
温热玉米T 32/齐319 F2∶3家系光周期敏感性分析
2021-01-16 09:58郭向阳王安贵刘鹏飞祝云芳陈泽辉
种子 2020年12期
刘 颖, 吴 迅, 郭向阳, 何 兵, 王安贵, 刘鹏飞,高 媛, 涂 亮, 祝云芳, 陈泽辉
(1.贵州大学农学院, 贵阳 550025; 2.贵州省农业科学院旱粮研究所, 贵阳 550006)
20世纪20年代,Garner 和 Allard 将Maryland Mammoth(一个烟草变种)南种北移时发现,短日照是这种烟草开花的关键条件,发现植物开花与日照长度有关,将植物通过感受日照长度长短变化而抑制开花的现象称为光周期现象[1]。植物对光周期现象反应的敏感程度称为光周期敏感性。热带或亚热带种质具有适应性广、抗性好、品质优良等优点。在现代育种中,引进热带、亚热带种质并进行驯化和改良,已经成为解决当前种质基础狭窄问题的有效途径之一。热带、亚热带玉米种质资源的引进和利用,有助于进一步发掘新的杂种优势类群和杂种优势模式,提高杂种优势利用水平[2]。玉米是一种短日照作物,温带玉米在很大程度上对日长反应不敏感,而热带玉米种质通常对日长的变化反应较为敏感[3]。陈彦惠等对热带玉米抽雄期、散粉期、吐丝期、株高、穗位高等性状的遗传分析表明,热带种质在温带条件下表现出明显的光周期敏感性[4]。此外,王铁固等利用玉米自交系黄早四和CML 288进行长短日照相互挪移处理,观察玉米茎尖的形态变化,通过对玉米的全株叶片数、生育期等性状进行调查发现,长日照能抑制玉米茎端分生组织向生殖生长转化,抑制开花,增加玉米的主茎叶片数,延长玉米的生育期[5]。王翠玲等利用永久F2群体在不同光周期环境下对玉米株高进行QTL定位,结果表明,随着纬度、光照长度的增加,群体间的株高表现出显著差异[6]。库丽霞等在长日照环境条件下对以温带自交系黄早四和热带自交系 CML 288 为亲本配置的组合衍生的一套 207个重组自交系不同发育时期的主茎叶片数、株高变化进行QTL分析,结果表明,黄早四和CML 288间的主茎叶片数和株高差异很大[7]。因此,热带或亚热带玉米种质引入温带地区时,随着纬度和光照长度的增加,往往表现出光周期敏感性强,即营养生长旺盛,茎节数和叶片数增多,生殖生长受到抑制,抽雄期、散粉期、吐丝期延迟,晚熟,雌雄不协调,甚至不能正常结实等明显的不适应现象,严重影响其在育种实践中的应用。由此可见,玉米光周期敏感性已经严重限制了热带种质在温带地区的引用,研究玉米光周期敏感性及钝化玉米光周期敏感性对现代玉米育种的改良具有重要意义。
本研究以对光周期敏感的热带自交系T 32(母本)和钝感的温带自交系齐319(父本)为亲本,通过杂交、自交获得150份F2∶3家系为材料,在不同光照环境(贵州贵阳和甘肃张掖)条件下对抽雄期、散粉期、吐丝期、株高、穗位高、叶片数等6个光敏相关性状进行田间表型评价,研究不同性状间变异方向以及相互关系,为今后培育高光效玉米种质提供依据。
1 材料与方法
1.1 试验材料
本研究以热带来源的玉米骨干自交系T 32(母本)和温带核心玉米自交系齐319(父本)为亲本(表1),构建的包含150份“T 32×齐319”的家系为材料。其中,自交系T 32是贵州省农业科学院旱粮研究所通过从泰国引进的自交系中选择变异株选育而成的[8],该自交系具有抗病性好、籽粒品质优、产量高等优势,但也存在长光照条件下表现出生育期长、茎节数和叶片数增多、雌雄不协调、不能正常结实的缺点,属于典型的光周期敏感的玉米自交系。齐319是山东省农业科学院玉米研究所利用美国先锋公司选育的玉米杂交种PN 78599,经多代自交选育而成[9],该自交系具有持绿性好,高抗病性,但在南方生态区的抗病性差,光周期钝感的特点。
2017年春,在贵州贵阳(26.33°N,106.64°E)以T 32作母本,齐319作父本,通过杂交获得F1种子。2017年冬,在海南九所(18.26°N,108.54°E)播种F1,套袋自交获得F2单穗种子,2018年春采用单籽粒法构建 150个F2∶3家系,即为本研究的供试材料。
1.2 试验方法
2019年春,将150份F2∶3家系分别种于贵州省贵阳市(纬度为26.57°N、海拔为1 200 m、平均日长为13.3 h)与甘肃省张掖市(纬度为38.93 °N、海拔2 400 m、平均日长为14.2 h)。试验采用完全随机区组设计,2次重复,单行区,行长3.5 m,行距70 cm,过道50 cm,每行14株。田间调查以小区为单位,共调查抽雄期(d)、散粉期(d)、吐丝期(d)、株高(cm)、穗位高(cm)、叶片数(片)等6个农艺性状,参考石云素等[10]的方法进行,即在灌浆期,每行从第2株开始连续选取5株。
1.3 统计分析方法
数据的初步整理利用Excel 2016软件进行,统计F2∶3家系各性状平均值和变异范围,对2点6个性状进行方差分析。用SPSS 26.0软件对2个环境F2∶3家系各性状作相关性分析、正态分布检测(包括峰度、偏度等参数)。
1.4 光周期敏感性评价
评价玉米光周期敏感性的指标不一,有研究认为,抽雄期、散粉期、吐丝期、株高、穗位高、全株叶片数等相关性状均能较好的反映出玉米光周期敏感性,并以这些相关性状作为光周期敏感性的评价指标[11-12]。为了使指标量化,张凤路等[13]利用敏感性指数=(某种质在长光下的观测值/ck 种质在长光下的观测值)×(某种质的敏感性系数/ck 种质的敏感性系数) ,即敏感指数=某性状在自然光照条件下的观测值/该性状在长光照条件下的观测值。或者用某性状在长、短日照条件下的相对差值(Relative differences,RD)来估算某一材料某一性状的光周期敏感指数[14-16]。计算公式如下:
RD(%)=[(Li-Si)/Si]×100%
其中:Li表示某一材料某一性状在长日照条件下表现的平均值,Si表示某一材料某一性状在短日照条件下表现的平均值。为了使家系间的光周期敏感指数能够进一步量化,同一个材料的每个光敏相关性状的RD值相加求和[17],即:RDs=∑ |RD| 。
2 结果与分析
2.1 两亲本间的差异分析及各个性状RD值
通过对2个亲本的差异分析可知,齐319在短光(贵阳)和长光(张掖)条件下株高、穗位高、叶片数均值无太大变化,T 32的株高、穗位高、叶片数最大值、最小值、均值在短光(贵阳)条件下均远远小于长光(张掖)条件下(图1)。
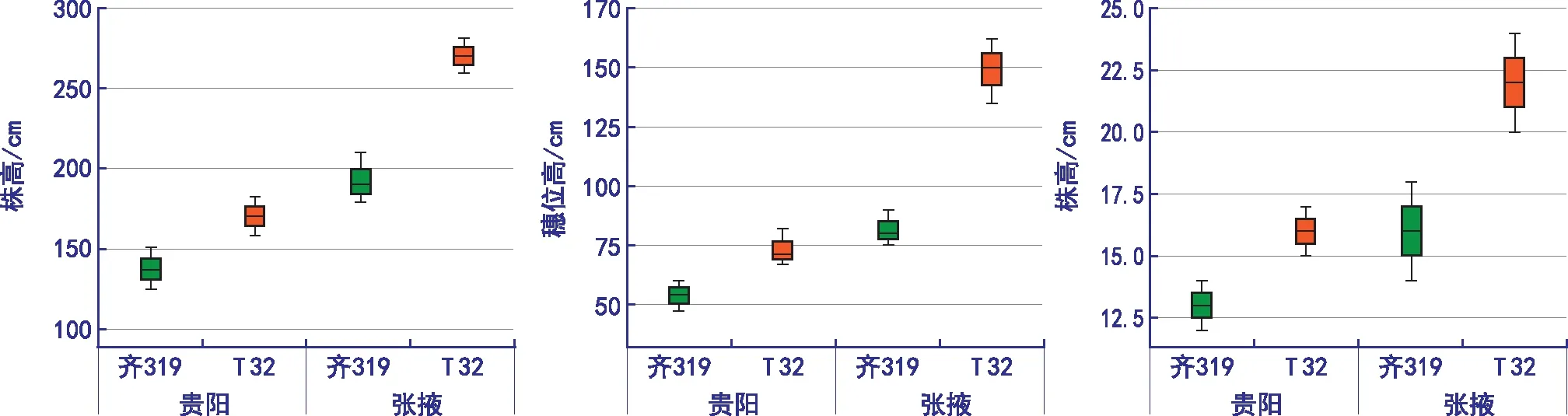
图1 不同环境条件下光敏系数比较
从表2可知,6个光敏相关性状的敏感程度为:穗位高>吐丝期>抽雄期>散粉期>株高>叶片数。
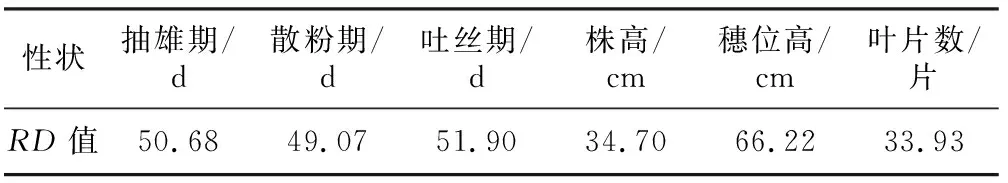
表2 F2∶3家系中6个光敏性状的RD值
2.2 F2∶3家系各性状方差分析和群体内家系的正态性检测
F2∶3家系各性状方差分析和群体内家系的正态性检测结果列于表3,结果显示,贵阳、张掖2个环境的抽雄期、散粉期、吐丝期、株高、穗位高、叶片数在家系之间的F值均达到极显著水平。表明150个家系间存在真实的遗传差异,可做进一步遗传分析。

表3 光敏相关性状分析
2个环境下F2∶3家系各性状偏度、峰度绝对值均小于2,且变异系数有较大幅度的变化,表现出数量性状遗传正态分布的典型特征。
2.3 光敏相关性状的频率分布
从频率分布图可以看出,总体变数比较集中,短光照(贵阳)的株高和穗位高的峰度大于1,相对于其他几个性状而言,分布比较陡峭,峰态明显,其他几个性状分布比较平缓;长光照(甘肃)的6个性状峰度均小于1,分布都比较平缓。

图2 短日照条件下的光敏相关性状分布
2.4 光敏相关性状在不同光长条件下的相关性分析
相关性分析结果显示,短日照条件下,F2∶3家系间的叶片数与株高、穗位高的相关性不显著,其余的光敏性状之间的相关性表现为显著的正相关和显著的负相关关系。
在长日照条件下,F2∶3家系的6个光敏相关性状之间都是正相关的关系,除了抽雄期与株高之间的差异不呈现显著性以外,其他的光敏性状之间都呈现出极显著正相关(表4)。
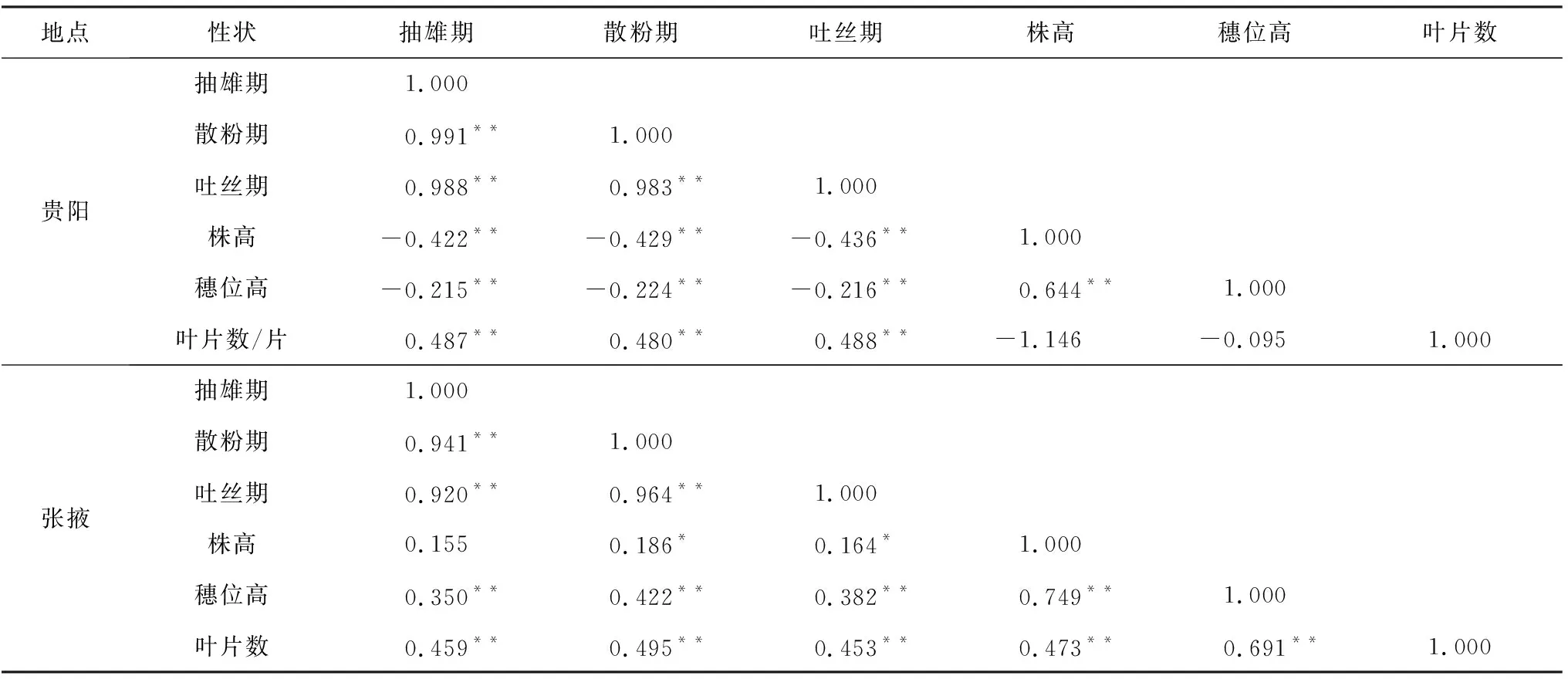
表4 光敏相关性状的相关性分析
3 结论与讨论
本研究显示,光敏相关性状在不同光长环境条件下均呈现极显著差异。说明热带、亚热带种质引种到温带后,延长日照数会导致玉米开花期延迟,雌雄不协调、营养生长旺盛,叶片数增加,晚熟,经济系数低,该结果与冯芬芬等[18]的报道相一致。无论是在张掖还是贵阳,穗位高的变异程度在6个光敏相关性状中均最大,变异系数分别为23%和22%,大于株高的16%和12%,说明穗位高的光周期敏感程度高于株高。2个光长环境下的6个光敏相关性状均呈现出连续性的正态分布,符合数量遗传的特征。同时,抽雄期、散粉期、吐丝期在2个光长环境下都表现为显著的正相关关系;随着光照长度的增加,生育期与叶片数有显著正相关,相关系数为0.459,说明生育期越长,叶片数越多,这与冯铸[19]对玉米叶片数和生育期的对应关系进行的研究分析结果相一致;株高与穗位高在不同光长环境下均表现出显著正相关,相关系数分别为0.644和0.769,反映了株高越高,穗位越高的关系,与赵强等[20]的研究一致。
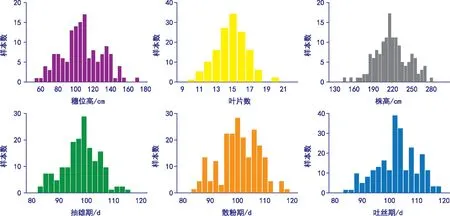
图3 长日照条件下的光敏相关性状分布
综上所述,理想的玉米生育期作为玉米育种的主要目标之一[21],抽雄期、散粉期、吐丝期是3个最重要的生育时期指标[22],生育期性状选择的好坏直接决定了育种目标能否实现。在不同纬度和海拔,应选择生育期长短适宜的品种;合理调整制种材料(父本和母本)的播期,可以使雌雄花期错开或者雌雄花相遇,提高结实率,增加制种产量[23]。通过对光敏系数的分析可以看出,不同光敏相关性状对光周期敏感程度是有差异的,生育期性状的光周期敏感程度表现为:吐丝期>抽雄期>散粉期;株型性状的光周期敏感性程度表现为:穗位高>株高>叶片数。郭国亮等[24]对热带Suwan玉米群体遗传变异的研究表明,生育期性状对光周期敏感程度为:抽雄至吐丝>散粉至吐丝>抽雄至散粉>吐丝期>散粉期>抽雄期;株型性状对光周期敏感程度为:穗位高>穗位系数>穗位叶面积>穗上叶面积>叶片数>雄穗分枝数>株高。陈彦惠等[17]对CIMMYT种质群体的不同性状的光周期敏感程度进行研究分析得出:ASI(雌雄穗开花间隔时间)>雄穗分枝数>穗位高>棒3叶叶面积>吐丝>叶片数>抽雄>散粉>株高>雄穗长度>茎粗。本研究通过对F2∶3家系的表型性状分析可以看出,株高和穗位高受环境的影响显著,这与何文昭[25]、许莹莹等[26]研究结果相同,株高和穗位高是决定玉米倒伏性、造成玉米减产的重要因素之一[27],李清超等[28]对株高、穗位高的QTL定位进行了挖掘,李凯等[27]利用穗位高系数对株高和穗位高进行阐述,一般认为,穗位高系数约等于2,即穗位高占株高的1/2。也有研究表明,穗位高系数与穗长、行粒数、千粒重、产量、穗长、行粒数、千粒重相关,可作为高产玉米杂交种的选育指标[29]。随着光照长度的增加,生育期与叶片有显著的正相关关系。大多数玉米品种随着光长增加,使雄穗延迟出现,开花期的延长,从而导致了叶片数的增加[30]。因此,在拓宽热带、亚热带种质与温带种质的育种工作中,要把生育期、株高、穗位高以及叶片数作为种质改良的重点指标,进一步提高热带、亚热带种质与温带种质利用,培育高光效玉米种质。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
作物学报(2022年8期)2022-05-29
林业科学研究(2022年1期)2022-03-26
河南科技学院学报(自然科学版)(2021年5期)2021-09-26
西北农业学报(2021年6期)2021-07-16
蔬菜(2020年4期)2020-12-14
水产科学(2020年2期)2020-03-20
森林工程(2018年1期)2018-05-14
少儿科学周刊·少年版(2017年7期)2017-09-29
