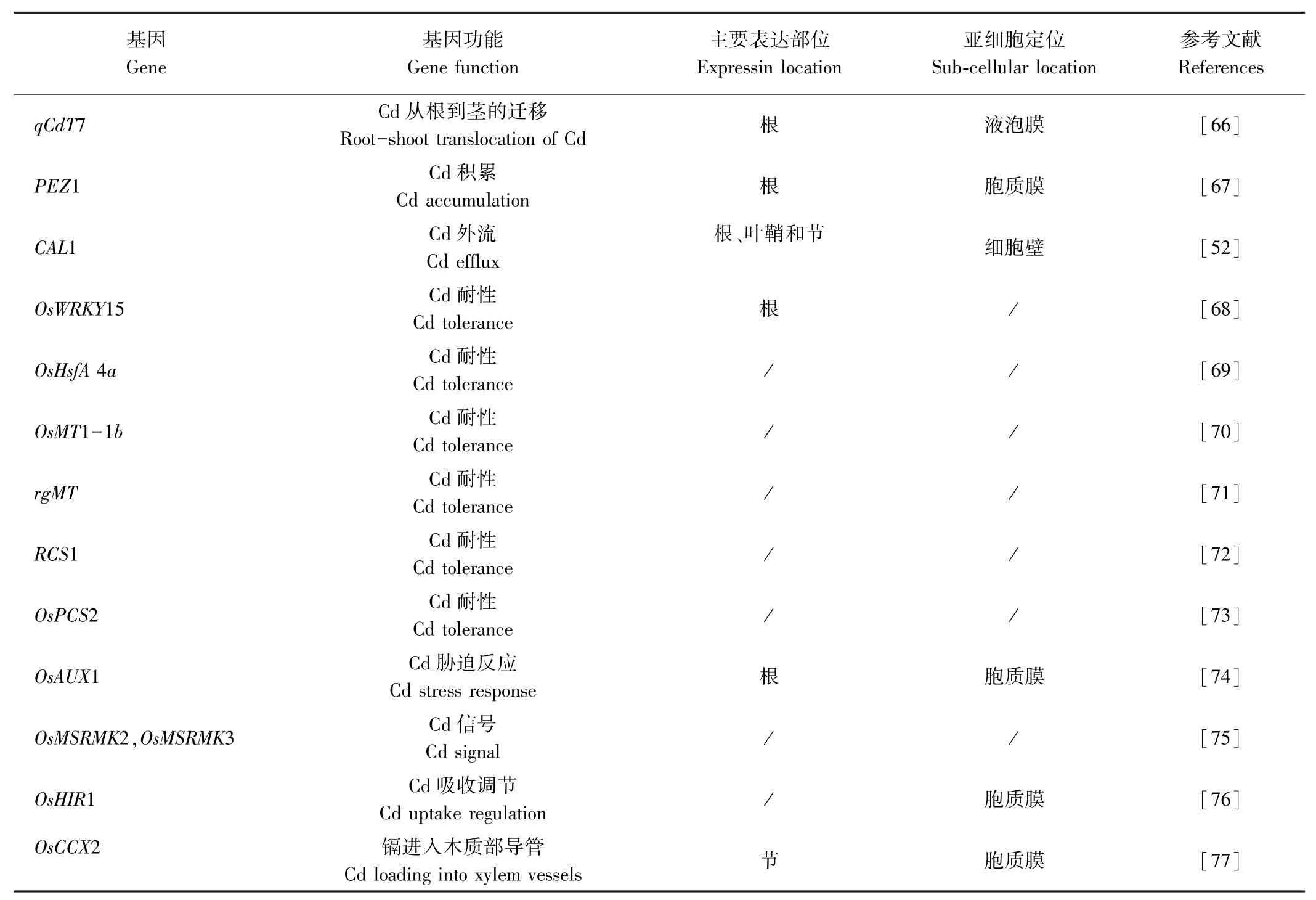
表1(续)
2.4 水稻镉吸收、转运和积累的影响因素
2.4.1 水稻自身遗传特性 不同水稻品种遗传背景不同,其生理特性和形态结构也不同,从而导致水稻品种间对镉的吸收和积累也不尽相同。仲维功等[78]选用43个水稻品种进行田间试验,发现粳稻根部镉积累能力最强,籼稻精米和籽粒中镉积累量最高,其次是杂交稻和粳稻;Ye等[3]选用9个水稻品种进行试验,得到了与仲维功等[78]相同的试验结果;邓伟等[79]发现高镉积累品种对镉的吸收特性表现为积累速度快或者时间长,而低镉积累品种则表现为积累速度慢或时间短;Zhong等[80]研究发现粳稻比籼稻更容易积累镉,但是镉使得籼稻产量明显下降,而粳稻产量均增加。
2.4.2 环境因素 土壤环境是影响水稻镉含量最主要的因素。水稻对镉的吸收与镉的总含量相关不大,而与镉的生物有效性直接相关。稻田土壤的物质组成(质地、有机质)、化学性质(pH值、氧化还原电位、阳离子交换量)及微生物活动都会影响土壤中镉的生物有效性[81]。黄德乾等[82]采用温室盆栽试验研究了3种不同类型土壤上水稻对镉污染的响应,发现不同类型土壤上水稻中镉的浓度大小为红壤>红壤性水稻土>乌栅土。
水稻对镉的吸收取决于土壤的物理和化学条件,其中土壤酸碱度(pH值)和土壤有机物质含量是决定镉吸收量的最主要因素[83]。土壤pH值在水稻根系对镉的吸附以及镉在土壤溶液中的形态、溶解度和迁移率方面起着最重要的作用[84]。研究表明稻田土壤酸碱度与水稻镉吸收量密切相关,在一定范围内,两者呈负相关[85]。因此,防止稻田土壤酸化可有效降低生物可利用镉的释放。在受镉污染稻田施加CaCO或Ca(H2PO4)2等碱性物质可提高稻田土壤的pH值,从而减弱水稻镉毒害作用。土壤有机物质能够与镉络合或螯合,因此,可以通过掺入有机物质,如施用有机肥(农家肥、堆肥、植物秸秆等),增加镉与有机物质的结合,进一步减少稻田中的可吸收镉。
此外,土壤氧化还原电位的变化也会引起镉的溶解度发生改变。研究表明,在淹水期间,稻田土壤氧化还原电位值降低,镉被吸附或形成硫化镉(CdS)沉淀,溶解度降低,因此镉不容易被吸收。然而,排水期间的稻田土壤需要氧气,其Eh值升高,镉将形成水溶性CdSO4,释放出可被根系吸收的镉离子[86]。加强稻田淹水管理也能够有效降低稻米镉积累。
土壤微生物在活动过程中能够分泌出有机物质、蛋白质和多聚糖等,这些物质不仅能够改变根际土壤的pH值,还能够吸附镉,对镉具有氧化还原、络合或沉淀等作用,从而影响根系对镉的吸收。研究表明,水稻根际微生物群落与根系分泌活动密切相关,两者相互促进[87]。根系分泌物包括有机配体(苹果酸、腐殖酸、柠檬酸、黏胶、糖类、酚类和氨基酸等)和无机配体(C1-、SO42-、NH4+和PO42-),这些分泌物进入到稻田土壤后,不仅作为微生物活动的能量来源,而且有些能够与镉螯合,使得根际周围土壤的pH和氧化还原电位值发生变化,改变稻田土壤中可利用镉离子的含量[88]。
3 低镉水稻育种
稻米镉积累会导致严重的食品安全问题,对人类健康存在极大隐患。为了防止人体摄入镉,培育和选择低镉水稻品种对于人类健康有深远意义。
3.1 常规杂交和诱变育种
杂交育种是选育低镉水稻的技术之一。可以根据水稻籽粒镉积累的规律,选育低镉不育系,低镉恢复系和配组获得低镉杂交水稻[89]。
诱变育种具有突变频率高、稳定时间短、操作方法简便、可打破性状连锁并实现基因重组等优点,是培育新品种,实现种质资源创新的一项重要技术。高能重离子束具有诱导突变频率高且辐射剂量低(经辐照后的植物能够正常生长)的优势,目前已经应用于辐照植物体以获得非转基因突变体。林园园等[90]利用重离子束辐照技术诱变水稻,发现重离子束能够诱发水稻对镉吸收能力的突变。由于诱变具有随机均匀特性,因此通过诱变技术可以在突变体中筛选出符合要求的低镉水稻新品种。Ishikawa等[91]也采用该辐照技术,筛选得到3个突变体,且这3个突变体的稻米镉积累浓度均低于0.05 mg·kg-1,属于低镉新品种。
将杂交育种与诱变育种方法结合运用,在杂交重组的基础上对尚未稳定的水稻杂交后代材料进行适当剂量的诱变处理,可以增加突变频率、扩大突变谱、打断基因连锁,从而促进基因重组过程产生更多突变类型。据统计,1970—1983年期间,国外以诱变突变体作为杂交亲本培育出的新品种占诱变育种新品种的60.5%[92]。庞伯良等[93]选用水稻作为试验材料,并将辐射诱变与常规杂交技术相结合进行试验,结果表明,复合处理产生的突变类型与频率均比各种单处理要高,且能够产生更好的育种效果。因此,诱变与杂交相结合也可以在一定程度上为选育低镉水稻品种创造丰富的育种材料,提高低镉水稻育种效果。
3.2 基因工程育种
参与水稻镉代谢的基因已鉴定30多个,其中有些是调控镉积累的关键基因,利用转基因手段对这些关键基因进行遗传操作可获得低镉水稻材料。例如,过表达OsHMA2转基因水稻,其籽粒中镉含量仅为野生型的一半[39],利用RNAi干扰技术,抑制OsLCT1的表达,获得的转基因植株籽粒镉含量大约是对照组的50%[42]。Shimo等[44]将T-DNA插入到OsLCD中,通过筛选,鉴定出了一个新的耐镉突变体lcd。
此外,可利用CRISPR技术在水稻中定点突变吸收镉基因或者导入负责排镉的基因,阻断镉进入到植物体内从而获得排镉水稻[94]。已有研究以杂交水稻的骨干亲本作为受体材料利用CRISPR创制出低镉不育系、恢复系,并采用杂交技术,配制出了高产、优质且低镉的杂交稻新品种,使得低镉杂交稻选育取得了突破性的进展[95]。
3.3 分子标记辅助育种
探索水稻镉积累特性费时费力,为此提出了建立分子标记辅助育种系统,该系统可以利用追踪水稻遗传组成的分子标记从突变体中培育水稻新品种,缩短育种时间,达到精准育种,提高选育效率并节约筛选成本。陈彩艳等[96]开发了4个低镉分子标记,并运用分子标记辅助选择技术对5 769份水稻新品进行筛选,最终获得携带1~3个低镉分子标记的80份新品系,经过田间试验证明了其中有些品种杂交后具备稳定的低镉遗传特性。Ishikawa等[91]研究确定降低镉吸收的基因为OsNRAMP5,为此构建了OsNRAMP5基因的分子标记辅助选择体系,在突变体后代群体中筛选携带OsNRAMP5的品种。由于离子束辐射产生的突变体是非转基因植物,更容易被消费者接受。
4 结论与展望
在过去几年内,国内外加大了对水稻镉代谢分子机制方面的研究,同时也取得了重大进展,发现了一些新的水稻镉转运体家族,然而由于镉与其他微量元素之间存在协同/拮抗等作用,水稻对镉的吸收、转运、积累机制复杂,解决稻米镉污染问题仍任重道远。未来低镉水稻的研究可以围绕以下几个方面深入开展:1)深入挖掘与水稻镉代谢相关的金属转运体,实施多基因聚合育种;2)开发与水稻低镉基因紧密连锁的分子标记,推动分子标记在育种中的应用并将分子育种与常规育种有机结合;3)加强各农艺措施的综合实施,降低外源镉的生物有效性,建立安全、高效的栽培技术体系,最终培育出稳定遗传,安全优质的低镉水稻新品种。
猜你喜欢
青少年科技博览(中学版)(2022年6期)2022-12-27 19:44:27
美食(2022年5期)2022-05-07 22:27:35
少儿科学周刊·儿童版(2021年21期)2021-12-11 01:45:00
现代畜牧科技(2021年4期)2021-12-05 15:34:45
军事文摘(2021年22期)2021-11-26 00:43:51
现代畜牧科技(2021年10期)2021-11-19 08:42:26
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中国粮食经济(2018年5期)2018-12-27 08:57:26
河北农业科学(2018年2期)2018-07-26 11:28:14
