
甘薯近缘野生种Ipomoea trifida(4 x)的染色体核型分析
2021-01-16 13:07李丽,李云,张璠,潘牧,3,杨航
种子 2020年12期
李 丽, 李 云, 张 璠, 潘 牧,3, 杨 航
(1.贵州省农业科学院生物技术研究所, 贵阳 550006; 2.贵州大学生命科学学院, 贵阳 550006;3.贵州农科特创农业科技开发有限公司, 贵阳 550006)
甘薯[Ipomoeabatatas(L.)Lam.]是重要的粮食、饲料、工业原料及新型能源用块根作物,广泛种植于100多个国家,在世界粮食生产中总产列第7位。我国甘薯年种植面积约660万hm2,占世界甘薯种植总面积的65.4%,年产量约1.5亿t,占世界甘薯总产的85.9%[1]。其原生种主要涉及野生近缘种Ipomoeatrifida的2 x、4 x和6 x多倍体复合体[2]。甘薯近缘野生种资源丰富,携带优良的抗性基因,在甘薯育种上发挥了极其重要的作用。由于甘薯染色体较小,数目较多,细胞质浓厚和染色能力弱,因而对其细胞学研究相对薄弱,滞后于水稻、小麦等其他作物[3]。国外对甘薯的染色体研究起步较早,薛启汗等[4]报道,甘薯近缘种I.tiliacea为二倍体 (2 n=2 x=30),I.trifida为六倍体(2 n=6 x=90),I.littoralis为四倍体(2 n=4 x=60)。与国际先进水平相比,我国甘薯的染色体核型研究工作起步较晚,现主要由中国农业科学院、徐州甘薯研究中心等单位开展工作,目前已取得较大进展。曹清河等[5]利用染色体压片法和花粉粒扫描电镜法对甘薯近缘野生种IpomoeahederaceaJacq.的体细胞染色体数目、核型及其分类地位进行分析和界定,明确了该甘薯近缘种的细胞遗传学信息及其在甘薯属中的进化程度。汤佳立等[3]利用荧光原位杂交技术分析栽培种甘薯(Ipomoeabatatascv.Xushu 18)染色体,发现徐薯18染色体上有8对或9对强弱不一的45 S rDNA信号,所有染色体的全长分布强烈而密集的杂交信号,着丝粒区、近着丝粒区和端粒区有增强的信号带。梁国鲁等[6]对甘薯近缘野生种Ipomoeatrifida的二倍体(2 n=2 x=30)和六倍体(2 n=2 x=90)的核型进行了观察分析,探讨了该多倍体复合体的染色体结构变异与系统演化。而关于甘薯近缘野生种Ipomoeatrifida(4 x)的染色体特征研究甚少,其核型信息也缺乏。因此,本研究采用常规染色体压片法对甘薯近缘野生种Ipomoeatrifida(4 x) 的染色体数目进行计数和核型分析,为探讨甘薯多倍体的遗传学特征、系统演化及杂交亲本选配提供细胞学依据。
1 材料与方法
1.1 材 料
供试材料甘薯近缘野生种Ipomoeatrifida(4 x)由国家甘薯产业体系贵阳综合试验站提供。
1.2 方 法
1.2.1染色体制片
试验材料均用甘薯近缘野生种Ipomoeatrifida(4 x)组培苗萌发生根,用改良的去壁低渗法[7]制片。
预处理:待根长1~1.5 cm时,洗净置于吸水纸上,吸去水分后并将根尖放在浓度为0.002 mol·L-1羟基喹啉中,室温处理40~60 min,取出用蒸馏水漂洗3~5次。
前固定:用卡诺氏固定液(甲醇∶冰醋酸=3∶1)对前低渗后的甘薯根尖进行固定2 h。
前低渗:将漂洗好的根尖材料放置蒸馏水中用蒸馏水作为前低渗液浸泡30 min,使染色体易于分散。
解离:将前低渗好的根尖材料放置于0.2 mol·L-1的盐酸溶液中在50~60 ℃温度下解离8~10 min。
后低渗:将解离好的根尖材料用蒸馏水漂洗3~5次用最后1次蒸馏水作为后底渗,低渗30 min。
染色:将低渗后的根尖置于干净载玻片上,加卡宝品红染液,染色15~30 min。
漂洗:将染色好根尖的材料用45%醋酸溶液漂洗8~10 min。
压片:将漂洗好的根尖(约0.2 cm),置于盖玻片上,在显微镜下观察、拍照。
1.2.2染色体镜检方法
选取染色体完整、分散良好、着丝点清晰的中期分裂相进行染色体计数,在放大倍数为10×100倍镜下显微照相,并结合Adobe Photoshop CS软件进行调整,对图片进行排序、剪贴、制成核型图。
1.2.3核型分析
采用李懋学等[8]的方法进行核型分析,Image-pro plus软件对染色体的数目、臂长(包括长臂和短臂)进行测量,然后按Levan等[9]的方法计算相应的数据(包括相对长度、臂比值)。

表1 着丝点位置划分
2 结果与分析
2.1 甘薯近缘野生种Ipomoea trifida(4 x)的染色体数目
对100个近缘野生种Ipomoeatrifida细胞的染色体数目及形态进行分析,发现近缘野生种染色体为四倍体型。根尖组织细胞中存在染色体数有2 n=50、52、54、56、58,其中含有56条染色体的细胞数所占比例达到58%,根据生物体染色体数目统计原则,甘薯近缘野生种Ipomoeatrifida的染色体数目为2 n=4 x=56。

表2 Ipomoea trifida的核型参数
2.2 甘薯近缘野生种Ipomoea trifida(4 x)的染色体特征
甘薯近缘野生种Ipomoeatrifida的染色体数目为56条,可分为14组(A-N),每组含有4条染色体(图1)。甘薯近缘野生种Ipomoeatrifida的臂比范围为(1.20±0.36)~(3.64±0.36)之间,相对长度介于(23±6.93)~(26±5.96)之间。根据 Levan等[9]对着丝点的位划分标准,臂比大于2 的染色体所占比例为0.64,在0.51~0.99 之间,且最长臂/最短臂比值约为1.43,核型不对称系数为58.22%,其核型分类属3 A型。其核型公式为:2 n=4 x=56=6 m+4 sm+1 st+3 t。
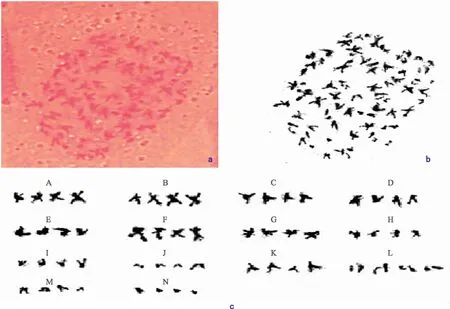
注:a,b为甘薯近缘野生种Ipomoea trifida染色体原始图; c为甘薯近缘野生种Ipomoea trifida染色体配对图
3 讨 论
Ipomoeatrifida是一个复合种,包括二倍体、 四倍体、六倍体及其杂种,广泛分布于墨西哥-厄瓜多尔-巴西的热带美洲[6]。将甘薯品种之间进行任意杂交,能够分离出茎细、不结块根的野生类型, 从表型上很难将其分开, 因此可以通过现代生物技术、基因工程等方法将其区别。染色体核型分析是开展杂交育种、多倍体诱导和雌核发育等遗传育种工作的基础,对研究植物系统演化、物种之间的亲缘关系、起源、进化与分类,远缘杂交及遗传工程中的染色体鉴别具有重要意义[10]。甘薯近缘野生种被认为是开拓甘薯育种新的抗病、优质、抗逆等基因的重要来源之一,以近缘野生种作母本,综合性状较好的地方品种、新育成品系和外引品种资源作父本,让其杂交,已成为甘薯常规育种的手段之一。Ipomoeatrifida(4 x)种包含多个系统,推测有的Ipomoeatrifida(4 x)株系是Ipomoeatrifida(2 x)直接加倍产生的,有的是基因组重组与部分变异产生的,但表型均为野生型[11]。由于甘薯染色体的总体特征表现为小且多,本试验中,甘薯材料黔薯7号和黔薯6号因淀粉粒较多,染色难,不能确定其染色体数目和核型特征。本研究仅对甘薯近缘野生种Ipomoeatrifida(4 x)作了初步分析,要深入探讨甘薯的起源问题,还需结合分子生物学手段对不同类型、品种的甘薯进一步开展核型分析。
猜你喜欢
医疗卫生装备(2022年2期)2022-03-16
云南医药(2021年6期)2022-01-08
小猕猴智力画刊(2021年6期)2021-08-05
川北医学院学报(2021年6期)2021-07-13
科技创新与应用(2017年19期)2017-07-08
中国实用医药(2016年36期)2017-06-20
作文大王·低年级(2016年3期)2016-03-11
湖北农业科学(2015年17期)2015-10-09
少年科学(2013年5期)2013-06-08
中学理科·综合版(2008年3期)2008-03-07
