
毛竹4-香豆酸辅酶A连接酶基因家族鉴定及表达分析
2021-01-28 02:24:26杨克彬单雪萌史晶晶朱成磊高志民
核农学报 2021年1期
杨克彬 单雪萌 史晶晶 朱成磊 高志民
(国际竹藤中心竹藤资源基因科学与基因产业化研究所,国家林业和草原局/北京市竹藤科学与技术重点开放实验室,北京 100102)
丙烷类代谢初始途径是以苯丙氨酸为原料,由苯丙氨酸解氨酶(phenylalnine ammonialyase,PAL)、肉桂酸4-羟化酶(cinnamic acid 4-hydroxylase,C4H)和4-香豆酸酯:辅酶a连接酶(4-coumarate:CoA ligase,4CL)3个酶介导催化的复杂的生化反应,并合成一系列植物所必需的物质,如木质素单体、植物激素、类黄酮和苯丙烯类等[1-2]。4CL作为该途径的最后一种酶,催化几种羟基肉桂酸转化为相应的辅酶A(coenzyme A,CoA)酯,从而控制碳通过苯丙烷代谢途径进入特定生物合成途径[3-4]。木质素作为丙烷类代谢途径的重要产物之一,是次生壁的主要成分,其在细胞壁的结构完整性、水分运输和保护植物免受病原体感染等方面发挥着关键作用。在4CL氨基酸序列中存在2个高度保守的基序,即SSGTTGLPKGV和GEICIRG,负责识别、结合和催化底物形成相应的CoA酯[5]。根据参与物质合成途径的不同,可将4CL分为两大类,其中Ⅰ类参与木质素单体的合成途径,而Ⅱ类主要参与黄酮类化合物合成的酶促反应[6-7]。在拟南芥中,Ⅰ类的At4CL1、At4CL2和At4CL4编码的同工酶参与木质素生物合成,而At4CL3激活p-香豆酸作为查耳酮合成酶的底物,负责类黄酮代谢[8-9];大豆Gm4CL3/4基因调控类黄酮的生物合成,且在根和上胚轴的表达较高[10]。
4CL作为木质素生物合成过程中关键的限速酶,已在数十种植物中分离并鉴定,如拟南芥[11]、小立碗藓[12]、毛果杨[13]水稻[14]和柑橘[15]等。竹子作为我国最具特色的非木质林产品资源,在当下木材资源短缺的情况下,竹材的优良材性使其成为木材潜在的重要替代品,倍受关注。竹材的材性与其木质素含量密不可分,研究人员已经从木质素的生物合成途径角度开展研究,包括参与竹子木质素生物合成的CoCOMT[16]、PePAL1[17]、C4H[18]和PeLAC[19]等基因。木质素生物合成的每个基因家族都包括多个成员,然而对于竹子中4CL基因家族的了解并不全面。因此,本研究以毛竹(Phyllostachysedulis)为对象,在全基因组水平筛选与鉴定4CL基因的基础上,系统分析其基因结构、蛋白保守结构域和进化关系,并通过毛竹转录组数据构建的基因表达谱测序(RNA-seq)分析4CL基因的组织表达特异性,结合笋木质素组织化学染色和实时荧光定量PCR(quantitative real-time PCR,qPCR)技术,研究随着竹笋高度的增加,其木质化程度和4CL基因的表达模式,以期为揭示4CL在竹子木质素生物合成和木质化中的功能提供参考。
1 材料与方法
1.1 试验材料
以江西南昌(28°45′58″N,115°45′39″E,海拔399.0 m)野外生长良好的毛竹作为试验材料,分别采集不同生长阶段竹笋(0.2、1.0、3.0和6.7 m)基部为样品。样品经液氮处理后,存储于-80℃冰箱中,用于提取RNA。此外,取相同样品固定在标准固定液(formalinacetic acid alcohol,FAA)中保存于4℃冰箱,用于树脂切片。
1.2 试验方法
1.2.1 毛竹4CL基因的鉴定 以模式植物拟南芥(Arabidopsisthaliana)和水稻(Oryzasativa)中4CL基因作为种子序列,在毛竹基因组数据库进行BLAST获取其中4CL的同源基因候选序列。运用BLASTN和BLASTP进一步对候选基因逐条比对分析,并对候选基因编码的氨基酸序列进行保守结构域完整性鉴定。最终获得毛竹4CL家族基因成员,并编号。采用在线工具ProtParam(http://web.expasy.org/protparam/)分析毛竹4CL基因编码蛋白的基本理化性质。利用Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/#)软件进行蛋白亚细胞定位预测。
1.2.2 毛竹4CL基因的生物信息学分析 运用GSDS(http://gsds.cbi.pku.edu.cn/)对毛竹4CL基因的结构,即内含子/外显子结构进行分析[20];运用在线软件MEME(http://meme.nbcr.-net/meme/intro.html)对毛竹4CL的保守蛋白基序进行分析,软件参数:motifs最大数量10;长度为6~300,其他参数为默认值。运用DNAMAN软件对毛竹4CL蛋白氨基酸序列进行多重序列比对后进行保守结构分析,并运用在线软件Pfam(ttps://www.ebi.ac.uk/Tools/hmmer/)和WEBLOGO(http://weblogo.berkeley.edu/logo.cgi)对其保守结构域进行功能注释并构建Seqlogo图[21-22]。
为了探索毛竹4CL的进化关系,利用毛竹、水稻、二穗短柄草(Brachypodiumdistachyon)、拟南芥、大豆(Glycinemax)和毛果杨(Populustrichocarpa)等植物的4CL氨基酸序列,通过MEGA 7.0内置的ClustalW程序对4CL家族成员的氨基酸序列进行多重序列比对,并采用邻接法(neighbor-joining method,NJ)构建系统进化树(使用系统默认值),校验参数Bootstrap值设置为重复1 000次。其中,水稻、二穗短柄草、拟南芥、大豆和毛果杨的4CL氨基酸序列分别从Rice genome annotation database、Arabidopsisgenome TAIR release 10.0和Phytozome数据库下载[23]。
1.2.3 毛竹4CL基因的组织特异性表达分析 从NCBIShort Read Archive(SRA)下载毛竹不同发育阶段的鞭、根、笋、叶片、鞘和芽的转录组数据[24],获取其中4CL基因的FPKM(每千个碱基的转录每百万映射读取的碎片值,fragments per kilobase of exon model per million fragments)值,用以表示基因的表达丰度。为方便统计,对每个表达数值取以2为底数的对数(Log2),使用 Matrix2png(http://www.chibi.ubc.ca/matrix2png/)绘制基因表达热图。
1.2.4 PEG树脂切片分析 参照Yang等[25]的试验方法对FAA固定液中的竹笋样品进行包埋处理,经过不同浓度的聚乙二醇(polyethylene glycol,PEG)逐级脱水后,包埋于PEG 4000中。用Leica RM2165旋转切片机(Leica,德国)进行切片,厚度为10μm;用0.05%甲苯胺蓝(toluidine blue O,TBO)对展片的切片于80℃染色3 min,并拍照观察。
1.2.5 毛竹4CL基因在笋不同发育时期中的表达模式分析 利用Primer Premier 5.0软件设计毛竹4CL家族基因特异性引物,由上海生工生物合成工程技术服务公司合成(表1)。采用Trizol(Promega,美国)法提取毛竹各样品的总RNA,用DNaseⅠ(TaKaRa,日本)除去基因组DNA后,使用反转录试剂盒(Promega,美国)合成cDNA第一链,操作参照相关试剂盒说明书进行。试验在qTOWER荧光定量PCR仪(耶拿,德国)进行,反应体系(10μL):2×SYBRⅡGreen 1 Master 5.0 μL、正向和反向引物各0.2μL(10μmol·L-1)、cDNA模板0.8μL、ddH2O 3.8μL。两步法进行PCR,扩增程序:95℃预变性6 min;95℃变性10 s,62℃退火10 s,45个循环。以PeNTB为内参基因[26],采用2-ΔΔCT法分析基因的相对表达量[27]。
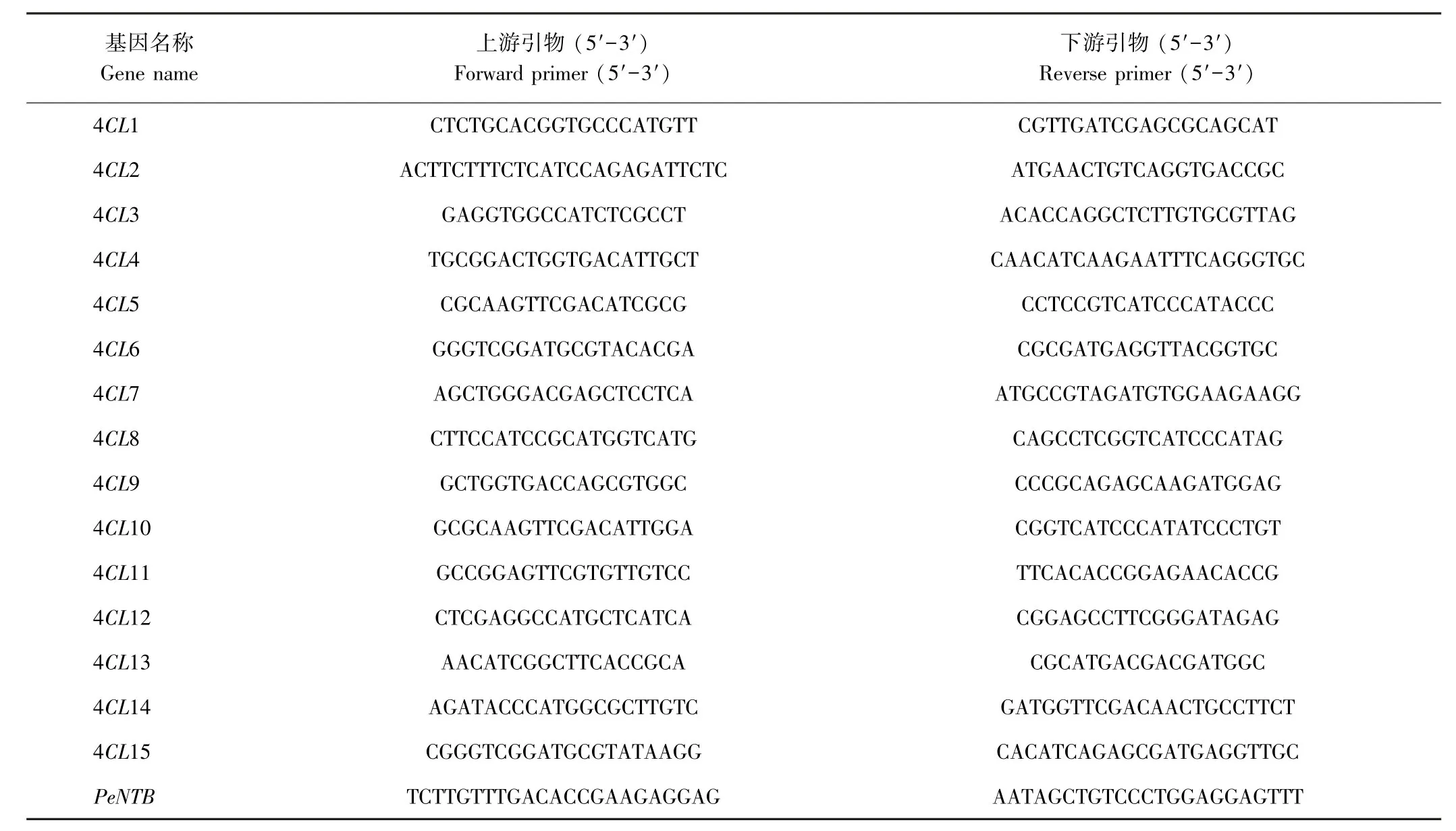
表1 qPCR所用引物Table1 Primers used in qPCR
2 结果与分析
2.1 毛竹4CL家族成员鉴定与蛋白理化性质分析
利用拟南芥和水稻同源基因序列,从毛竹基因组数据库中经BLAST分析,共鉴定获得15个4CL家族成员,均具有完整的保守结构域,依次命名为Pe4CL1~Pe4CL15。通过ProtParam工具,对Pe4CLs编码蛋白质的长度、分子量及等电点等理化性质进行分析,结果显示,Pe4CLs的氨基酸长度为520~671 aa,对应分子量为55 044.15~72 445.28 Da;Pe4CLs的蛋白不稳定系数为33.41~51.43,其中7个成员的不稳定系数小于40属于稳定蛋白,其余为不稳定蛋白;平均亲水系数为-0.135~0.207,等电点为5.00~8.72,且除Pe4CL4和Pe4CL14外,其余氨基酸的等电点均小于7.00;经过亚细胞定位预测,15个Pe4CLs均被定位在过氧物酶体上(表2)。
2.2 Pe4CLs基因结构及其编码氨基酸的保守基序分析
基因结构分析表明,15个Pe4CLs均具有内含子,数量为2~5个;大部分成员含有5个或6个外显子(图1)。Pe4CLs的基因结构差异明显,结构具有多样性,具体表现在内含子的大小和位置存在较大差异。
根据序列的相似性和系统进化,毛竹的4CL基因家族被分为2个亚家族(A和B),分别含有7个和8个Pe4CLs成员(图2),通过MEME在线软件分析Pe4CLs中的保守基序,研究其中基序组成的多样性与保守性。由图2可知,在Pe4CLs中共鉴定出10个保守基序,命名为Motif 1~Motif 10。Motif 1~3、Motif 5和Motif 7为15个Pe4CLs所共有,其中,Motif 1和Motif 5分别包含4CL特征保守域SSGTTGLPKGV和GEICIRG,而Motif 10只存在于亚家族A中。大多数同亚类的成员具有相同的Motif组成,例如亚家族A中各成员均含有全部的10个基序,表明这些成员可能具有相似蛋白功能。
2.3 Pe4CLs保守结构域分析
为了研究Pe4CLs的保守结构域序列特征,对其氨基酸序列进行多重序列比对分析其保守结构域,结果显示,从蛋白质的N端到C端,依次发现了以下5个保守结构域:BoxⅠ(SSGTTGLPKGV)、BoxⅡ(QGYGMTE)、Box Ⅲ (GEICIRG)、Box Ⅳ(GWLHTGD)和BoxⅤ(VDRLKELIK)(图3)。BoxⅠ和BoxⅢ为2个已知的4CL特征保守域,负责识别、结合和催化反应底物,在毛竹中具有很高的保守性,但也存在差异位点,如BoxⅢ4号位的半胱氨酸(Cys)是最保守的氨基酸位点,直接参与酶催化反应,而在毛竹中少数Pe4CLs变为了色氨酸(Trp)和苯丙氨酸(Phe)。虽然BoxⅡ、BoxⅣ和BoxⅤ各自的功能尚不明确,但是通过在线软件Pfam分析保守结构域发现BoxⅠ~BoxⅤ共同组成了腺苷-1-磷酸(adenosine 5′-monophephate,AMP)结合结构域,行使酰基CoA合成酶功能,通过产生AMP参与到苯丙烷代谢途径中。

表2 Pe4CLs蛋白的理化性质和亚细胞定位预测Table2 Putative basic physical and chemical characteristics,and subcellular localization of Pe4CLs
2.4 系统进化分析
为了确定Pe4CLs与其他物种已知的4CL蛋白之间的进化关系,对毛竹、水稻、二穗短柄草、拟南芥、大豆和毛果杨的4CL氨基酸序列进行了多重比对,并利用MEGA 7.0软件构建了系统发育树。结果显示,6个物种的4CL成员被分成两个分支(A和B),且分支A中涵盖6个物种的4CL成员,而分支B只包含毛竹的8个成员(Pe4CL1/2/4/6/7/11/14/15)(图4)。进一步分析表明,分支A中的4CL又分为3个类型,即TypeⅠ~TypeⅢ[14],其中TypeⅠ仅包含一个毛竹成员(Pe4CL3),与Os4CL2聚类在一起;TypeⅡ中不包含毛竹成员,全都来自双子叶植物;TypeⅢ全是来自单子叶植物毛竹、水稻和二穗短柄草的4CL,且毛竹与水稻的距离较近,而与拟南芥、大豆和毛果杨的距离较远。
2.5 Pe4CLs组织特异性分析
由图5可知,15个Pe4CLs在毛竹不同组织和不同生长时期有着不同程度的表达,表达丰度存在明显差异。7个基因(Pe4CL1/2/5/8/10/13/15)在各组织中均检测到表达,它们可能作为组成型基因在毛竹整个生长发育过程中发挥重要作用,其中Pe4CL8、Pe4CL10和Pe4CL15在各个组织中相对表达量均较高,其余8个基因具有组织特异性,即只在特定组织中检测到表达且相对表达量较高,如Pe4C3在发育成熟的根和叶片中较其他组织具有较高的相对表达量。在不同生长阶段的笋中,大多数Pe4CLs的相对表达量具有相同的变化趋势,即随着竹笋高度的增加呈先升高后降低的趋势,且在1.0 m或3.0 m处达到峰值;同一基因在同一高度竹笋不同部位的相对表达量则表现为中下部高于尖部。此外,Pe4CL2在0.2 m竹笋中的相对表达量高于其他高度,且尖部的相对表达量高于中下部。
2.6 Pe4CLs在不同高度笋中的表达模式分析
毛竹笋切片TBO染色结果表明,随着竹笋高度的增加,维管束的染色加深区域明显逐渐扩大,说明其木质化逐渐增强,尤其是后生木质部导管周围的厚壁细胞木质化程度最高(图6)。
由图7可知,12个Pe4CLs在不同生长高度竹笋中均呈现差异表达,随着竹笋高度的增加,除了Pe4CL6的表达下调外,其他基因均不同程度地上调,且具有两种变化趋势。具体表现为Pe4CL3和Pe4CL15的相对表达量持续上调,在6.7 m竹笋中Pe4CL3和Pe4CL15的相对表达量分别是0.2 m笋中的82倍和3倍;其他9个基因(Pe4CL1/2/5/8/9/10/11/13/14)表现出类似的表达趋势,即先升高后降低,峰值出现在1.0 m或3.0 m竹笋中,且它们在峰值处的相对表达量均较0.2 m竹笋均上调10倍以上,其中Pe4CL9的最高,上调幅度达到3 862倍。然而,这9个基因在6.7 m竹笋中的表达量均较峰值明显下降,幅度超过85%。Pe4CL6的相对表达量呈现完全不同的趋势,即与0.2m竹笋中的相对表达量相比,该基因在1.0 m竹笋中的表达量急剧下调(约为0.2 m竹笋的2%),并在之后不同高度的竹笋中保持相对稳定的低水平。此外,本研究还对另外3个基因(Pe4CL4/7/12)进行了定量分析,但由于设计不出特异引物而未获得理想的结果。
3 讨论
木质素含量对竹材的机械强度起着决定性作用,4CL是木质素生物合成的关键酶。因此,开展毛竹4CL基因的结构特征和表达模式分析,对于揭示其在竹材形成过程中的作用具有重要的参考价值。本研究从毛竹中鉴定出15个具有完整结构域的4CL家族成员,而在双子叶植物拟南芥和杨树中,或在单子叶植物水稻和二穗短柄草中,4CL成员均不多于5个,这可能与毛竹进化过程中经历了染色体加倍事件有关[28]。进一步分析发现,BoxⅠ和BoxⅢ作为底物结合区域的绝对保守结构域,在Pe4CLs中某些位点并不保守,说明它们进化过程中功能发生了分化,可识别与结合不同的反应底物,尤其是BoxⅢ的Cys残基的突变可能造成它们功能弱化或者失去催化羟基肉桂酸的功能,这与杨树和水稻等物种中的4CL类似[10,29-30]。
本研究中,系统进化分析发现,A亚家族中所有Pe4CLs(7个)均与水稻的4CL聚类到较近的分支,分布在TypeⅠ和TypeⅢ中,而与双子叶的4CL成员距离较远。对Os4CL1/3/4/5体外表达重组蛋白活性研究表明,4-香豆酸和阿魏酸是水稻木质素单体生物合成的主要底物[14,31],Os4CL2则参与类黄酮的形成[31]。随着水稻茎秆的生长,其中木质素含量增加,木质化程度加强,Os4CL1/2/3/4/5呈现不同的表达模式,但转录水平Os4CL3>Os4CL5>Os4CL1>Os4CL4>Os4CL2,其中抑制Os4CL3的表达,水稻木质素含量显著降低[14]。TypeⅢ中Pe4CL5和Pe4CL10与Os4CL3、Pe4CL8与Os4CL4、Pe4CL9和Pe4CL12与Os4CL5、Pe4CL13与Os4CL1分别聚类到较近的分支,推测毛竹4CL可能与对应的水稻4CL具有相似的功能,参与竹子木质素生物合成;而Pe4CL3在TypeⅠ中与Os4CL2聚类到一起,Pe4CL3可能具有参与类黄酮形成的功能。
基因的表达模式是其生物学功能的具体表现形式之一。本研究转录组表达谱热图结果显示,7个基因(Pe4CL1/2/5/8/10/13/15)在各组织中均检测到表达,表明这些基因不仅参与竹笋成熟过程中木质化进程,同时也可能参与到其他组织的生长过程中,尤其是Pe4CL8、Pe4CL10和Pe4CL15在各个组织中的高表达说明它们对这些组织的形成均具有重要作用,但是否直接参与竹秆成熟过程的木质化,需要进一步验证。其他8个基因如Pe4CL12在箨片中、Pe4CL14在叶片中特异表达,而Pe4CL4在叶片、叶鞘和箨片均表达较高,说明它们可能参与特定组织细胞壁的木质化过程。此外,现有转录组结果中毛竹根系中尚不存在明显的与木质素含量相关的Pe4CLs表达模式。
本研究中,随着竹笋高度的增加,木质化程度也逐渐加强,这与本试验前期研究结果一致[20]。同时,切片结果显示随着竹笋高度的增加,木质化的速度减缓,3.0 m和6.7 m竹笋纤维束的染色范围和深浅变化不明显。随竹笋高度的增加,11个Pe4CLs的转录水平显著上调,且有9个基因的相对表达量呈现先升高后下降的变化趋势,这与切片显示的笋木质化程度变化相一致,其原因可能是6.7 m竹笋较4.0 m竹笋中基因表达量下降导致其木质化程度减缓。而Pe4CL3和Pe4CL15的相对表达量持续增加,说明这两个基因对笋木质化的作用持续增强。以上结果说明,Pe4CLs通过表达参与到毛竹木质素的生物合成中,基因上调的表达模式表明它们可能是木质素合成的正向调控因子,这与前人对模式植物的研究结果相一致[29,32-33]。B亚家族中Pe4CL6的表达随着竹笋高度的增加而下调,可能该基因只在幼笋中发挥作用,但其具体功能尚需验证。由此表明,大部分Pe4CLs可能在毛竹的苯丙氨酸代谢途径中控制着底物是否流向木质素,在木质素的合成过程中发挥着重要作用;而有3个基因的表达未被检测到,1个基因表达下调,它们可能具有新的功能。此外,qPCR检测的基因表达模式与转录组数据存在一定的差异,这可能是由于所取样品差异造成的。
4 结论
本研究基于基因组与转录组数据,利用生物信息学方法从毛竹中鉴定了15个4CL成员,根据进化关系将它们分为两个亚家族。转录组表达谱分析揭示出不同Pe4CLs在毛竹不同组织与不同发育时期的表达存在明显差异。竹笋组织切片染色和qPCR结果证实,随着竹笋高度的增加,木质化程度加深,大多数Pe4CLs的相对表达量上调,与笋的木质化程度相一致,表明它们参与木质素的生物合成。本研究结果为进一步揭示Pe4CLs参与竹材形成的分子机制提供了参考依据。
猜你喜欢
意林·少年版(2020年13期)2020-08-02 11:02:50
创新作文(小学版)(2018年7期)2018-08-20 03:11:02
小天使·一年级语数英综合(2018年4期)2018-06-22 10:33:26
小天使·一年级语数英综合(2017年11期)2017-12-05 07:18:15
小布老虎(2017年4期)2017-08-10 08:22:40
小学生作文(低年级适用)(2017年3期)2017-07-06 12:06:53
国际木业(2016年8期)2017-01-15 13:55:23
国际木业(2016年3期)2016-12-01 05:04:53
国际木业(2016年1期)2016-12-01 05:04:11
福建农业科技(2015年1期)2015-02-27 10:20:39
