
小鼠神经系统中蛋白质降解通路调控分子的表达和分布
2021-01-21 01:25宋彬彬董文洲贾炳泉
实验动物与比较医学 2020年6期
宋彬彬, 董文洲, 贾炳泉, 甄 然, 彭 宇, 杨 璇, 于 佳
(北京中医药大学附属北京老年医院,北京 100095)
蛋白质降解是细胞重要的生命活动,具有蛋白质量控制、清除细胞内错误折叠蛋白质、促进蛋白激活等作用。真核细胞内蛋白质降解通路主要有3种:泛素-蛋白酶体系统(ubiquitin-proteasome system,UPS)、自噬(autophagy)和钙蛋白酶(Calpain)系统,三者降解底物或作用部位各有不同[1-2],且受到多种分子的精细调控,包括UPS中的泛素(Ubiquitin),自噬通路中的组织蛋白酶D(Cathepsin D)、溶酶体关联膜蛋白2(lysosomalassociated membrane protein 2,LAMP2)、自噬微管相关蛋白轻链3A/B(microtubule-associated protein 1A/1B-light chain 3,LC3A/B)、自噬相关蛋白3(autophagy-related protein 3,Atg3)和Ras相关GTP结合蛋白7(Ras-related GTP-binding protein 7,Rab7),以及Calpain途径中的Calpain和钙蛋白酶抑制蛋白(Calpastatin)等。本实验通过生物化学方法,以小鼠眼、脑、脊髓、坐骨神经和肌肉组织为材料,检测C57BL/6J小鼠完整的神经信号转导通路中蛋白质降解通路相关蛋白在不同组织、解剖部位和亚细胞结构(胞体和轴突)的表达和分布差异。
1 材料与方法
1.1 实验动物
6周龄野生型C57BL/6J小鼠,雄性4只,雌性1只,体质量16~20 g,购自北京维通利华实验动物技术有限公司[SCXK(京)2016-0011],饲养于北京老年医院屏障设施[SYXK(京)2019-0012],根据实验动物相关管理规定进行操作和处理。小鼠处死后迅速取眼(eye)、嗅球(olfactory bulb)、皮层(cortex)、海马体(hippocampus)、纹状体(striatum)、中脑(midbrain)、小脑(cerebellum)、脑干(brain stem)、脊髓(spinal cord,SC)、坐骨神经(sciatic nerve,SN)和肌肉(muscle),冻存于-80℃冰箱。动物实验经本单位动物福利伦理委员会批准(2018BJLNYY-伦理-意见第2018-013号)。
1.2 主要试剂和仪器
10%十二烷基硫酸钠(SDS)溶液、蛋白酶抑制剂、PierceTMBCA 蛋白定量试剂盒、蛋白质电泳预制胶和超声波细胞破碎仪均购自美国Thermo Fisher Scientific公司;10% TGX Acrylamide Kit、Turbo小型预制转印包和Trans-Blot Turbo转印系统均购自美国Bio-Rad公司;兔抗Calpain1大亚基(large)多克隆抗体、兔抗Calpastatin多克隆抗体、兔抗LC3A/B多克隆抗体和兔抗Atg3多克隆抗体均购自美国Cell Signaling Technology公司,大鼠抗LAMP2单克隆抗体、兔抗Ubiquitin单克隆抗体、鼠抗Rab7单克隆抗体和鸡抗神经丝重链(neurofilamen heavy polypeptide,NFH)多克隆抗体均购自英国Abcam公司,鼠抗β-actin和Calpain小亚基(small)单克隆抗体均购自美国Sigma-Aldrich公司,山羊抗Cathepsin D多克隆抗体购自美国R&D Systems公司;荧光标记二抗和Odyssey CLX双色红外激光成像系统均购自美国LI-COR Bioscience公司。
1.3 蛋白质印迹法检测蛋白表达水平
速冻的小鼠组织称质量后,按30 µL/mg加入 1% SDS裂解液,迅速剪碎,超声破碎,室温16 800 ×g离心10 min,收集上清液为总蛋白提取液。用BCA试剂盒测定蛋白含量,取各组等量蛋白提取液,经10% SDS-PAGE(Bio-Rad TGX acrylamide kit)或4%~12% NuPAGE Bis-Tris预制凝胶电泳分离,再将凝胶中的蛋白质电转移至硝酸纤维素膜上。将硝酸纤维素膜放入5%脱脂牛乳中室温封闭1 h或Odyssey封闭液中室温封闭30min后,加入山羊抗Cathepsin D(1∶1 000)、大鼠抗LAMP2(1∶1 000)、兔抗LC3A/B(1∶1000)、兔抗Atg3(1∶1000)、鼠抗Rab7(1∶1 000)、兔抗Ubiquitin(1∶500)、兔抗Calpain1 large(1∶1 000)、鼠抗Calpain small(1∶500)、兔抗 Calpastatin(1∶1 000)、鸡抗NFH(1∶5 000)、鼠抗β-actin(1∶10 000)一抗,4 ℃孵育过夜。PBST(含0.1% Tween-20的磷酸盐缓冲液)漂洗3次(每次5 min)后,将膜放入荧光素IRDye标记的山羊抗兔或山羊抗鼠IgG(1∶5 000)二抗中,室温避光孵育1 h,PBST充分漂洗后,采用Odyssey凝胶成像系统检测蛋白条带的显色情况,使用Image J软件分析蛋白条带的相对灰度。
1.4 统计学分析
使用统计软件GraphPad Prism 7(美国GraphPad Software公司)进行分析。数据以表示,空间分布用柱形图表示,两组均数间差异比较采用t检验,P<0.05为差异有统计学意义。
2 结果
2.1 自噬和钙蛋白酶途径调控分子在小鼠神经系统中的分布
蛋白质印迹法检测结果显示,Cathepsin D、LAMP2、LC3A/B、Atg3、Rab7、Calpain大小亚基和Calpastatin在小鼠眼、嗅球、皮层、海马、纹状体、中脑、小脑、脑干、SC、SN和肌肉中都有一定表达(图1A),但含量具有组织和解剖部位差异性(图1B)。肌动蛋白β-actin为非肌肉型肌动蛋白,在神经系统中广泛表达,在肌肉中不表达。
2.2 蛋白质降解调控分子在小鼠脊髓和坐骨神经中表达和分布
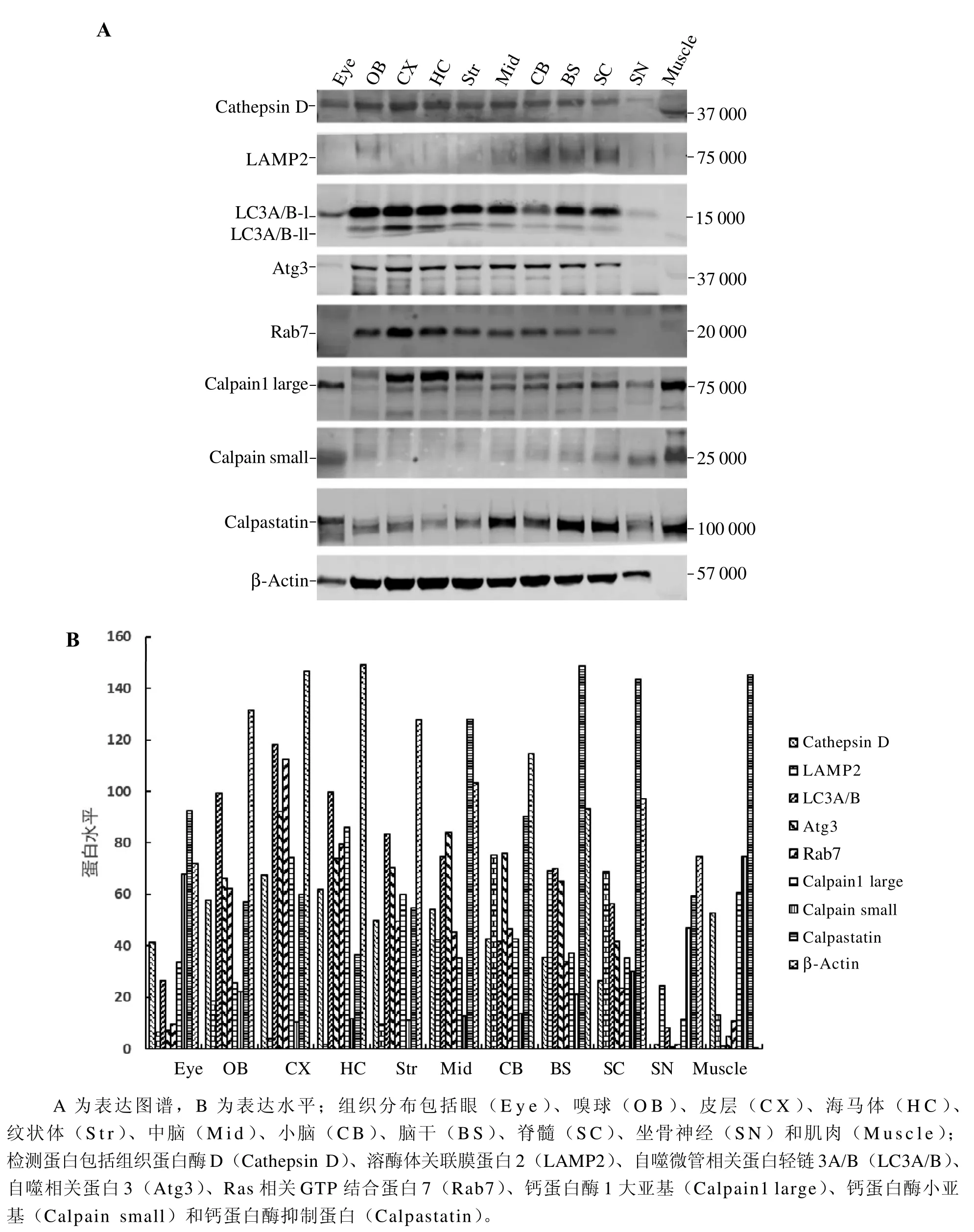
图 1 蛋白质印迹法检测自噬和钙蛋白酶途径调控分子在小鼠组织中表达Figure 1 Expression of autophagy and Calpain pathway molecules in the tissues of mice detected by Western blotting
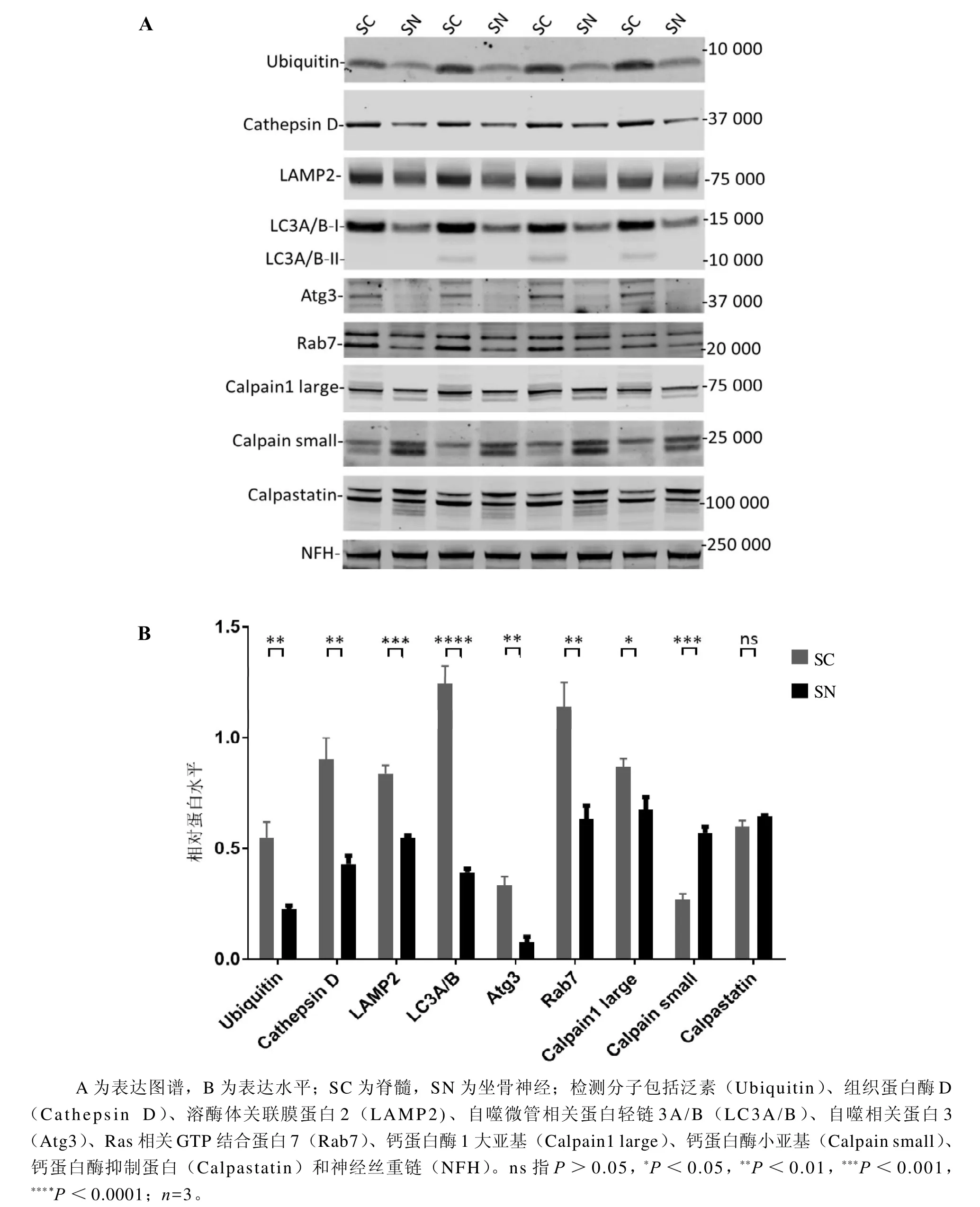
图 2 蛋白质印迹法检测蛋白质降解通路调控分子在小鼠脊髓和坐骨神经中表达和分布Figure 2 Expression of protein degradation pathway molecules in the spinal cord and sciatic nerve of mice detected by Western blotting
蛋白质印迹法检测结果显示,蛋白质降解通路调控分子在小鼠SC和SN中有差异性表达(图2A)。与SC相比,小鼠SN中UPS关键蛋白Ubiquitin和自噬通路中的Cathepsin D、LAMP2、总LC3A/B、Atg3、Rab7含量均明显下降(P<0.01),且未检测到LC3A/B-Ⅱ蛋白;钙蛋白酶途径中的Calpain1 large总含量(包含全长相对分子量80 000和剪切型相对分子量75 000)明显下降(P<0.05),但剪切型Calpain1 large含量相对增加;而Calpain small总含量(包含全长相对分子量26 000和剪切型相对分子量18 000)明显增加(P<0.001),且剪切型Calpain small含量增加相对明显;Calpastatin含量无明显差异(P>0.05)(图2B)。
3 讨论
UPS是高效、高度选择的蛋白降解途径,其广泛参与机体多种代谢活动,主要降解短寿命蛋白质和应激条件下细胞内异常蛋白。UPS由Ubiquitin、泛素激活酶(E1)、泛素结合酶(E2)、泛素连接酶(E3)、蛋白酶体和去泛素化酶组成。在ATP作用下,Ubiquitin被活化后与E1结合,转移到E2,再通过E3与靶蛋白结合,泛素化的靶蛋白进入26S蛋白酶体(ATP依赖性蛋白水解复合体)被降解为多肽或氨基酸。去泛素化酶可将Ubiquitin从底物上解离下来,继续识别其他待降解蛋白质[3-4]。Pasquini等[5]早期研究提出,SN中不易形成泛素-蛋白质偶联物。本研究发现,与SC相比,SN中Ubiquitin含量明显减少,这可能使蛋白质泛素化过程受阻,从而引起轴突束中UPS降解蛋白质功能减弱。
自噬是真核细胞中广泛存在的一种溶酶体依赖的降解途径,较UPS具有更强的降解能力,可降解长寿命蛋白质(错误折叠蛋白、蛋白质复合物、积聚物)以及受损细胞器和细胞内病原体[6]。自噬过程分为3个阶段:启始、成熟和降解[7]。自噬起始复合物(ULK1、ULK2、ATG13、FIP200和ATG101)磷酸化后激活,活化的ULK1可以招募自噬相关蛋白(Atg)到自噬体形成位点,触发自噬体形成[8]。前自噬体结构(pre-autophagosomal structure,PAS)初步延伸可包裹待降解物,随后形成Atg5-Atg12-Atg16复合物,与3-磷酸磷脂酰肌醇结合且定位到自噬体膜上使膜延伸;或者Atg8经泛素样酶Atg7和Atg3加工后,与磷脂酰乙醇胺(phosphatidyl ethanolamine,PE)结合形成Atg8-PE,Atg8-PE位于自噬体膜,促进自噬体膜延伸和招募待降解物[9]。细胞质内Atg4家族蛋白还可以剪切LC3前体生成LC3Ⅰ,LC3Ⅰ可通过Atg7和Atg3与PE偶联后形成LC3Ⅱ,故LC3Ⅱ特异性与自噬体结合,LC3Ⅱ水平与自噬体数量正相关,可作为自噬体形成的标志物[10]。包裹待降解蛋白质或细胞器的自噬体(autophagosome)被运输到溶酶体中被降解,且降解产物可供细胞再循环利用,以维护细胞内稳态。Cathepsin D是溶酶体中主要水解酶[11]。LAMPs是溶酶体膜上含量丰富的跨膜蛋白,LAMP1和LAMP2共占溶酶体膜蛋白的约80%,可维持溶酶体膜完整性[12]。LAMPs在溶酶体腔面的氨基末端高度糖基化,以防止溶酶体腔内酸性水解酶对其自身结构造成破坏[13]。细胞内吞形成内体,晚期内体也可携带特定待降解蛋白质,与溶酶体融合后进行降解。Rab7既位于晚期内体,也定位于自噬体,参与调控自噬体与溶酶体的结合,影响自噬体成熟和运输[14]。本研究发现,与SC相比,SN中Cathepsin D、LAMP2、总LC3A/B、Atg3和Rab7含量均明显减少,且未检测到LC3A/B-Ⅱ。这说明自噬体和溶酶体主要分布在神经元胞体,轴突束中自噬体形成和溶酶体分布均相对减少。有研究发现,当SN受到损伤的早期,LC3-Ⅱ蛋白含量显著增加,自噬水平提高可促进轴突、髓鞘再生和运动功能恢复[15-16]。
当细胞受损、刺激等引起细胞质中钙离子水平升高时,Calpain被活化,发挥蛋白降解功能。Calpain可水解细胞骨架蛋白、膜相关蛋白、激酶和磷酸酶、转录因子和凋亡相关蛋白等百余种蛋白质,参与维护细胞结构、信号转导、死亡等过程[17-18]。根据激活所需钙离子浓度差异可将Calpain分为µ-Calpain(Calpain1)和m-Calpain(Calpain2),二者是哺乳动物普遍表达类型,均由具有催化活性的大亚基和具有调节活性的小亚基组成[19]。Calpain大亚基具有4个主要结构域:当Calpain大亚基被钙离子激活后,结构域Ⅰ发生自溶,可暴露水解活性域;结构域Ⅱ是蛋白质水解的主要功能域,可分为Ⅱa和Ⅱb,钙离子水平较低时,两亚区域分离,抑制蛋白质水解;结构域Ⅲ为调节中心,参与Calpain膜转移过程;结构域Ⅳ含有5个EF手性结构域,直接与钙离子结合后,酶构象改变,大小亚基分离,逐步暴露水解活性结构域Ⅱa和Ⅱb,参与底物水解[20]。本研究中Calpain1 large抗体可检测全长(相对分子量80 000)和28位亮氨酸自剪切后(相对分子量75 000)的Calpain1large蛋白。与SC相比,SN中总Calpain1large含量明显减少,但剪切型Calpain1large蛋白(相对分子量75 000)含量相对增加。Calpain1和Calpain2具有相同的Calpain small,Calpain small为调节亚基,具有Ⅰ、Ⅱ两个结构域,对于维持Calpain活性具有重要作用。本研究中Calpian small抗体可检测全长(相对分子量26 000)和自溶剪切型(相对分子量18 000)Calpain small。研究发现,与SC相比,SN中总Calpain small含量明显增加,且剪切型Calpain small含量增加相对明显,提示Calpain small较活化。Calpain活性随钙离子浓度提高而增加,当Calpain过度活化时,Calpastatin可特异性抑制Calpain活性[21]。Baudry[22]研究提出,Calpain广泛分布于机体各组织。本研究进一步发现,在神经系统中,SC与SN相比,Calpastatin含量无明显差异;而SC和SN中Calpain大小亚基含量和活性具有明显差异,提示Calpain调节的蛋白质降解主要发生在神经元轴突,较少发生于胞体。当SN受损时,轴突内钙离子水平迅速升高可激活Calpain[23],降解细胞骨架蛋白使轴突变性[24],而Calpastatin对轴突和突触结构及功能具有保护作用[25]。
蛋白质合成与降解平衡是维持细胞内稳态的重要条件,蛋白质降解通路受损引起蛋白质异常积聚可导致多种神经退行性疾病。本研究发现,与SC相比,小鼠SN中UPS和自噬通路相关蛋白含量明显降低,而依赖于钙离子的Calpain系统中Calpain small含量明显增加,这提示UPS和自噬主要降解神经元胞体中蛋白质,而轴突束中主要通过Calpain途径降解底物。在正常生理条件下,Calpain参与调节细胞迁移和轴突发育等;但在氧化应激、创伤、兴奋毒性等病理条件下,钙离子大量内流,Calpain高度活化,可能导致轴突变性、断裂,甚至神经元死亡。蛋白质降解通路异常与疾病发生发展紧密相关,深入研究蛋白质降解通路及其关键分子可为神经系统疾病的治疗提供新的思路。
猜你喜欢
浙江临床医学(2021年12期)2021-11-29
生物化工(2021年2期)2021-01-19
世界最新医学信息文摘(2020年68期)2020-12-25
生物化工(2020年1期)2020-02-17
烟台大学学报(自然科学与工程版)(2020年1期)2020-02-08
读与写(2019年35期)2019-11-05
现代职业教育·高职高专(2018年7期)2018-05-14
中华神经创伤外科电子杂志(2015年1期)2015-01-21
中南民族大学学报(自然科学版)(2014年2期)2014-01-22
中国医学科学院学报(2013年6期)2013-03-11
