
光周期与光质对栎属幼苗生长及叶绿素荧光的影响
2022-03-26 03:44:00刘建锋江泽平常二梅赵秀莲
林业科学研究 2022年1期
姚 宁,刘建锋*,江泽平,2,常二梅,赵秀莲,谢 瑞,王 奇
(1.中国林业科学研究院林业研究所,国家林业和草原局林木培育重点实验室,北京 100091;2.中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室,北京 100091)
自然条件下,林下光环境是林下更新的重要驱动因子之一,对林下幼苗建成、生长及生物量分配具有重要影响[1-4]。同时,温室育苗过程中容易形成弱光环境,影响叶片的光合同化过程,不利于植株的形态建成[5]。因此,研究人工补光措施对林木培育有重要意义。近年来,许多学者已针对光环境对树木生长的影响做了大量的研究[6-10]。光周期作为季节性休眠的诱导因子[9],影响叶片对光合有效辐射(PAR)吸收积累的时间长度,会对物质生产产生显著影响[11];而光质以触发植物体内的光信号受体的方式,影响植物体内相关激素含量和相关酶活性[12-13],进而影响植物的物质合成、新陈代谢和生长发育等[8]。在自然光环境下,在高密度环境下生长的植物可感知入射光红光远红光比率(R∶FR)降低,这是由于冠层叶片吸收红光和来自邻近植物的远红光反射所致,而低R∶FR 会诱导植物产生避阴反应(SAS),即在冠层下方遮阴处或有较高邻近植物的环境中生长的植物在感应到低R∶FR后,表现出如茎和叶柄伸长、叶面积增加等现象[1,14-16],促其叶片投射到日光未衰减的区域。此外,植物对不同光周期或光质条件的生理响应也存在种间和种内差别[17-21]。然而,目前针对植物幼苗对光周期和光质及其交互作用机制的研究还较少,需要进一步研究来确定其对植物光合及生长的影响机理。
栎属(QuercusL.)是北半球天然分布最广的木本植物类群之一。我国拥有丰富的栎属植物资源,据第九次森林资源清查[22],我国栎类林在森林面积和蓄积量均列10 大树种组的首位;栎类树种作为我国暖温带和温带森林的重要建群种[23],在涵养水源、保持水土和维持生物多样性等方面发挥了重要功能。同时,由于栎属植物普遍具有较高的抗逆性[24],在困难立地造林和园林绿化中应用广泛[25]。然而,大量研究[26]表明,在栎类天然林分中,由于种子及幼苗有较大比例被动物取食[27],且幼苗早期生长受到林分密度[28]、光照[7]等微环境的限制,导致天然更新困难;而在人工育苗实践中,不同栎类树种幼苗的适宜受光条件尚缺乏深入研究。在栎类树种中,栓皮栎(Q.variabilis)和短柄枹栎(Q.glandulifera)分布较广,在我国大部分地区均有分布[29];蒙古栎(Q.mongolica)则主要分布在东北及华北。因此,本研究以3个落叶栎属植物为例,通过LED 人工光源的诱导,来探讨不同光周期与光质比对栎属植物幼苗的生长及叶绿素荧光参数的影响及种间和种内的响应差异,为栎属植物育苗壮苗及人工辅助更新提供科学参考。
1 材料和方法
1.1 试验材料
2018 年9—10 月,选择5 个不同纬度的栎属植物天然林分收集种子,包括3个栓皮栎种源(Qv):辽宁大连(LN,北方种源)、陕西太白(SX,中部种源)、云南安宁(YN,南方种源);1个蒙古栎种源(Qm,黑龙江加格达奇)和1个短柄枹栎种源(Qg,河南内乡)(表1)。各种源地地理气候信息见表2。将采集到的种子用60℃温水浸泡2 h 杀虫处理后阴干至表面干燥,然后将种子储存至4℃冰箱。2019 年春,选择健康且大小均一的种子进行催芽以缩短发芽时间。发芽后于同一天播种于定制育苗容器(内径7 cm,深度36 cm)内预培养2 个月,每容器播1 粒,培养基质为珍珠岩。预培养期间,每3 日浇1 次1/2 倍浓度的Hoagland 标准营养液来提供幼苗生长所需养分,同时观测生长动态。试验在中国林科院科研温室中进行(40°0′10′′ N,116°14′38′′ E),温室昼夜温度约为25/20℃,湿度约为70%/85%,透光度为50%~60%。

表1 主要缩略词中英对照Table 1 List of main abbreviations
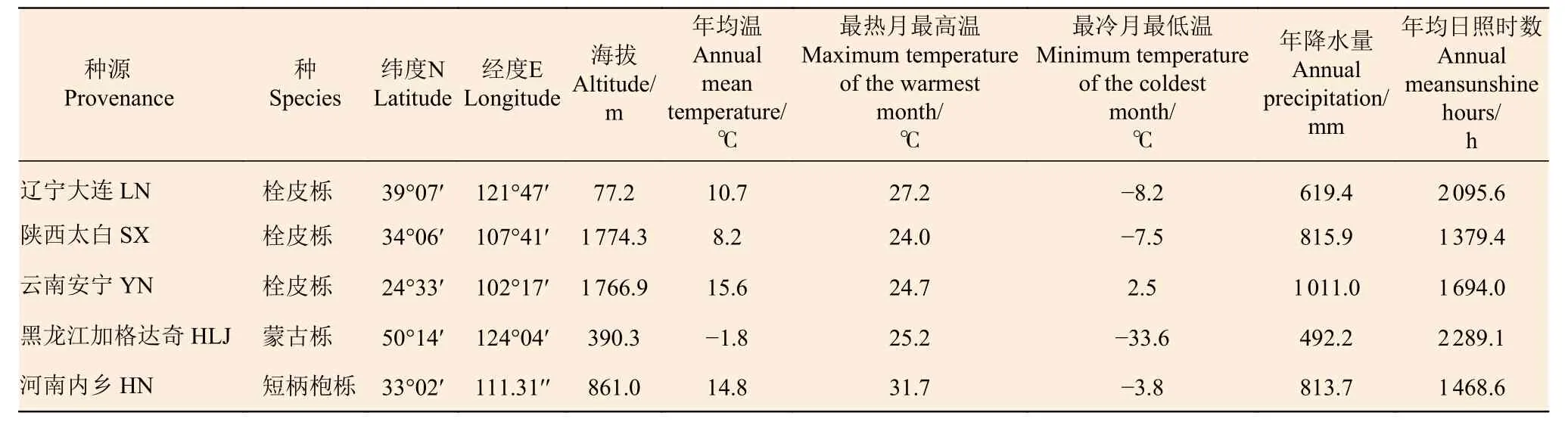
表2 各栎属种的种源地地理位置与气候概况Table 2 Geographic location and climate characteristics of different provenances
1.2 试验设计
将5 个种源幼苗分别进行不同光周期处理[日夜时长分别为10 h/14 h(短光周期,S),14 h/10 h(中光周期,M),18 h/6 h(长光周期,L)],并照射不同红光/远红光比(R :FR)的光照[分别为0.83 ± 0.04(低光质比,D)、1.37 ± 0.04(正常光质比,N)、4.69 ± 0.04(高光质比,H)],不同光质比(R :FR)的值是基于Ballaré[30]等和Smith[31]等研究确定。每个处理每个种(源)随机分配4 株幼苗,3 次重复。通过智能定时开关(GND-1,Bull,China)来控制不同照光时长。不同的光质比由220 W 的定制LED 补光灯(APO,深圳市联邦重科电子科技有限公司)来实现,每种补光灯由不同比例的白光灯珠、红光灯珠、远红光灯珠组合而成。白光灯珠为全光谱,红光(R)灯珠波长为660 nm,远红光(FR)灯珠波长为730 nm。用照度计(LI-250A,LI-COR,USA)测得低光质比(D)和正常光质比(N)的PAR 均为513 ± 3 μmol·m-2·s-1(在植株高度);高光质比(H)下,PAR 为495 ± 3 μmol·m-2·s-1。在高光质比(H)下,由于补光灯光强略低,灯的位置略微降低以达到相同PAR。不同处理之间用四面包围的遮光布隔开,顶部设置补光灯,留有通风口,底部为育苗容器,容器放置在透气的铁丝网苗床上。
1.3 测定指标及方法
1.3.1 相对生长速率(RGR)利用游标卡尺(0.01 mm)和直尺(0.1 cm)对每株幼苗进行2 次基径(cm)和株高(cm)的测量,每处理测3 个重复,两次测量间隔15 天。进而计算相对生长速率(RGR),计算公式如下[32]:
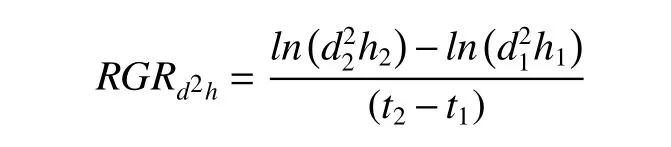
其中,d1、h1、t1分别为第1 次测量时幼苗的基径、株高和测量时间,d2、h2、t2分别为第2 次测量时幼苗的基径、株高和测量时间,t2-t1为2 次测量的时间间隔(月)。
1.3.2 叶绿素荧光参数 选择长势良好的幼苗暗适应20 min 后,用FluorPen-FP100 手持式荧光仪(Photon System Instruments,Drasov,Czech Republic)测定叶片的OJIP 曲线,OJIP 曲线由3 000 μmol·m-2·s-1的脉冲 红光诱 导,每处理 测3 个 重复。按照每次重复OJIP 曲线各时间点的荧光强度求其平均值,按照平均值绘制OJIP 曲线,曲线横坐标以对数形式表示。其中20 μs 测量的荧光强度为O 相荧光强度(Fo),2 ms 测量的荧光强度J 相荧光强度(Fj),30 ms 测量的荧光强度I 相荧光强度(Fi),最后达到最大荧光强度P 相(Fm)。通过JIP-test 分析并计算得出叶片的PSII 最大光化学效率(Fv/Fm)、光合性能指数(PIABS)、单位反应中心活性(ABS/RC)等参数[33]。
1.4 数据分析
所有数据用RStudio 软件(Version 1.3.109 3,http://www.rstudio.com)进行三因素(种源、光周期和光质)方差分析、多重比较,以及显著性检验(α=0.05),用RStudio 及OriginPro 2019b 制图。
2 结果与分析
2.1 相对生长速率(RGR)
栓皮栎的不同种源间(指栓皮栎种源辽宁大连LN、陕西太白SX、云南安宁YN 之间,以下简称为“种源间”)的RGR 有显著差异(p< 0.05)(表3)。光周期对RGR 有显著影响,短光周期S 下的RGR 分别比中光周期M、长光周期L 低8.11%、37.84%(p< 0.05)(图1)。3 个种源的RGR 均随着光周期延长而呈现增加的趋势,种源LN 的RGR 则在中光周期M 下最高。光质比处理对种源间RGR 无显著影响(p> 0.05)。
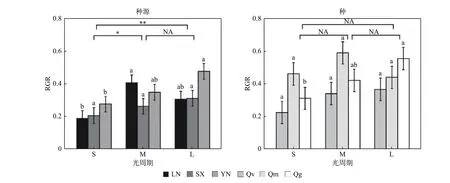
图1 相对生长速率(RGR)对不同处理的响应Fig.1 Response of relative growth rate (RGR) to different treatments
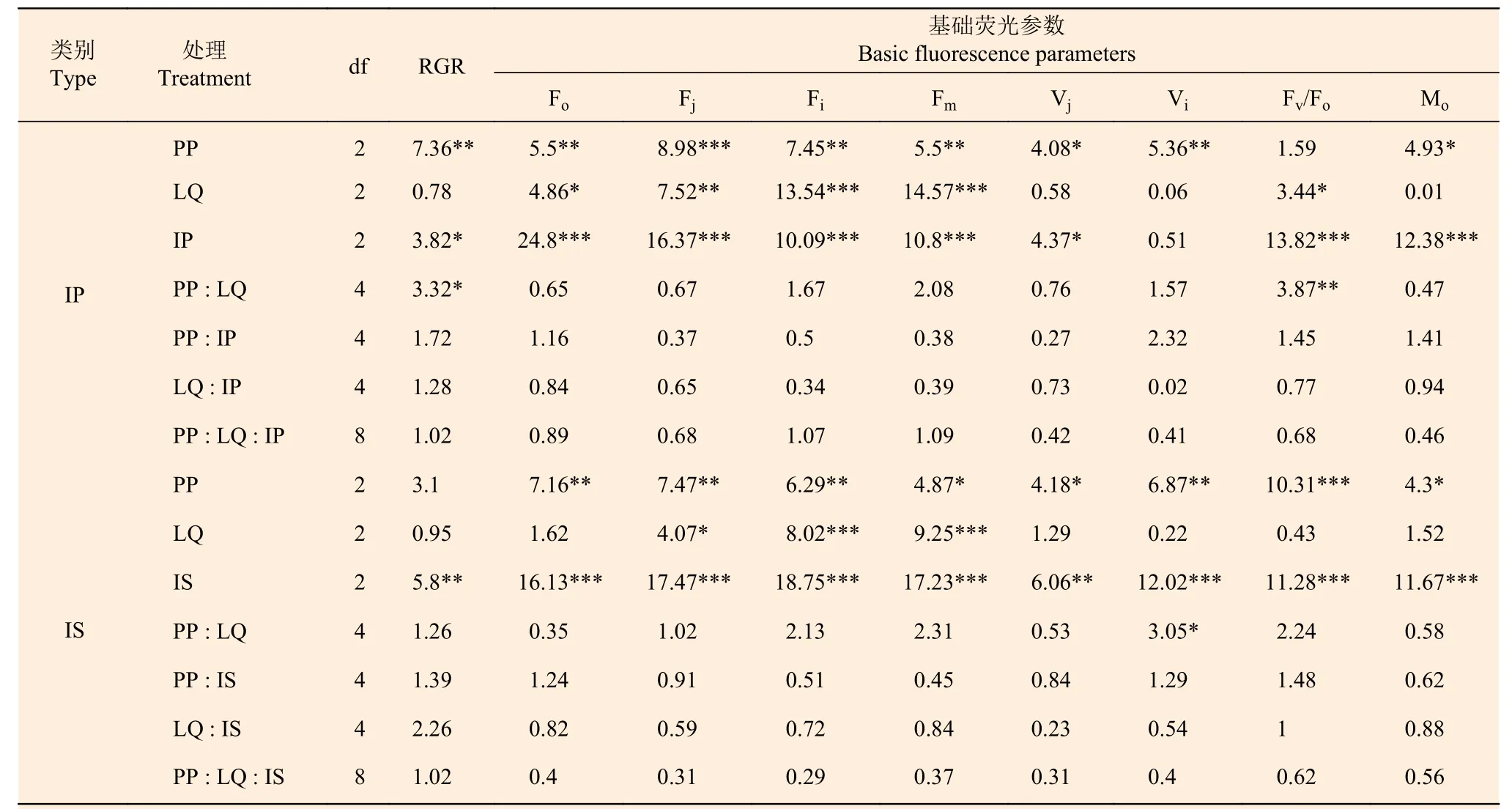
表3 光周期、光质比及不同种源/种对RGR 与基础荧光参数的三因素方差分析Table 3 Two-way ANOVA on RGR and basic fluorescence parameters by photoperiod,light quality ratio and different provenances/species(F value)
3 个栎属植物种间(指栓皮栎Qv、蒙古栎Qm、短柄枹栎Qg 之间,以下简称为“种间”;栓皮栎Qv 数据为栓皮栎3 个种源LN、SX、YN 数据取均值)的RGR 有显著差异(p< 0.05)(表3)。光周期对种间RGR 无显著影响,光周期S 下的RGR 虽然分别比光周期M、L 低26.67 %、26.67 %但差异不显著(图1)。光周期处理对栓皮栎Qv、蒙古栎Qm 的RGR 无显著影响,但短柄枹栎的RGR 随光周期延长而升高。光质比处理对种间RGR 无显著影响(p> 0.05)。
光周期与光质比的交互效应仅在栓皮栎种源间显著(表3)。在中光周期M 下,低光质比D 的RGR 最高,且显著高于正常光质比;但短光周期S 与长光周期L 下,均是正常光质比N 下RGR 最高(图2)。
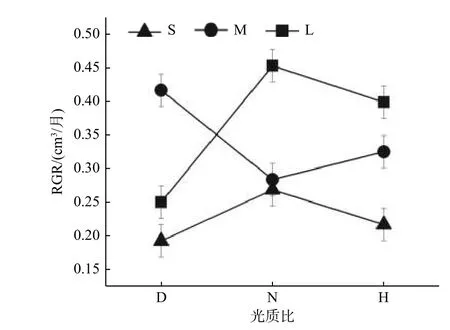
图2 栓皮栎种源间不同光周期幼苗的RGR对光质比处理的响应Fig.2 Response of RGR of Qv seedlings under different photoperiods to light quality ratios
2.2 叶绿素荧光诱导动力学
2.2.1 OJIP 曲线及基础荧光参数 光周期、光质比以及种源对O、J、I、P4 个拐点的荧光强度(Fo、Fj、Fi、Fm)均有显著或极显著影响,但3 个因素的交互作用均不显著(表3)。光周期处理中,中光周期M 的Fo、Fj、Fi、Fm值显著高于长光周期L(p< 0.05);光质比处理中,低光质比D 下Fj、Fi、Fm均显著高于正常光质比N 和高光质比H(p< 0.05)。在种间,除光质比处理对初始荧光(Fo)无显著影响外,光周期、光质比以及种对O、J、I、P4 个拐点的荧光强度(Fo、Fj、Fi、Fm)均有显著或极显著影响,但3 个因素的交互作用均差异不显著(表3)。由于篇幅限制,仅展示部分处理组合下(固定两个因素,对第三个因素进行比较)的OJIP 曲线图(见图3)。
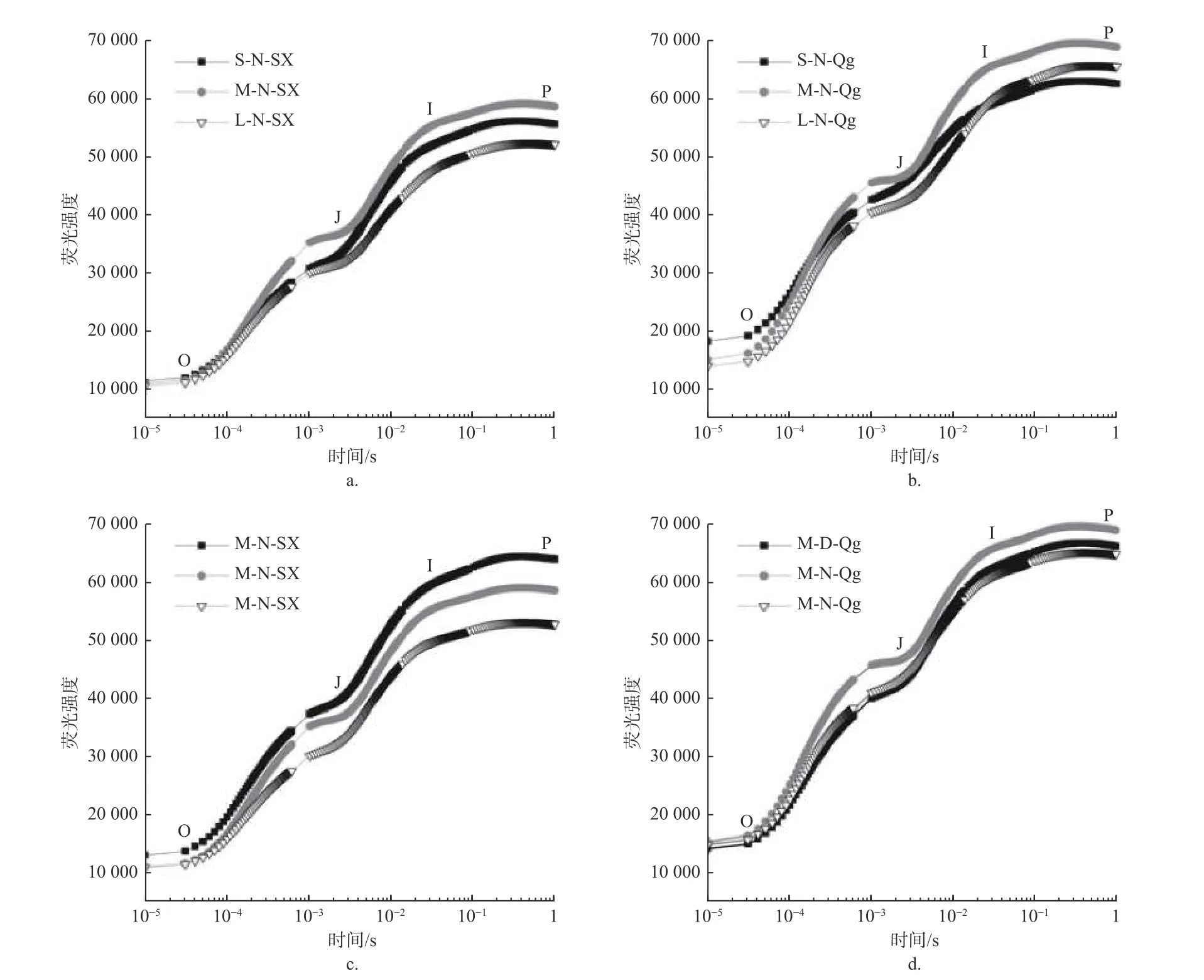
图3 不同处理下的OJIP 曲线Fig.3 OJIP curve under different treatments
在种源水平,光周期处理主要影响Vj、Vi和Mo,如中光周期M 处理下的Mo、Vi、Vj分别比长光周期L 高10.34 %、2.25 %、6.25 %(p< 0.05)(图4)。光质比处理主要影响Fv/Fo值,正常光质比N 处理下的Fv/Fo比高光质比H 高5.41 %(N:3.51;H:3.48)。
光周期处理在种间对基础荧光参数均有显著影响,中光周期M、长光周期L 处理下的Fv/Fo显著高于短光周期S(S:3.06;M:3.37;L:3.41),短光周期S 下Mo(S:1.36;L:1.24)、Vi(S:0.91;L:0.9)、Vj(S:0.53;L:0.5)均显著高于长光周期L(p< 0.05),中光周期M 下的Vi也显著高于长光周期L(M:0.91;L:0.9)(图4)。光质比对Vj、Vi、Fv/Fo以及Mo作用均不显著。
在种源间(图4),南方种源YN 在长光周期L 下的的Fv/Fo、Fv/Fm显著高于短光周期S(p<0.05),Mo显著低于短光周期S(p< 0.05),中光周期M 下Vi最高。但种源LN、SX 在不同光周期下Fv/Fo、Fv/Fm、Mo、Vi、Vj均差异不显著(p>0.05)。各种源幼苗的Fv/Fo、Fv/Fm、Mo、Vi、Vj对光质比处理的响应均不显著。
在种间(图4),短柄枹栎的Fv/Fo、Fv/Fm在短光周期S 下均显著低于光周期M、L,Vj、Vi显著高于光周期L;但栓皮栎的Fv/Fo、Fv/Fm、Mo、Vj对光周期处理响应均不显著,Vi在中光周期M 下最高。蒙古栎的Fv/Fm、Mo、Vi、Vj对光周期处理响应均不显著,Fv/Fo在短光周期S 下显著低于光周期M、L。3 个栎属种的Fv/Fo、Fv/Fm、Mo、Vi、Vj对光质比处理的响应均不显著。
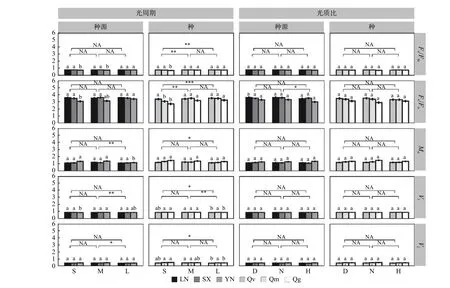
图4 部分基础荧光参数对不同处理的响应Fig.4 Response of some basic fluorescence parameters to different treatments
2.2.2 PSII 反应中心活性 光周期处理对种源间各能量分配参数无显著影响,但对Ψo、φEo有显著影响(p< 0.05)。长光周期L 的Ψo(S:0.51;M:0.49;L:0.52)、φEo(S:0.39;M:0.38;L:0.41)显著高于中光周期M(p< 0.05)(图5)。光质比处理对种源间各能量分配参数及量子产额参数均无显著影响。
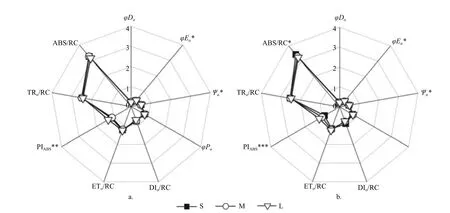
图5 不同光周期处理下PSII 反应中心活性和电子传递的变化Fig.5 Changes of PSII reaction center activity and electron transfer under different photoperiod
除ETo/RC 以及TRo/RC 外,光周期对种间各能量分配参数及量子产额参数均有显著影响(p<0.05)。短光周期S 下ABS/RC(S:3.43;M:3.31;L:3.24)、DIo/RC(S:0.9;M:0.78;L:0.76)显著高于长光周期L(p< 0.05),虽也高于中光周期M 但差异不显著。长光周期L 的Ψo(S:0.47;M:0.48;L:0.5)、φEo(S:0.35;M:0.37;L:0.39)显著高于短光周期S(p< 0.05)。中光周期M、长光周期L 的φDo显著低于短光周期S(S:0.25;M:0.23;L:0.23)(p< 0.05)。此外,长光周期L 的PIAbs显著高于短光周期S 和中光周期M(S:0.87;M:1.02;L:1.18)。光质比处理对种间的各能量分配参数及量子产额参数均无显著影响。
3 讨论
光周期作为环境信号因子激活植物体内的不同信号途径的发生,同时通过影响叶片光合作用制造同化物的时长来影响同化物合成和分配[34-35]。本研究结果表明,延长光周期可提高栎属植物的相对生长速率RGR,这与前人关于光周期[36-37]的研究结果一致。如朱开元等[37]的研究也表明,延长光周期显著增加了罗汉松(Podocarpus macrophylus(Thunb.)D.Don)和鸡爪槭(Acer palmatumThunb.)的株高,表明光合作用制造的同化物主要用在了植物茎的高生长上;缩短光周期会导致干物质积累不足,导致幼苗高生长缓慢。光质比处理对栎属植物幼苗RGR 均无显著影响。但在中光周期下,低光质比的幼苗RGR 最高,且显著高于正常光质比,表明避阴反应(即低光质比会促进茎伸长的现象)并非在任何环境下均有效,在本实验中仅在中光周期下表现出了明显的避阴反应,缩短、延长光周期均会使这一效应不明显,进一步说明光周期对避阴反应存在介导效应。这可能是由于短光周期下干物质积累不足,无法供应茎伸长所需养分,而长光周期下茎生长速度均已达到较高水平,无需光质比信号的诱导就有充足的养分供应高生长。郑芬等[4]通过监测天然林林下光照及幼树株高等指标,也发现R/FR 变化对林下幼树的高径比无显著影响,认为复杂的天然林生境下植株高生长会更多地受到其他因素(如凋落物和种子特性等)的影响[38]。因此,虽然许多研究[1,39-42]表明植物存在避阴反应,但不同植物对其敏感度不同,同种植物在不同环境条件下(如光周期、凋落物等)也会对避阴反应有不同的响应。
荧光的快速上升反映了光合电子传递链中几种不同氧化还原反应的动力学所影响的的浓度。因此,OJIP 瞬态可用于表征PSII 光化学的量子产率和电子传递活性[43]。本研究中,延长光周期会导致栎属植物幼苗的Fo、Fj、Fi、Fm值的降低,表明延长光周期会影响光合机构的功能,比如增加反应中心复合体的数量、增加QA到QB的电子传递速率等。φPo(Fv/Fm)和PIABS是研究植物光合生理状态的重要指标,Fv/Fm反映了植物叶片PSII 原初光能转化效率[43],PIABS可从光能吸收、捕获和电子传递3 方面综合反映光系统II 的活性[44]。本研究中,除缩短光周期会显著降低栎属植物种水平的Fv/Fm外,其他光周期处理下Fv/Fm均变化不明显,表明缩短光周期会阻碍叶片转化能力。延长光周期会提高PIABS,表明延长光周期会增强PSII 反应中心活性。Mo、Vj、ψo、φEo等参数可以反映出叶片PSII 供体侧变化[45]。例如,Vj可以反映QA到QB的积累量[46-48]。本研究发现,延长光周期处理会使得Vj和Mo明显降低,说明QA被还原的速率下降,积累量减少甚至无积累。延长光周期会促进PSIIQA向QB的电子传递更通畅。缩短光周期导致φDo增加,说明缩短光周期会增加热耗散比例,降低PSII 电子传递速率。与此同时,延长光周期造成ψo、φEo增加,这进一步促进了PSII 中的电子传递。在种水平上的ABS/RC、DIo/RC随着光周期的延长逐渐降低,但光周期处理对TRo/RC、ETo/RC 均无显著影响,表明随着光周期的延长,单位反应中心吸收的光能逐渐降低,但光合系统会通过降低热耗散的方式,优先保障传递到QA的能量以及QA传递到QB的能量,来维持光合作用的能力。李冬梅等[49]和刘杰等[50]的研究也表明,缩短光周期会降低φPo、ψo、PIABS,这与本研究发现的变化规律一致。综上,叶片的光合过程对光周期较为敏感,随着光周期延长,QA被还原的速率下降,积累量减少甚至无积累,PSIIQA向QB的电子传递更通畅,增加PSII 电子传递速率。虽然单位反应中心吸收的光能逐渐降低,但光合系统通过降低热耗散,优先保障传递到QA的能量以及QA传递到QB的能量,来维持光合作用的能力。
有研究认为在植物对干旱等外界环境变化不敏感时,Fo和Fm可能发生相同趋势的变化,进而维持Fv/Fm值不变[51],这与本研究的结果一致。降低光质R/FR 比值会导致不同种源栓皮栎幼苗的Fo、Fj、Fi、Fm值的升高,Fv/Fm则无显著差异。因此可以认为在种源水平上叶片对光质比变化不敏感。光质比处理对栎属植物幼苗的能量分配参数及量子产额参数均无显著影响,表明光质比处理并未显著影响光合作用的结构。总体上,本研究中南部分布的种(短柄枹栎和栓皮栎)或种源(如栓皮栎南方种源YN)相比北方分布的种(如蒙古栎)或种源对光周期处理更敏感,延长光周期会显著提高南方种或种源的Fv/Fo、Fv/Fm,增加PSII 光能转换效率和潜在活性,促进光系统线性电子传递,进而提高光合同化效率。这与本课题组前期的研究结果相吻合[52],即当栓皮栎南方种源(适应于当地短日照环境)移植到北方环境(长日照)后,其羧化效率、光化学猝灭系数和相对生长速率均显著高于南方环境。
4 结论
本研究通过对栎属植物幼苗生长及荧光参数对光周期和光质比的响应分析,发现其对光周期和光质比存在种内和种间差别。栓皮栎南方种源对光周期变化响应较之中部与北部种源更加敏感,延长光周期会显著增加南方种源的Fv/Fo、Fv/Fm及RGR。在种间水平上,短柄枹栎和栓皮栎较之蒙古栎对光周期的响应更强烈,缩短光周期会显著降低短柄枹栎的Fv/Fm及RGR,延长光周期显著提高了栓皮栎和短柄枹栎的RGR,但蒙古栎的Fv/Fm及RGR 对光周期的响应均不显著。R/FR 比值降低会导致栎属植物幼苗的Fo、Fj、Fi、Fm值同步升高,但Fv/Fm不变,对各能量分配参数、量子产额参数及RGR也无显著影响。栎属植物幼苗总体对光质比处理不敏感。此外,光质比与光周期对RGR 存在显著交互效应。在中光周期下,幼苗表现出明显的避阴反应,即低光质比下RGR 最高,且显著高于正常光质比和高光质比。
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
南方农业·下旬(2020年1期)2020-06-01 07:47:32
现代园艺(2019年24期)2019-01-07 01:19:36
现代农业科技(2009年21期)2018-09-03 00:00:00
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
浙江农业学报(2017年1期)2017-05-17 06:13:36
