
葡萄BBX基因家族的鉴定与表达分析
2021-01-28 02:26:08葛孟清刘众杰朱旭东卢素文房经贵
核农学报 2021年1期
葛孟清 刘众杰 朱旭东 卢素文 管 乐 房经贵
(南京农业大学园艺学院,江苏 南京 210095)
光在植物生长发育过程起着重要的作用,它不仅参与植物各个阶段的生长发育,还为光合作用提供能量。光参与的发育过程包括芽、茎、叶和根的生长发育,向光性、叶绿素的合成、分枝以及花的诱导等。光作为一种外部信号,与植物体内激素网络协调,影响植物生长和发育[1]。植物中的几种蛋白质因其在光调节发育中的重要作用而被鉴定出来[2-3],其中包括响应光信号的重要转录因子BBX基因家族[4]。BBX蛋白(B-box-type protein)在植物和动物中参与细胞生长、细胞分化和转录调控等活动,在植物光形态建成、花发育及对逆境响应等过程中发挥重要作用[5],是控制生长发育调控网络中的关键因素。
BBX蛋白是锌指结构蛋白家族的一个亚家族,BBX基因家族特征是在N末端具有一个或两个B-box结构域,有时还在C末端具有一个CCT结构域[6]。目前已在水稻[7]、拟南芥[8]、番茄[9]、苹果[10]及梨[11]等多种植物中鉴定出BBX基因家族,但对葡萄中的BBX家族还未进行系统的研究。模式植物中对BBX基因功能的研究较多,例如,拟南芥中的AtBBX1/CO是较早发现的BBX蛋白,其与SPA1互作增强FT基因表达,可以促进长日照下的植株开花[12];拟南芥AtBBX24和AtBBX25通过调控HYH蛋白间接地对hy5产生负调控,对幼苗的光形态发生进行微调[13];拟南芥AtBBX21在避荫条件下下调生长素、油菜素类固醇和乙烯信号通路成分的基因表达,过表达AtBX21可以改善植物的形态和生理特性,并提高光合速率[14];在高剂量的UV-B辐射下,拟南芥AtBBX31促进紫外线保护物类黄酮和酚类化合物的积累[15]。近年来,BBX基因在其他植物中的研究也在不断增加,水稻中OsBBX14基因对抽穗期有调节功能,对水稻的光形态发生有积极的调节作用[16];PpBBX16在梨皮中的短暂过度表达增加了花青素的积累,而病毒诱导的PpBBX16基因沉默减少了花青素的积累[17]。有试验证明SlBBX20是番茄类胡萝卜素积累的正向调控因子,与野生型相比,SlBBX20过度表达会导致果实和树叶绿色更深以及更高水平的类胡萝卜素合成[18]。
BBX基因家族广泛参与了植物生长发育的各个阶段以及对逆境的响应过程,有望作为植物的潜在遗传标记,特别是在功能分析和确定其在多变量胁迫下的作用方面。故本研究通过对葡萄BBX基因家族的鉴定和表达分析,以期了解葡萄VvBBX基因对光信号的响应机制,为葡萄果实营养品质的改良提供方法。
1 材料与方法
1.1 试验材料
供试葡萄品种为1年生阳光玫瑰盆栽苗,购自山东志昌葡萄研究所,种植于南京农业大学白马教学科技基地。分别置于自然光、黑暗、红光、蓝光、远红光条件下照射24 h,取植株的幼嫩叶片,液氮速冻后存于-80℃备用。
1.2 试验方法
1.2.1 葡萄BBX基因家族成员的鉴定 在Pfam(http://pfam.xfam.org)网站下载基于动植物B-box保守结构域的隐马尔可夫模型文件(Pfam00643),然后利用hmmer 3.0 windows程序对葡萄全基因组蛋白序列进行搜索,得到候选的葡萄BBX蛋白序列。将所有的候选蛋白序列去重复之后,将每一条候选序列分别提交到SMART(http://smart.embl-heidelberg.de/)数据库和Inter Pro(http://www.ebi.ac.uk/interpro/)数据库,验证这些候选蛋是否含有B-box结构域。具有Bbox结构域的候选蛋白即为葡萄中的BBX蛋白,将编码这些蛋白质的基因定为VvBBX基因。将得到的VvBBXs蛋白序列提交至ExPASY(http://web.expasy.org/protparam/)在线分析网站,预测其分子量(molecular weight,Mw)和等电点(pI)。利用WoLF PSORT(http://www.genscript.com/wolf-psort.html)在线网站进行亚细胞定位分析。
1.2.2 葡萄BBX基因在染色体上的分布 根据VvBBX基因的基因号,在CRIBI(genomes.cribi.unipd.it/grape/)网站上搜索,将VvBBX的定位数据找到,利用MapChart软件作出VvBBX在染色体上的分布图。
1.2.3 葡萄BBX基因结构和保守结构域分析 从葡萄基因组中提取VvBBXs基因结构信息,利用在线软件GSDS绘制基因结构图。将葡萄BBX蛋白序列提交到MEME(http://meme.nbcr.net/meme)网站鉴定葡萄BBX蛋白的保守基序,参数设置为基序数量10个,使用TBtools软件将蛋白保守序列的结果做成可视化图。利用MEGA 6.0软件构建系统进化树,进化树生成算法为NJ(Neighbor-Joining),校验参数Bootsrtrap重复1 000次。
1.2.4 葡萄BBX基因系统发育树的构建 在葡萄全基因组数据库Grape Genome(http://genomes.cribi.unipd.it/grape)和Phytozom (https://phytozome.jgi.doe.gov/pz/portal.html)下载葡萄、拟南芥和水稻全基因组数据,获取拟南芥、水稻BBX基因家族的蛋白序列,利用MEGA 6.0软件对3种植物的87个BBX蛋白序列进行系统发育分析。
1.2.5 葡萄BBX基因家族的同源性分析 利用TBtools软件对VvBBXS进行葡萄全基因组,葡萄和拟南芥基因组的同源性分析。
1.2.6 葡萄BBX基因启动子序列分析 获取BBX基因家族成员起始密码子ATG上游2 000 bp的序列提交到Plantcare(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)网站上,对所有家族成员的启动子序列上的作用元件进行分析比较。
1.2.7 葡萄BBX基因的表达模式分析 葡萄芯片数据来源于NCBI的GEO数据库,芯片编号为GSE36128,葡萄芯片数据包括53个葡萄样品,覆盖了葡萄生长过程不同阶段的不同器官。利用TBtools软件绘制热图。
1.2.8 不同光质照射下葡萄BBX基因的实时荧光定量PCR 将葡萄植株分别置于自然光、黑暗、蓝光、红光、远红光的条件下照射,以自然光照下生长的葡萄植株作 对 照,对BBX2、BBX6、BBX7、BBX8、BBX9、BBX11、BBX13、BBX17、BBX21、BBX24这10个基因进行实时荧光定量PCR。对VvBBXs基因家族的编码序列进行引物设计(表1)。cDNA合成用HifairII®1st Strand cDNA Synthesis SuperMix for qPCR反转录试剂盒。反转录产物-20℃保存备用。实时荧光定量PCR应用Kubotech Quantagene q225仪(北京酷搏科技有限公司)进行扩增,以葡萄Actin基因为内参,扩增总体系为10μL体系,上下游引物各0.4μL、200 ng·μL-1cDNA 1μL、qPCR SYBR Green Master Mix 5μL、ddH2O 3.2μL。反应程序为95℃预变性300 s;95℃变性10 s,58℃退火30 s,共40个循环。试验设3次重复,数据使用OriginPro 8.0软件分析。
总RNA的提取采用FastPure Plant Total RNA Isolation Kit试剂盒(南京诺维赞生物科技有限公司),反转录HifairII®1st Strand cDNA Synthesis SuperMix for qPCR试剂盒、荧光定量染料SYBR GreenI均购自于上海翊圣生物科技有限公司。所用引物由通用生物系统(安徽)有限公司合成(表1)。

表1 VvBBX基因实时荧光定量PCR引物Table1 VvBBX gene fluorescence quantitative real-time PCR primers
2 结果与分析
2.1 葡萄BBX基因家族的鉴定与分析
对葡萄基因组的蛋白序列进行搜索,共得到51条候选序列,最终在葡萄基因组中共鉴定得到25条非冗余葡萄BBX蛋白的编码基因,根据它们在染色体上的分布,命名为VvBBX1~25(表2)。所有成员的编码区序列长度介于384~1 425 bp之间,蛋白长度介于127~474 aa之间,其中VvBBX3的蛋白长度最长,为474 aa,蛋白长度最短的是VvBBX13,为127 aa。所有蛋白等电点介于4.29~9.01,其中等电点大于7的只有4个,分别是VvBBX4、VvBBX10、VvBBX12、VvBBX13。将所有蛋白序列上传至TargetP 2.0网站,发现有4个成员定位到分泌途径上,分别是VvBBX2、VvBBX5、VvBBX7、VvBBX20,有2个成员定位到叶绿体上,分别是VvBBX23、VvBBX24,还有1个成员VvBBX11定位到线粒体上。通过SMART网站对所有成员进行蛋白结构预测,结果可以分为4类,第一类包括4个成员,特点是C端有2个B-box结构域,N端有一个CCT结构;与第一类相比,第二类N端不具有CCT结构,包括13个成员;第三类包括3个成员,特征是C端具有一个B-box结构,N端有一个CCT序列;最后一类是只在C端有一个B-box结构,包括5个成员。基于25个VvBBXs家族基因在染色体上的位置信息,比较分析葡萄的25个VvBBX蛋白序列。使用Mapchart软件绘制出BBX家族基因在染色体上的分布图(图1)。结果表明,22个成员主要分布在11条染色体上(chr1、chr3、chr4、chr5、chr7、chr9、chr11、chr12、chr14、chr18、chr19),其中chr11染色体上分布基因成员最少,只有VvBBX11基因,最多的是chr12,有4个成员,分别是VvBBX12、VvBBX13、VvBBX14、VvBBX15。此 外,VvBBX23、VvBBX24、VvBBX25这3个家族成员未能定位到具体染色体上。此外,所有成员大都分布在染色体的上端、中上端和下端,位于染色体中端的成员仅有5个,分别是VvBBX12、VvBBX13、VvBBX14、VvBBX15和VvBBX18,且VvBBX12、VvBBX13、VvBBX14形成1个基因簇,可见BBX基因家族主要以分散复制和片段复制的方式来完成扩增。
2.2 葡萄BBX家族基因结构的分析
使用MEGA 6.0、GSDS 2.0、TBtools软件进行分析,得到葡萄BBX基因家族的基因结构和蛋白Motif分布(图2、图3)。葡萄25个BBX基因家族成员被分到3个类别,每一类都包含2个亚类、8个成员,且第一类成员中含有的Motif数量要高于其他类,第二类有6个成员,其余11个成员是第三类。由图2可知,同一类的成员之间所含Motif种类基本相同,且所有基因都含有Motif1和Motif4。除VvBBX19基因外,其他所有基因都至少含有一个UTR区,UTR区域长度大小不一。所有葡萄BBX基因成员中只有BBX10、BBX16和BBX17基因不含内含子。
2.3 葡萄BBX家族的系统发育分析
?
由图4可知,所有BBX蛋白分成了A、B、C、D、E共5个分支。其中,25个葡萄BBX蛋白家族成员分布在A、B、C、D这4个分支中,E分支中没有葡萄BBX蛋白成员。葡萄BBX蛋白家族成员在A分支中存在最多,为8个;在B分支中存在最少,仅有3个,其余2个分支各有7个成员。
2.4 葡萄BBX基因家族的同源分析
基因家族是来源于同一个祖先,由一个基因通过基因重复构成的一组基因,通常它们在结构和功能上具有明显的相似性,编码相似的蛋白质产物[19-21]。同一家族的基因可以紧密排列在一起,形成一个基因簇,但多数时候,它们是分散在同一染色体的不同位置,或者存在于不同的染色体上[22]。基因复制对基因家族的产生作用很大,基因重复为新基因提供原材料,促进新功能的产生[23]。基因的复制主要包括基因组复制/片段复制、串联复制以及基因和染色体水平的重排。串联复制和片段复制在基因家族的进化和扩展中经常发生[24]。串联重复通常会引起基因簇,片段性重复可能导致家族成员的分散。由图5可知,葡萄BBX基因家族中在12号染色体上存在一个由VvBBX12、VvBBX13、VvBBX14形成的基因簇,在葡萄基因组中鉴定出4对片段重复形成的基因对(VvBBX21/VvBBX14、VvBBX24/VvBBX14、VvBBX4/VvBBX18、VvBBX7/VvBBX18),表明在葡萄BBX基因家族的扩增中,片段复制事件可能比串联复制更重要。
2.5 葡萄BBX基因家族启动子序列分析
在NCBI上获取VvBBX1~25基因起始密码子上游2 000 bp的启动子序列,利用Plantcare进行分析,共发现31种作用元件,不包括CAAT-box、TATA-box,共发现了8类作用元件(表3)。其中光响应元件有ACE、AE-box、Box-4、G-box、I-box、MRE、Sp1、LAMP等21种,响应激素类的作用元件分为8种,参与茉莉酸甲酯反应的有CGTCA-motif、TGACG-motif,参与赤霉素反应的有GARE-motif、TATC-motif、P-box,与脱落酸、生长素、水杨酸相关的顺式作用元件均只有1种,分别是ABRE、TGA-element、TCA-element。此外,还发现了响应低温的作用元件LTR,与防御功能相关的元件TC-rich repeats,与伤口反应相关的WUN-motif,还有参与昼夜节律控制的作用元件circadian以及厌氧诱导所必需的作用元件ARE。还有一些出现次数很少的作用元件分别是与分生组织表达相关的元件CATbox、参与胚乳表达的作用元件GCN4-motif、与栅栏叶肉细胞分化相关的作用元件HD-Zip1、响应干旱胁迫的元件MBS,以及与玉米醇溶蛋白代谢调节相关的元件O2-site。
由图6可知,每个成员中都含有大量的启动子核心作用元件CAAT-box、TATA-box,除核心作用元件之外,每个成员都含有较多的光响应元件和激素响应元件,其中有14个成员(BBX3~6、BBX8、BBX12~14、BBX16、BBX18~21、BBX24)含有与厌氧诱导相关的元件,6个成员(BBX1、BBX5、BBX6、BBX9、BBX12、BBX14)含有与昼夜节律相关的元件,6个成员(BBX12、BBX16、BBX18、BBX21、BBX22、BBX25)含有响应低温的顺式作用元件。
2.6 葡萄BBX基因家族表达分析
通过对葡萄基因芯片数据的分析,得到25个葡萄BBX基因家族成员在53个葡萄组织中的基因表达谱。利用TBtools软件绘制基因表达图谱(图7),在图中用红色和蓝色表示基因表达的强度,红色表示信号强度较强,蓝色表示信号强度较弱。结果显示,BBX家族大部分基因表达强度较强,不同基因在葡萄不同器官、不同时期的表达存在差异。在进化树中处于同一分支的基因表达情况相似,例如,VvBBX12和VvBBX22基因在葡萄发育的不同时期表达量都很低;VvBBX6和VvBBX11基因在葡萄的整个生长发育期都处于高表达状态。同样在整个发育期都呈较高表达的还有VvBBX5、VvBBX7、VvBBX8、VvBBX9、VvBBX15、VvBBX23、VvBBX24、VvBBX25。此外,VvBBX24和VvBBX25基因在葡萄果实的果皮和果肉中都呈现高表达。
2.7 不同光质下葡萄BBX基因的表达
为探究BBX家族基因在光形态建成及光合作用方面的作用,对葡萄植株进行不同的光处理。由图8可知,黑暗处理下,VvBBX2、VvBBX6、VvBBX7、VvBBX13、VvBBX17、VvBBX21、VvBBX24基因表达量升高,其中VvBBX24基因的表达水平升高了5倍;蓝光照射下,VvBBX8、VvBBX13、VvBBX24基因表达量上调,其余基因与对照组相比,都表现不同程度的下调,其中VvBBX21基因在蓝光下表达量极低,相对表达量接近于0;远红光照射下,除VvBBX17外,其他基因均表现为下调;红光照射下,VvBBX2、VvBBX6、VvBBX9、VvBBX11、VvBBX21基因相对表达量与对照组相比表现为下调,其中VvBBX21基因相对表达量最低。BBX家族基因在不同光质照射下变化明显,说明BBX家族基因在光调节反应中有着重要的作用。
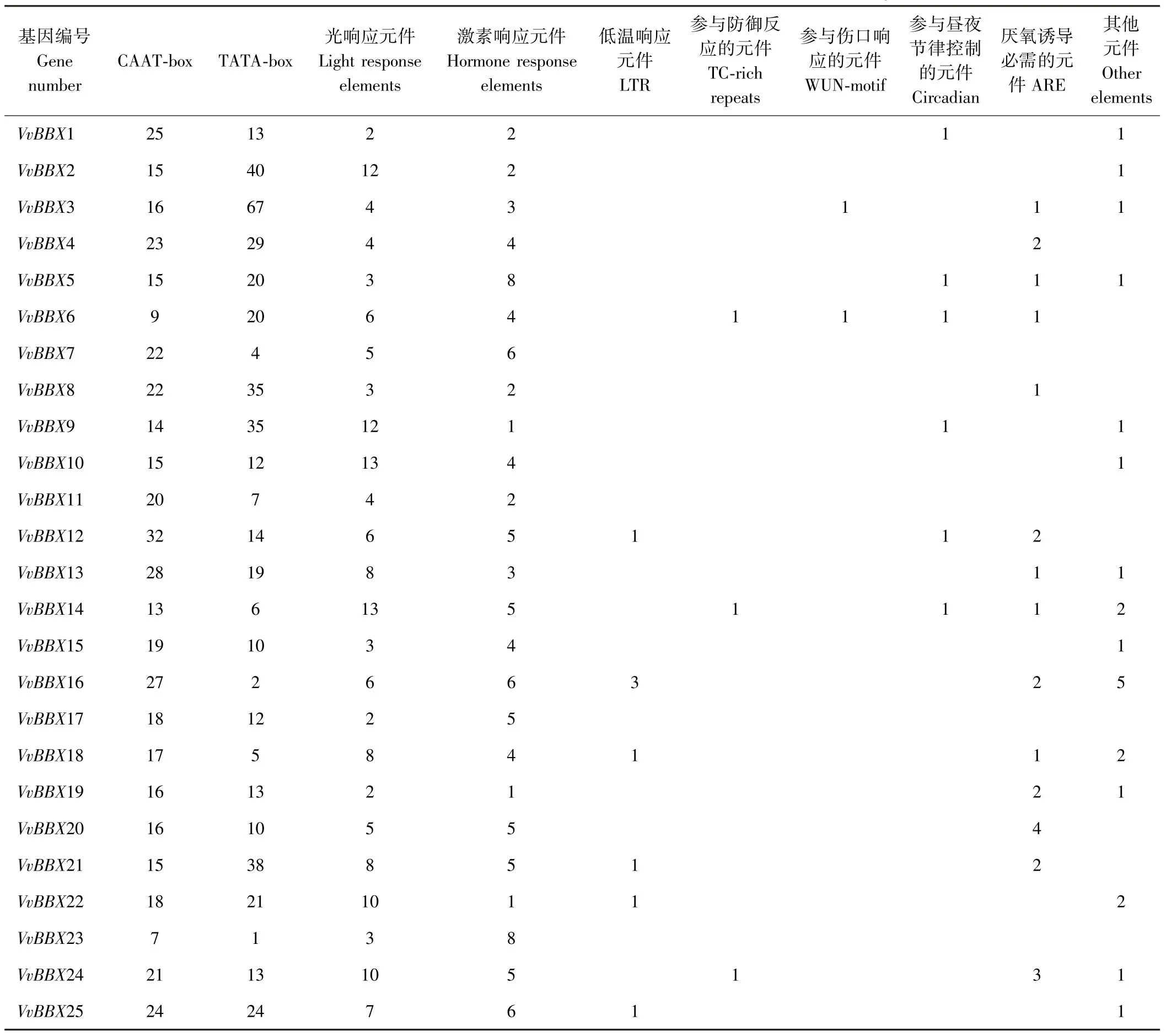
表3 葡萄BBX蛋白家族成员启动子顺式作用元件的分析Table3 Analysis of cis-acting elements in promoters of BBX protein family in grape
3 讨论
本研究利用生物信息学技术鉴定出25个葡萄BBX基因家族成员,这25个VvBBXs基因均存在Bbox结构域,部分成员在C端存在CCT结构。所有成员在染色体上所处的位置具有一定的差异,多数分布在染色体的两端,只有VvBBX12、VvBBX13、VvBBX14三个成员形成基因簇分布在12号染色体的中部。所有成员的理化性质存在一定的差异,25个成员中有4个成员的等电点大于7,为碱性氨基酸。通过基因家族多序列对比发现,它与拟南芥、水稻的BBX蛋白距离较近,在葡萄、拟南芥、水稻3个物种构建的进化树中,25个VvBBXs分别聚类到A、B、C、D四个分支中,聚类到同一分支中的基因功能可能存在相似性。对拟南芥和葡萄的基因共线性分析发现,葡萄基因组内一共存在4个基因对,葡萄与拟南芥基因组间有24个基因对,表明葡萄BBX蛋白在进化过程中,在染色体之间发生片段复制的概率更高,且在不同物种间BBX蛋白具有高度保守性。此外,本研究结果表明,葡萄BBX基因家族成员在葡萄的不同发育时期、不同组织的表达存在差异,例如VvBBX24、VvBBX25在芽、果实中的表达量较高,VvBBX8则在叶片中表达量较高;不同的家族成员在葡萄植株的整个发育期的表达量差异较大,BBX12、BBX19和BBX22在整个生长期表达量都在较低的水平。Bai等[25]研究指出,在苹果中BBX基因与MYB存在互作,促进了苹果果皮中花青素的积累。本研究发现了ACE、AE-box、Box-4等21种光响应元件,P-box、ABRE、TGA-element等8种激素类响应元件,还发现了低温响应元件LTR以及与昼夜节律控制相关的作用元件等。同时也发现了MYB结合位点,这表明葡萄BBX基因与果实的着色相关,其功能还有待进一步研究。
BBX蛋白参与植物的光形态建成过程,为了探究葡萄VvBBX基因在光形态建成中的作用,本研究对葡萄植株进行不同光照条件处理,对随机挑选的10个来自不同组的BBX基因进行表达量分析,结果显示,不同的光照条件下,10个基因都发生了明显的变化。在黑暗处理中,有7个基因表达量升高,这些基因很有可能参与到葡萄植株昼夜节律的调控;在蓝光、红光以及远红光照射下,基因都呈现不同程度的变化。红光和蓝光照射下BBX21基因几乎不表达,而BBX8基因在蓝光和红光照射下表达量增高,这些变化说明BBX基因家族在光调节途径中存在复杂的调控机制,还需进一步验证。光质对果树形态建成、生长发育及果实品质具有重要的调控作用,且对种子萌发,根、茎、叶的生长,叶片的衰老,基因表达以及次生代谢等产生影响[26-29]。研究表明,红光促进植物叶片伸展和胚轴伸长,提高净光合速率,影响同化产物向营养器官的分配,促进植株的生长等;蓝光抑制胚轴伸长,增加茎粗,提高生物量,增加类胡萝卜素含量,提前开花[30]。对在不同光质下的葡萄BBX基因表达进行深入研究将有助于在葡萄生产上实现提质增效的目标。
4 结论
基于葡萄全基因组数据,本研究在葡萄中共鉴定出25个BBX家族基因,将其命名为VvBBX1~VvBBX25。对所有成员进行生物信息学分析发现,25个成员分布在11条染色体上,其中包括4个分泌通路信号肽、2个叶绿体转运肽和1个线粒体靶向肽;BBX基因家族的进化分析表明其具有很强的保守性;不同成员在葡萄不同组织中表达存在差异,在不同发育时期不同家族成员的表达也存在差异;葡萄BBX基因家族启动子区域具有丰富的光响应元件和激素类响应元件,表明BBX基因家族是光调节反应和植物生长发育的重要因子。本研究结果将为葡萄着色机理、葡萄的光调节反应以及葡萄的生长发育研究提供新的思路、新的方向。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
科学之谜(2019年3期)2019-03-28 10:29:44
科学之谜(2018年8期)2018-09-29 11:06:46
生命科学研究(2018年1期)2018-05-29 01:12:47
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
现代工业经济和信息化(2016年2期)2016-05-17 05:34:16
中央民族大学学报(自然科学版)(2015年2期)2015-06-09 08:45:16
电子工业专用设备(2015年4期)2015-05-26 09:10:40
