
基于线粒体基因的云南省烟田烟蚜茧蜂的种群遗传多样性研究
2021-01-28 01:37冯增贝张翠萍李红娟杜振勇周文兵
云南民族大学学报(自然科学版) 2021年1期
冯增贝,张翠萍,李红娟,杜振勇,李 虎,周文兵
(1 中国农业大学 植物保护学院昆虫学系,北京 100193;2 云南烟草公司,玉溪市分公司,玉溪 653100)
蚜虫是一类重要的农业害虫,其繁殖率高、生命周期短、防治难度大.长期以来使用化学农药防治,使蚜虫对大多数农药产生了一定的抗药性[1].此外,过度使用化学农药还带来许多不利后果,如农药残留和农业面源污染等,严重影响生态环境、农业生产、农产品的质量和安全,威胁人类健康和农业可持续发展[2].生物防治技术利用了生物物种间的相互关系,可以通过一次性或多次引入天敌来抑制害虫,达到良好的防治效果,大大降低化学农药的使用[3-4].烟蚜(Myzuspersicae)亦称桃蚜,是烟草上的常发性害虫和重点防控对象,寄主包括30科400多种植物[5-8],能够传播100多种病毒[9-10],烟蚜是在烟草整个生长期均有发生,通过刺吸取食烟叶,传播烟草病原物,造成烟叶的质量和产量下降.
烟蚜茧蜂(Aphidiusgifuensisashmaed)是烟蚜优势内寄生蜂[5,12-15],自然寄生率达30%~60%[14],对烟蚜有较强的控制作用[15].具有产卵量大、寿命长、适应性强的特点,能长时间持续控制蚜虫危害,在烟蚜的生物防治中具有十分广阔的应用前景[8].
我国针对烟蚜茧蜂的研究开始于20世纪80年代,主要集中在其生物学、生态学及生物防治利用等方面.赵万源等[5]曾在云南玉溪烟田对烟蚜茧蜂开展生物学及应用的初步研究,结果表明利用烟蚜茧蜂防治烟蚜的想法是有可能实现的.随后,汤玉清等[16]、忻亦芬[17]、毕章宝等[18-19]分别在烟蚜茧蜂的生物学、生态学等方面开展了相关研究.2007年,云南烟区开始规模化扩繁和释放烟蚜茧蜂进行烟蚜的田间防治,并取得了良好的应用效果;经过十多年的研究和应用,云南省主要烟区烟蚜茧蜂繁育和释放的设施与技术均相对成熟.近几年,该技术逐渐开始在在全国范围内推广应用[20],目前云南、贵州、广西、吉林、山东、湖南等国内主要烟区都实现了烟蚜茧蜂的大规模扩繁和产品的开发,并投入到实际生产中[20-25].虽然烟蚜茧蜂具有广阔的应用前景和良好的防治效果,但是在其工厂化扩繁和人工释放等过程中仍存在许多问题,尤其是蜂群的退化问题[26].工厂化扩繁的烟蚜茧蜂存在长期近亲繁殖、不当引种、饲养环境不良等因素的影响[27],造成蜂群遗传多样性下降和种群的衰退,释放到田间后适应环境的能力下降,表现出生殖力衰退、寄生效果下降、整体种群竞争力减弱等问题而难以定殖.
目前,关于云南省主要烟田烟蚜茧蜂种群的遗传多样性和遗传结构的特点,以及长期的大规模人工扩繁和田间释放是否对其种群遗传多样性产生了影响等问题尚不明确,相关研究缺乏.本研究利用atp6、cox1、cob和nad2 4个线粒体基因的全序列作为分子标记,对云南省11州市19个样点的烟田和工厂繁育种群以及1个野生种群的种群遗传多样性进行初步研究,为该寄生蜂的科学利用和蜂种合理复壮提供参考.
1 材料与方法
1.1 种群样品的收集
本研究的样品于2018—2019年在云南省11市州的烟田、繁蜂工厂和野外蔬菜田采集获得,其中烟田种群(S)6个,工厂繁育种群(G)13个,野生种群(Q)1个(表1),样点的选择覆盖了云南主要植烟区域.野外烟蚜茧蜂种群的采集主要是通过采集被寄生的僵蚜,并置于网兜内直到烟蚜茧蜂羽化,立即将其浸泡在无水乙醇中,每个僵蚜个体采集距离不低于5 m,回到实验室之后,更换一次无水乙醇,置于-20 ℃冰箱内保存.标本经过形态和DNA条形码鉴定后,进行后续试验.
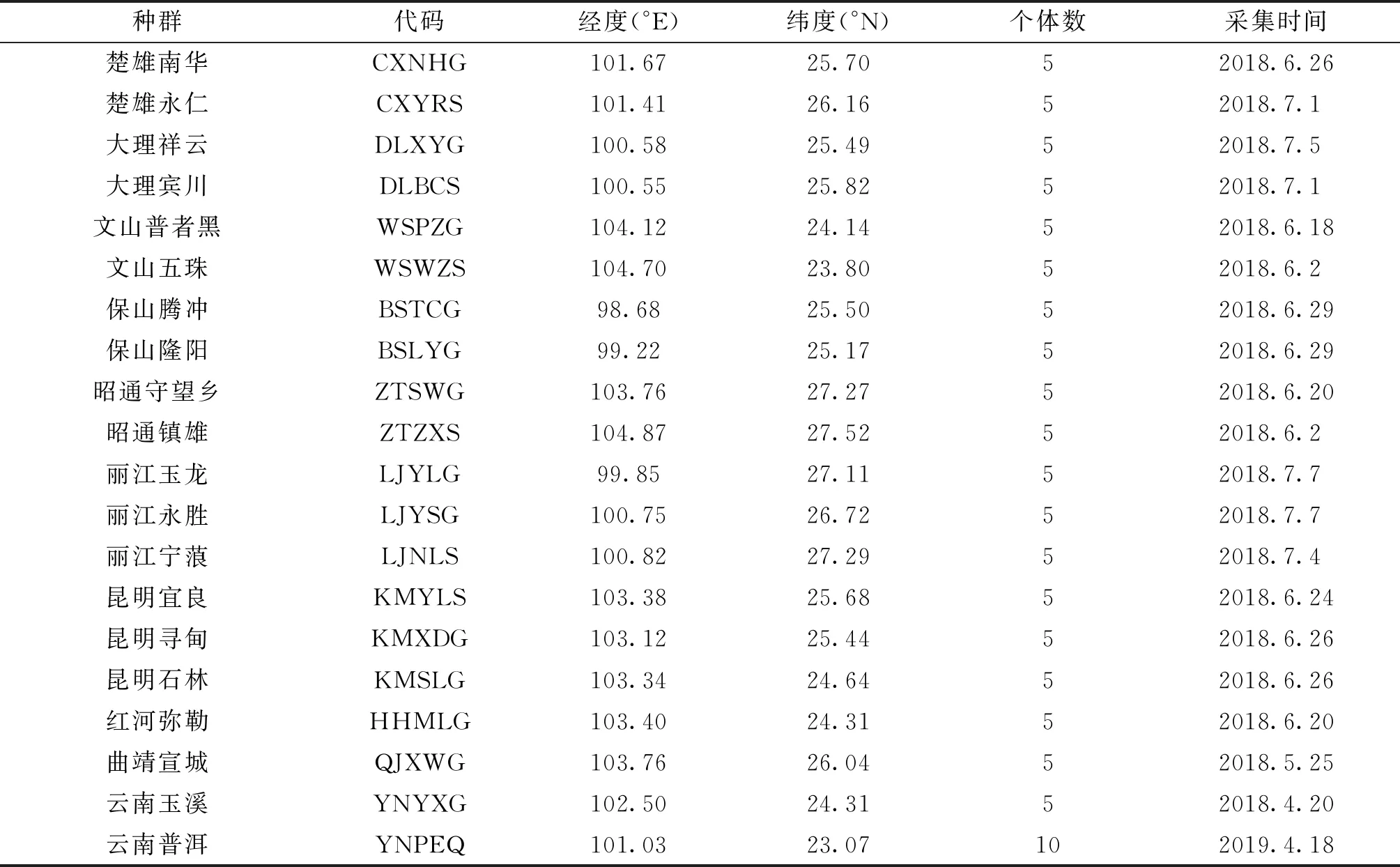
表1 云南省主要烟区烟蚜茧蜂种群样本的采集信息
1.2 基因组DNA提取和线粒体基因数据的获取
将雌成蜂单头样本置于 2 mL 研磨管中,加入1大1小2个灭菌后的研磨珠,使用研磨仪(上海净信实业发展有限公司,tissuelyser-32)进行研磨,最后使用天根血液/细胞/组织/基因组DNA提取试剂盒(DP304)进行DNA的提取.基因组DNA构建插入片段长度为350 bp的2代测序文库,利用Illumina NovaSeq 6000(北京贝瑞和康生物技术有限公司)平台进行150 bp双端测序.原始测序数据下机后经过质量控制获得clean data.以本课题组测序完成的烟蚜茧蜂线粒体基因组作为参考序列(GenBank登录号:MT264907),选取atp6、cox1、cob和nad2基因序列在测序数据中调取目的片段,利用Geneious 10.1.3(http://www.geneious.com)软件“map to the reference”操作,参数设置为:最小重叠40 bp,最小序列相似度95%.利用MAFFT[28]软件对所有个体的线粒体基因分别进行同源比对,最后串联为1个数据集用于种群遗传多样性的分析.
1.3 种群遗传学分析
利用DnaSP5.10[29]软件计算单倍型数(number of haplotypes, Ht)、单倍型多样性(haplotype diversity, Hd)、核苷酸多样性(nucleotide diversity, Pi)、平均核苷酸差异数(average number of nucleotide differences, K)、多态性位点数(the number of polymorphic sites, S)等种群遗传多样性参数.利用Arlequin3.5[30]软件计算不同分组变异分布及分化系数(AMOVA)和种群遗传分化指数(Fst).通过PopART1.7[31]软件的中值邻接法构建单倍型进化网络,利用IQTREE[32]软件构建系统发育树,碱基替代模型为自动预测,并进行1000次自展检验.
2 结果与分析
2.1 烟蚜茧蜂的种群遗传多样性
烟蚜茧蜂4个线粒体蛋白质编码基因串联后的总长度为4,359 bp(GenBank登录号:MT653697~654116).云南省20个种群105个个体中,共有10种单倍型,单倍型多样性0.501、核苷酸多样性0.00743、平均核苷酸差异数32.385、多态位点数158.总的来看,核苷酸多样性最高的是工厂繁育种群,最低的是烟田种群;而单倍型多样性最高的是普洱野生种群,最低的是烟田种群(表2).
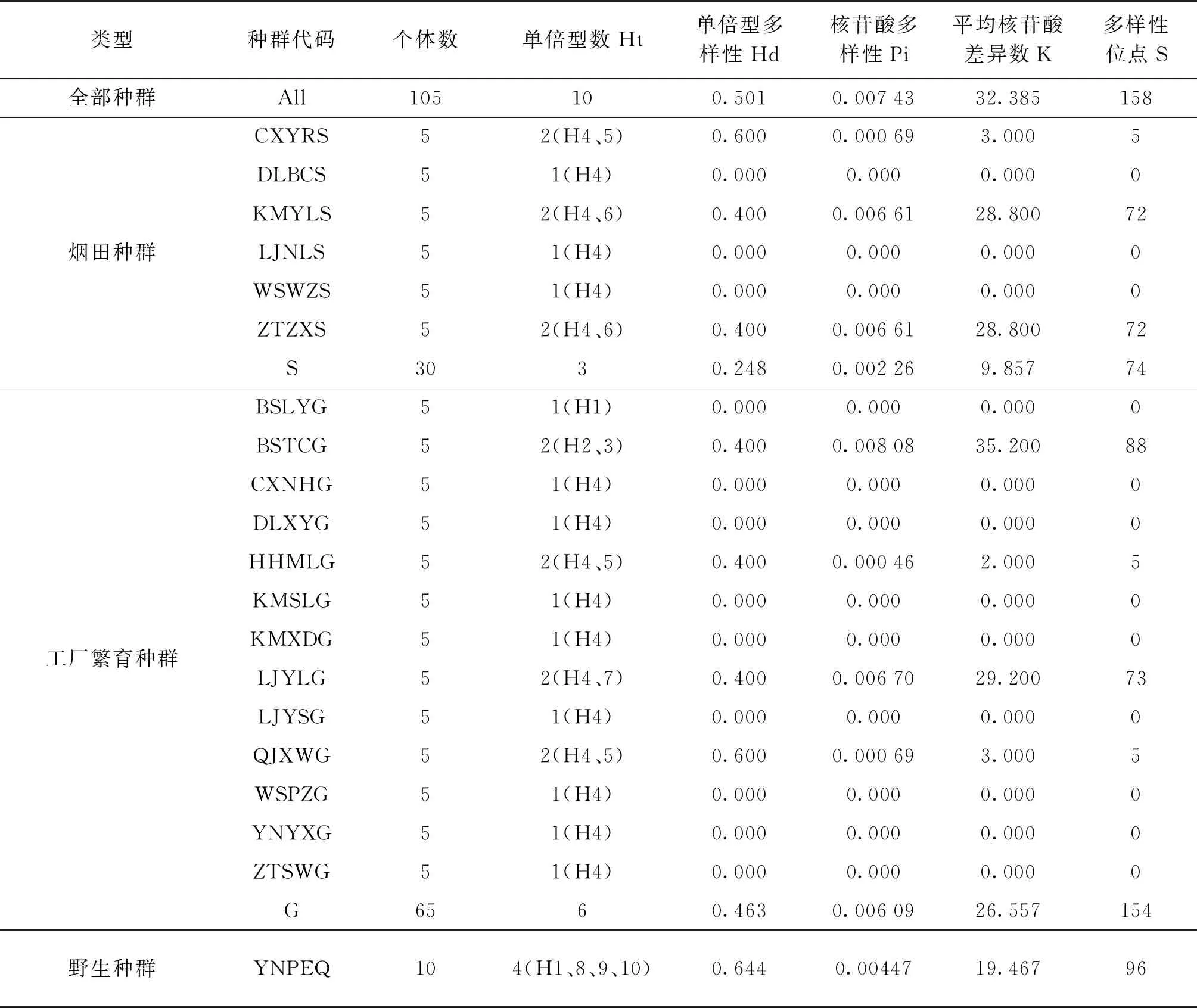
表2 云南省主要烟区烟蚜茧蜂种群遗传多样性参数
在6个烟田种群中,有3个没有遗传多样性,另外3个种群核苷酸多样性为0.000 69~0.006 61,昭通镇雄(ZTZXS)和昆明宜良(KMYLS)种群核苷酸多样性最高;在13个工厂繁育种群中,有9个种群没有遗传多样性,保山腾冲(BSTCG)和丽江玉龙(LJYLG)种群具有较高的核苷酸多样性,曲靖宣城(QJXWG)和红河弥勒(HHMLG)种群核苷酸多样性较低.整体上看,云南省主要烟区烟田和工厂繁育种群总体的遗传多样性偏低.
10种单倍型中,4个为共享单倍型,6个为独有单倍型(图1).烟田种群只有3种单倍型,均为共享单倍型,其中Hap6为烟田种群所独有.工厂繁育种群有6种单倍型,其中3个为独有单倍型(Hap2、Hap3、Hap7).烟田种群中楚雄永仁(CXYZS)、昆明宜良(KMYLS)、昭通镇雄(ZTZXS)均有2种单倍型(Hap4和Hap5,Hap4和Hap6),而同一州市的工厂繁育种群只有1种单倍型(Hap4).Hap4是11个种群的唯一单倍型,只有保山腾冲(BSTCG)、保山隆阳(BSLYG)和云南普洱(YNPEQ)种群中没有出现.而保山腾冲(BSTCG)(Hap2、Hap3)和云南普洱(YNPEQ)(Hap8、Hap9、Hap10)2个种群中就有5种独有单倍型,保山隆阳(BSLYG)和云南普洱(YNPEQ)种群共享Hap1.
从单倍型数量和频率来看,Hap4数量最多,共有73个个体,占所有样本的69.5%,在17个种群均有出现,包含了所有的烟田种群和多数工厂繁育种群.其中Hap4在烟田种群中出现26次,占所有烟田样本的86.7%,在工厂繁育种群出现47次,占所有工厂繁育样本72.3%.
单倍型网络图(图1)和系统发育树(图2)的结果一致,大致分为5支,各分支差异较大.云南普洱和保山腾冲种群的单倍型出现在关系较远的两个分支上,保山2个工厂繁育种群的单倍型均与其它工厂繁育种群的单倍型亲缘关系较远.除保山外,其他烟田和工厂繁育种群单倍型差异较小.
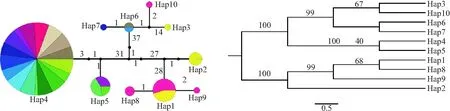
图1 云南省主要烟区烟蚜茧蜂种群单倍型网络图 图2 云南省主要烟区烟蚜茧蜂种群单倍型系统发育树
2.2 烟蚜茧蜂的种群遗传结构
云南多数种群之间成对的种群遗传分化系数没有显著性差异,且仅有同一单倍型,没有任何分化.除了保山、曲靖、红河仅有工厂繁育种群外,其他地区同州市的烟田种群与工厂繁育种群之间基本没有分化,具有相同的遗传背景;不同区域工厂种群间分化较低,差异不显著(p<0.05);野生种群(YNPEQ)与保山隆阳种群(BSLYG)基本无分化(Fst=-0.079),而与其他所有种群都有较大分化(Fst >0.5),其中与11个种群(只有Hap4)的遗传分化系数最大,为0.853 7;保山腾冲种群(BSTCG)与所有种群都有较大分化(Fst >0.5),其中与11个种群(只有Hap4)的遗传分化系数最大为0.789,与野生种群(YNPEQ)遗传分化系数最小为0.582 5.
对6个州市的烟田和工厂繁育种群(楚雄、丽江、昆明、大理、文山、昭通)的AMOVA分子方差分析结果显示,按照不同州市或烟田和工厂繁育种群不同类型划分的群组之间均没有显著分化,表明变异主要来自单个种群内部.由此可见,云南不同地区的烟田和工厂繁育种群具有较为一致的遗传背景.

图3 云南省主要烟区烟蚜茧蜂种群间分化系数(Fst)

表3 云南省主要烟区烟蚜茧蜂种群不同分组的遗传变异AMOVA分析
3 讨论
烟蚜茧蜂大规模扩繁技术成熟,防治烟蚜效果显著,但是种群衰退问题是限制烟蚜茧蜂应用的重要因素[26].随着烟蚜茧蜂防治烟蚜技术的全国推广,越来越多的繁蜂工厂被建立,明确人工释放对烟田种群遗传多样性的影响,对烟蚜茧蜂保护利用以及合理复壮有重要意义.云南省是我国烟草的主要种植区,也是最早开展烟蚜茧蜂工厂化繁育以及大规模释放和推广应用的地区[20].了解云南工厂繁育和烟田种群的遗传差异,可以为后续制订科学有效的种群扩繁、复壮和田间释放策略提供参考依据.
本研究中,半数烟田种群只有1种单倍型,对应地区的工厂繁育种群大部分也没有遗传多样性,全部19个烟田和工厂繁育种群中有17个种群都共享单倍型Hap4,该单倍型是分布最广泛的,在9个州市的工厂繁育种群均有出现,但是野生种群的所有单倍型均与Hap4相距较远,变异较大.同时,分子方差分析结果也显示烟田和工厂繁育种群遗传分化低,遗传背景相似,由此可见,烟田烟蚜茧蜂种群的遗传多样性与工厂繁育种群具有一致性,大规模释放的烟蚜茧蜂在烟田能够有效定殖,因此工厂繁育种群的遗传多样性反映了烟田种群的遗传多样性.不同州市间较低的遗传分化表明,这9州市的蜂种可能具有同样的来源,并且缺少野生亲本的补充.保山的2个工厂繁育种群遗传多样性较高,且与云南普洱的野生种群共享Hap1,与Hap8、Hap9的关系较近,遗传分化分析也显示保山工厂繁育种群与普洱野生种群基本无分化,而与其他种群分化较高,因此,保山的蜂种可能经历了来自野生种群的复壮,种群的遗传多样性得到提升.在以往的研究中,人工繁育种群的遗传多样性显著低于野生种群的现象在凡纳滨对虾、光裸方格星虫、兰州鲇等水产动物养殖以及大熊猫等珍稀动物保护的研究中都有类似的报道[33-36],但在天敌工厂化繁育及昆虫研究中尚未有相关报道.在工厂化繁育中,由于缺少野生亲本的补充,导致种群有效亲本数量降低,近交几率增加,遗传多样性丧失,种群衰退风险增大.而种群的衰退可能直接导致烟蚜茧蜂释放到烟田后控害能力下降,达不到预期的生物防治效果.
本研究选取了4个线粒体蛋白质编码基因作为分子标记,初步解析了云南省烟田烟蚜茧蜂种群遗传多样性的特点;总的来说,工厂繁育种群能够在烟田有效定殖,但是长期工厂化繁育可能对烟田和工厂种群的遗传多样性造成了一定影响,因此云南省主要烟区在繁育和释放烟蚜茧蜂进行生物防治时应注意及时补充野生亲本对蜂种进行合理复壮,同时应该开展长期的遗传多样性监测,警惕可能存在的种群衰退的风险.线粒体基因具有进化速率较快、母系遗传、同源性高且易于分析等特点,在解析物种的种群遗传结构方面具有一定的优势,但是线粒体基因由于其重要的功能往往受到较大的选择压力,在探究物种的适应性进化及其遗传机制等方面作用不足,后续可使用基因组层次SNPs等高分辨力的分子标记进一步研究,在基因组水平更好地理解烟蚜茧蜂的种群分化格局和种群衰退的遗传基础等问题.此外,本研究中种群的样本量较少,特别是缺少其他野生种群的样本,因此不能确定大规模繁育和释放对当地野生种群的影响,也不清楚云南省不同地区的野生种群分化程度如何,这需要后续进一步的取样分析.

猜你喜欢
清华金融评论(2022年4期)2022-04-13
云南画报(2021年5期)2021-07-22
国际放射医学核医学杂志(2021年10期)2021-02-28
云南画报(2020年9期)2020-10-27
房地产导刊(2020年7期)2020-08-24
中国外汇(2019年14期)2019-10-14
湖北农业科学(2017年6期)2017-04-26
现代农业科技(2016年22期)2017-03-24
现代农业科技(2017年1期)2017-03-06
现代农业科技(2016年21期)2017-03-06
