
中链脂肪酸在仔猪生产中的研究与应用
2021-01-25 06:45:50赵小刚郭吉余梁运祥
中国饲料 2021年1期
王 俊, 赵小刚, 郭吉余*, 梁运祥
(1.广东海大集团畜牧水产研究中心,广东广州 511400;2.华中农业大学,湖北武汉 430070)
抗生素作为抑菌剂可以抑制仔猪肠道内病原菌生长,为减少仔猪腹泻,提高仔猪生长性能和降低饲养成本曾做出巨大贡献。但是,随着抗生素的不规范使用,加速了抗生素耐药菌株的产生以及猪肉中抗生素残留,严重威胁动物和人类健康。我国于2020年7月全面禁止使用抗生素作为促生长剂。因此,寻找抗生素替代品用于维持仔猪的健康和维护公共安全迫在眉睫。近年来,中链脂肪酸(MCFAs)受到国内外研究者的广泛关注,其具有快速供能、抗菌、抗炎、抗氧化和提高机体肠道免疫的作用,本文就MCFAs在仔猪生产上的应用及发挥作用的可能机制进行综述,旨在为其作为饲料添加剂在生产养殖上的开发及合理利用提供理论依据。
1 中链脂肪酸的化学结构和特性
中链脂肪酸(MCFAs)是一类含有6~12个碳原子的饱和脂肪酸,分为偶数碳中链脂肪酸和奇数碳中链脂肪酸。偶数碳中链脂肪酸包括己酸(C6:0)、 辛 酸 (C8:0)、 癸 酸 (C10:0) 和 月 桂 酸(C12:0)。 偶数碳 MCFAs存在于天然食物中,如椰子油、棕榈仁油、牛奶和母乳中,目前所说的MCFAs一般指偶数碳MCFAs(Zentek等,2011;Jensen,2002;Sprong等,2001)。 奇数碳 MCFAs包括庚酸(C7:0)、壬酸(C9:0)和十一烷酸(C11:0)。奇数碳MCFAs在天然食物中含量极少,主要由化工合成。MCFAs与甘油发生酯化反应生成甘油三酯,被称为中链脂肪酸甘油三酯(MCTs)。与长链脂肪酸(LCFAs)相比,MCFAs熔点低,水溶性相对较高,在中性环境中基本解离。
2 中链脂肪酸的消化吸收和代谢特性
MCFAs/MCTs具有碳链短、分子量小和水溶性较高的特点。因此,MCFAs/MCTs的消化、吸收和代谢过程与LCFAs和长链脂肪酸酯 (LCTs)明显 不同 (Greenberger 和 Skillman,1971)。 因 为MCTs具有较高的水溶性,所以一般不需要胆盐乳化即可被胰脂酶快速水解为MCFAs和甘油单酯(Ferreira等,2014)。MCFAs在胃和十二指肠上段被快速吸收,少部分MCTs也可以不经过水解而直接被肠上皮细胞吸收 (Ferreira等,2014;Ramirez等,2001)。肠上皮细胞以被动扩散的方式吸收大部分未经降解的MCFAs,也有一部分被酯化后吸收(Carvajal等,2000)。在肠上皮细胞中,MCFAs与脂肪酸结合后蛋白亲和力低,因此不会被再次酯化而是扩散到门静脉中与白蛋白结合直接进入肝脏 (Guillot等,1994)。只有一小部分的MCFAs以乳糜微粒的形式被吸收 (Bach和Babayan,1982)。而 LCTs不溶于水,因此需要经过胆盐乳化才能被胰脂酶水解成LCFA和相应的单甘油酯。在肠上皮细胞中,LCFA和长链单甘油酯会被重新酯化成甘油三酯,然后与脂蛋白结合形成可溶性的乳糜微粒,通过淋巴系统进入血液循环被运输到肝脏 (Rego Costa等,2012)。因为MCFAs到达肝脏后不依赖肉毒碱的转运就可以通过线粒体膜进入线粒体,所以大部分MCFAs被线粒体以β-氧化的方式利用并快速供能(Rasmussen等,2002)。另外,MCFAs极少用于脂类合成,因此不会在肝脏和脂肪中蓄积。而LCFAs供能慢,进入线粒体依赖于肉毒碱转运。进入肝细胞内的LCFAs多用于脂类合成直接被转运到脂肪组织储存起来。
3 中链脂肪酸的抗菌作用
3.1 不同中链脂肪酸的抗菌特性 研究表明,MCFAs对细菌、病毒和寄生虫都具有抗菌作用(Zentek等,2011)。不同种类的MCFAs对革兰氏阳性菌和革兰氏阴性菌的抗菌效果并不一样(Skˇr ivanová等,2005;Skˇrivanová等,2004;Marounek等,2003)。总的来说,碳链长的MCFAs对革兰氏阳性菌的抗菌作用好,碳链短的MCFAs对革兰氏阴性菌的抗菌作用好。有学者系统比较了不同MCFAs对革兰氏阴性菌(E.coli和 Salmonellas)和革兰氏阳性菌(Clostridium perfringens)的抗菌效果。结果表明,C8对E.coli的抗菌效果最好,C10次之,C6和C12最差。C8对Salmonellas的抗菌效果最好,C6、C10和 C12次之。而 C12对Clostridium perfringens的抗菌作用最好,C10次之,C8和C6最差(Skˇrivanová等,2005;Skˇrivanová 等,2004;Marounek 等,2003)。 总的来说,C8对革兰氏阴性菌的抗菌作用最好,C12对革兰氏阳性菌的抗菌作用最好。
3.2 中链脂肪酸的抗菌机理 研究表明,MCFAs无论是对革兰氏阳性菌和革兰氏阴性菌都具有很好的抗菌效果,对细菌的直接损伤可能是其发挥抗菌作用的主要方式(图1)。一般认为,MCFAs属于非离子表面活性剂,可以进入细菌细胞膜并造成瞬间或永久的空隙,进而增加细菌细胞膜的通透性,引起细菌内容物释放,最终导致细菌死亡(Desbois等,2010;Altieri等,2009)。 而非解离形式的MCFAs进入细菌后会对其产生细胞毒性。由于细菌的生活和繁殖依赖于体内相关酶类的协调互作,在中性环境中,这些酶类能够发挥正常的生理功能。但是,由于MCFAs进入细菌细胞后导致细胞内pH降低,细菌细胞内酶类的活性随之迅速下降。进入细菌细胞质中的MCFAs会解离为H+和阴离子,降低胞质内的pH,进而灭活胞质中对pH敏感的酶类,最终导致细菌的死亡(Hsiao和Siebert,1999)。为了保持细胞质内pH的稳定,细菌被迫利用能量启动H+-ATPase泵出多余的H+,但是此过程会消耗大量能量,将造成细菌代谢障碍和衰竭,最终引起细菌的死亡(Suiryanrayna和Ramana,2015)。除了直接对细菌造成损伤外,MCFAs还可以诱导细菌产生一种自溶酶,引起细菌的死亡(Tsuchido等,1985)。
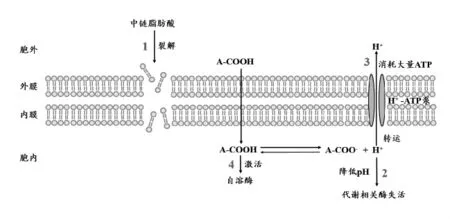
图1 中链脂肪酸的抗菌机制
4 中链脂肪酸的抗炎和抗氧化作用
胃肠道不仅是营养物质消化、吸收和代谢的重要场所,而且是重要的免疫器官,含有大量的免疫细胞。致病或非致病的刺激因素都可以激活胃肠道的免疫系统,产生多种特化的细胞和信号分子,如促炎因子TNF-α、IL-1β和IL-6(Wei等,2017;Xiong 等,2016)。 但是,过多的促炎因子会导致肠道黏膜损伤和功能紊乱。研究表明,中链脂肪酸具有抗炎作用。Nakatsuji等(2009)发现,痤疮丙酸杆菌可以诱导ICR小鼠产生炎症反应;而注射月桂酸后可以缓解小鼠的炎症反应。除了月桂酸外,癸酸也具有抗炎作用。Lee和Kang(2017)研究表明,环磷酰胺可以提高IPEC-J2细胞和小型猪肠道促炎因子 TNF-α、IFN-γ、IL-6和 IL-8的表达量,同时降低抑炎因子IL-4和IL-10的表达量。在添加癸酸处理后,显著改善了IPEC-J2细胞和小型猪肠道的炎症状态。除了改善炎症状态外,体外试验(小型猪)和体内试验(IPEC-J2细胞)均表明,癸酸还可以通过提高SOD和GPx的水平,降低MDA水平来缓解环磷酰胺诱导的氧化应激(Kang 和 Lee,2017)。
5 中链脂肪酸对仔猪生长性能的影响
MCFAs是一种容易得到的能源物质,在体内可以快速供能,还具有很强的抗菌活性。因此,MCFAs可以代替抗生素作为断奶仔猪饲料的良好补充剂,提高仔猪生长性能。但在断奶仔猪饲料中添加MCFAs对仔猪生长性能的影响并不一致,这可能与MCFAs添加类型、组合方式、添加剂量和试验周期有关(表1)。
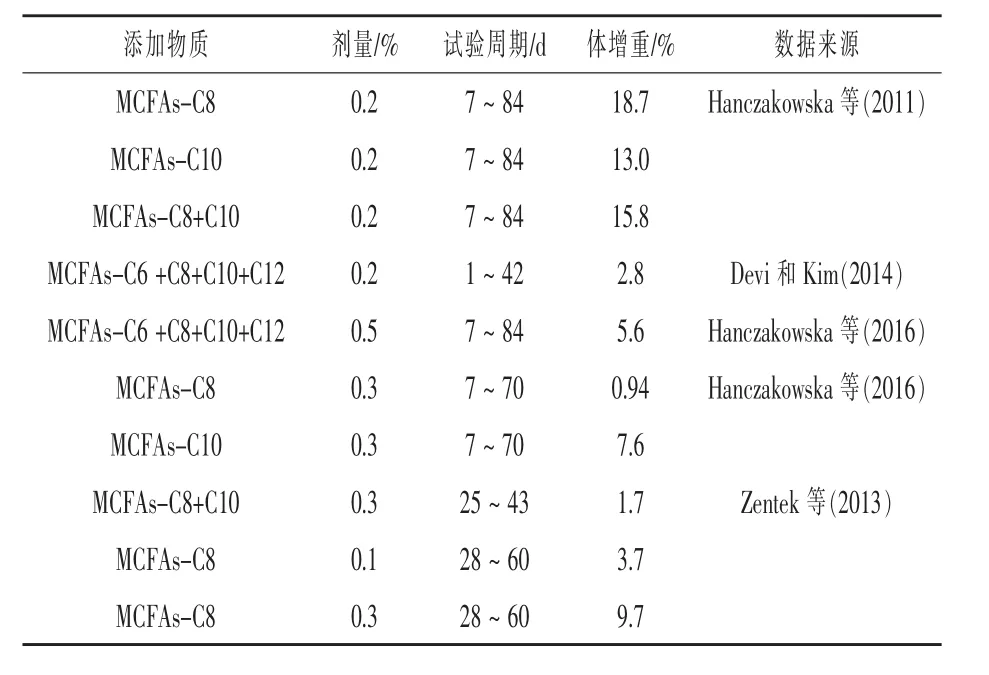
表1 MCFAs对仔猪生长性能的影响
MCFAs提高仔猪生长性能可能与其提高营养物质消化率,抑制胃肠道病原菌生长和保护肠道结构有关。Hanczakowska等(2010)研究表明,在仔猪饲料中添加MCFAs(C6+C8+C10+C12)降低了仔猪小肠中大肠杆菌和霉菌的数量,并且提高了仔猪小肠绒毛高度。Devi和Kim(2014)在仔猪饲料中添加MCFAs(C8或C10)提高了仔猪对饲料中干物质、氮和能量的利用率。Hanczakowska等(2011)在饲料中添加 MCFAs(C8,C10 和 C8+C10)提高了仔猪对干物质、粗蛋白质、粗脂肪和粗纤维的利用率;降低回肠中大肠杆菌、厌氧菌、梭状芽孢杆菌、白色念珠菌和念珠菌属的丰度,提高了绒毛高度。
6 结语
中链脂肪酸具有快速供能、抑制肠道中病原菌生长和抗炎抗氧化等作用,有利于预防仔猪腹泻、提高仔猪生长性能。中链脂肪酸来源于食物中,又具有光谱的抗菌作用,可以作为一种安全的抗生素替代品,具有广阔的应用前景。但是,中链脂肪酸对仔猪生长性能影响的结果并不一致,这可能与中链脂肪酸添加类型、组合方式、添加剂量和试验周期有关。因此,深入研究中链脂肪酸的效果以及其作用机理,不仅有利于稳定中链脂肪酸的作用效果,而且能为畜禽新型饲料添加剂的开发和合理利用提供理论依据。
猜你喜欢
中国典型病例大全(2022年12期)2022-05-13 18:24:49
中国动物保健(2021年8期)2021-11-24 01:40:22
祝您健康·文摘版(2021年8期)2021-08-10 21:24:04
猪业科学(2021年1期)2021-02-24 03:28:50
中国生殖健康(2020年5期)2021-01-18 03:00:06
中国饲料(2020年16期)2020-11-06 06:57:56
中国生殖健康(2018年5期)2018-11-06 07:15:56
今日畜牧兽医(2018年12期)2018-02-14 00:24:58
浙江农业学报(2017年1期)2017-05-17 06:13:47
中国洗涤用品工业(2015年8期)2015-02-28 19:02:49
