
外源添加乙偶姻对酿酒酵母产2,3-丁二醇及其菌株的影响
2021-01-15 08:27吕雨泽邓利廷葛菁萍
中国农学通报 2021年2期
张 弛,吕雨泽,邓利廷,孙 健,葛菁萍
(1黑龙江大学农业微生物技术教育部工程研究中心,哈尔滨 150500;2微生物黑龙江省高校重点实验室/黑龙江大学生命科学学院,哈尔滨 150080)
0 引言
2,3-丁二醇(2,3-BD)是一种应用化合物,被作为一种重要的化工原料,并且2,3-BD被航天、能源以及食品等多个领域广泛应用[1]。通常人们利用微生物以石油为底物的方式进行生产2,3-BD被广受欢迎,然而,现如今2,3-BD的生产含有不足之处:一是产量较低,二是2,3-BD的结构十分特殊,利用化学法合成2,3-BD的成本很高,所以不能用于工业化大规模生产[2]。近年来利用生物法来生产2,3-BD等产品受到人们的关注,同时在国内外其研究与发展速度飞快[3],优势在于其成本低廉,工艺技术简单,同时具有降低污染,保护环境的作用[4-5]。
在大自然中,有许多常见的菌种可以用来生产2,3-BD,包括克雷伯氏菌属(Klebisellasp.)[6-7]、芽孢杆菌属(Bacillussp.)[8-9]以及酵母菌属(Saccharomycessp.)[10]等。不幸的是,克雷伯氏菌属和芽孢杆菌属等被归类为II类致病微生物,容易造成具有危害性的工业生产[11-12]。相比之下,酿酒酵母已经被作为安全性较高的微生物,且具有2,3-BD生产能力[13-14]。这使得研究学者们将酿酒酵母作为生产2,3-BD的焦点。
但是,野生型酿酒酵母本身产生2,3-BD的能力有限,如果不加以人为干涉的话,还不具备进行大规模生产的潜力。因此人们从改造酿酒酵母代谢途径的角度出发,试图通过阻断其支路代谢途径[15]、过表达关键酶基因的方式[16],来提高2,3-BD的产量。在酿酒酵母细胞中,2,3-BD的合成途径其一是通过丙酮酸在丙酮酸脱羧酶(pyruvate decarboxylase/PDC)的作用下生成乙醛,乙醛在PDC催化下生成乙偶姻,最终乙偶姻在2,3-丁二醇脱氢酶(2,3-butanediol dehydrogenase/BDH)作用下生成2,3-BD。所以,乙偶姻成为2,3-BD合成途径中重要的前体物质[17-18]。本课题组前期已经尝试对丙酮酸脱羧酶1和丙酮酸脱羧酶5基因进行了敲除,导致2,3-BD的产量明显提升。那么,外源添加适当浓度的物质乙偶姻后,是否会继续提高2,3-BD的产量以及菌株的生长?本研究将利用上述的两种突变株以及S.cerevisiaeW5和S.cerevisiaeW141原始菌株对此问题进行探究。
1 材料与方法
1.1 菌株与培养基
1.1.1 试验菌株S.cerevisiaeW5和S.cerevisiaeW141,由黑龙江大学微生物重点实验室保藏。S.cerevisiaeW5-Δpdc1/Δpdc5,经过前期构建,其中W5-Δpdc1和W5-Δpdc5分别是敲除了丙酮酸脱羧酶1和丙酮酸脱羧酶5的基因工程菌株。
1.1.2 培养基 YPD液体培养基:蛋白胨20 g/L,酵母提取物10 g/L,葡萄糖20 g/L,pH自然,108°C高压灭菌20 min;葡萄糖单碳源发酵培养基(g/L):葡萄糖80 g/L,YNB 3.4 g/L,蛋白胨 20 g/L,K2HPO43 g/L,KH2PO411.8 g/L,(NH4)2SO410 g/L,pH自然,108°C高压灭菌20 min。
1.2 方法
1.2.1OD值与细胞干重二者关系确定 将培养至对数期的4株菌株,接种到培养基中培养12 h,30°C,140 r/min,在600 nm处吸光度下,测定吸光值。根据吸光值,进行梯度稀释,将菌液稀释成6个不同的浓度,进行离心后,弃上清,加入适量无菌水,在再行离心处理,将样品放入烘箱中烘干称重。
1.2.2 乙偶姻最适添加量的确定 将培养至对数期的S.cerevisiaeW5/W5-Δpdc1/W5-Δpdc5/W141菌株,进行接种培养,外源添加0、2、4、6、8、10、12、14 g/L乙偶姻至培养基中培养84 h,30°C,140 r/min,每12 h取样,测定pH值、细胞干重(Dry Cell Weight,DCW),利用高效液相色谱法(HPLC)对葡萄糖的消耗量、2,3-丁二醇产量以及乙偶姻剩余量进行检测,进一步确定添加乙偶姻的最适量。
2 结果与分析
2.1 外源添加乙偶姻方案的确立及候选菌株的确定
通过对S.cerevisiaeW5/W5-Δpdc1/W5-Δpdc5/W141乙偶姻产量和BDH酶活进行检测(图1),发现S.cerevisiaeW5-Δpdc5的乙偶姻积累量和BDH酶活性均明星呈现先增加后降低的趋势,并且在发酵60 h时达到最大值,此时的乙偶姻积累量为0.626±0.04 g/L,BDH酶活性为0.0257±0.003 U/mg。然而,原始菌株S.cerevisiaeW5的乙偶姻产量为0.110±0.02 g/L,BDH酶活性为0.0099±0.001 U/mg,同时S.cerevisiaeW5-Δpdc5的乙偶姻产量是野生菌株S.cerevisiaeW141产量(0.282±0.02 g/L)的2.2倍,BDH酶活性两者一致。另外,研究发现S.cerevisiaeW5-Δpdc1的乙偶姻积累量呈现先上升后下降再上升的微弱趋势,在发酵24 h时积累量最大为0.129±0.01 g/L;BDH酶活性同样在24 h时达到最大值为0.0122±0.001 U/mg;同原始菌株S.cerevisiaeW5相比,乙偶姻积累量提高了17.3%,BDH酶活提高了23.2%,差异明显。然而,野生菌株S.cerevisiaeW141的乙偶姻产量和BDH酶活性均高于S.cerevisiaeW5-Δpdc1。综上所述,从乙偶姻产量和BDH酶活性随发酵时间变化的趋势可以看出,乙偶姻的量直接影响了BDH酶的活性,同时间接影响了2,3-BD的产量,最终研究确定外源添加乙偶姻的方案。
以上数据表明:第一,S.cerevisiaeW5-Δpdc1/Δpdc5的2,3-BD产量较比原始菌株S.cerevisiaeW5的都有所提高,并且乙偶姻的积累量也会提高,同时促进了BDH酶的活性;第二,通过对S.cerevisiaeW141和S.cerevisiaeW5-Δpdc1/Δpdc5的比较发现 ,S.cerevisiaeW141自身对乙偶姻的积累和BDH酶活性的作用,都是其作为2,3-BD出发菌株主要的优势。综上所述,针对于S.cerevisiaeW5/W141的优势和S.cerevisiaeW5-Δpdc1/Δpdc5的不足之处,研究确定使用S.cerevisiaeW5/W141共同作为本实验生产2,3-BD的出发菌株。
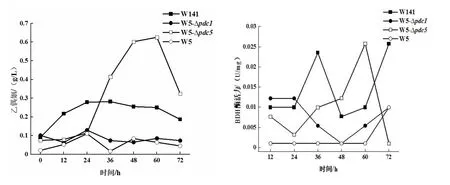
图1 乙偶姻产量和BDH酶活检测结果
2.2 OD值与细胞干重关系曲线
利用OD600nm值来测定菌体生物量,按照1.2.1的方法。根据OD600nm值与DCW具体关系分别为公式(1)~(2),结果证明,S.cerevisiaeW5/W141菌株OD600nm值和细胞干重成正比例关系。
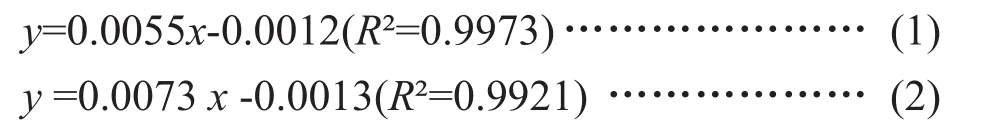
2.3 乙偶姻最适添加量的确定
将菌株接种到发酵培养基中,外源添加0、2、4、6、8、10 g/L和0、2、4、6、8、10、12、14 g/L乙偶姻,30°C,140 r/min振荡培养72 h,每隔12 h取样,葡萄糖消耗情况(图2)、2,3-BD产量和乙偶姻剩余量随发酵时间的变化情况(图3)。
由图2可以看出,外源添加不同浓度的乙偶姻对S.cerevisiaeW141/W5,在代谢过程中,葡萄糖随着发酵时间的增加,而逐渐被消耗,则在24 h左右,葡萄糖均基本耗尽。

图2 S.cerevisiae W5/W141的葡萄糖消耗量随发酵时间变化的关系曲线
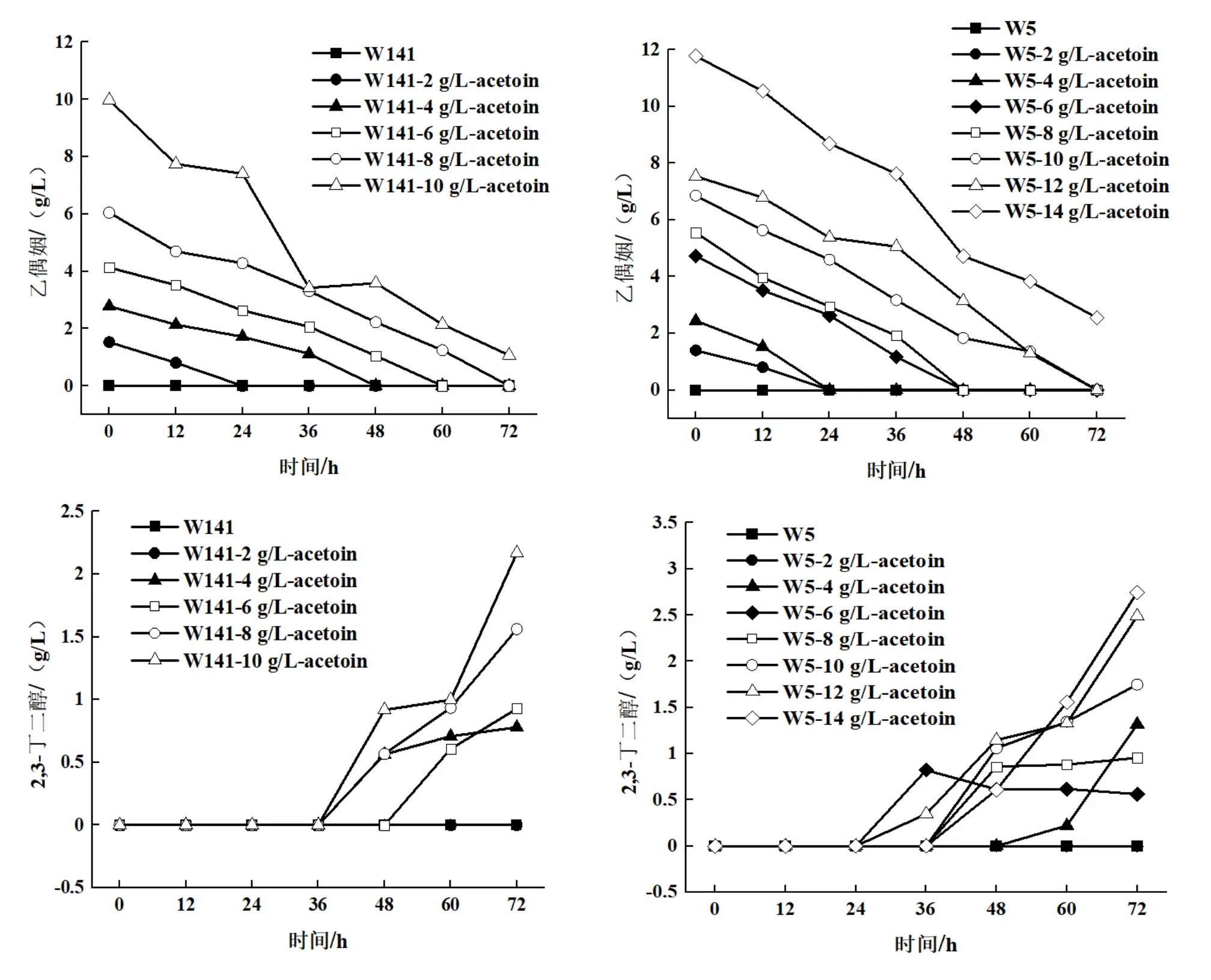
图3 S.cerevisiae W5/W141的乙偶姻剩余量和2,3-丁二醇产量随发酵时间变化的关系曲线
由图3可以看出,外源添加不同浓度的乙偶姻对S.cerevisiaeW5/W141来说,2,3-BD的产量随着乙偶姻添加量的增加而不断提高。对于S.cerevisiaeW141而言,当外源添加8 g/L乙偶姻时,2,3-BD的产量达到1.56±0.04 g/L(72 h),乙偶姻在72 h时消耗完全;当外源添加10 g/L乙偶姻时,2,3-BD的产量达到2.17±0.01 g/L(72 h),但乙偶姻并没有完全被消耗,72 h时积累了1.07±0.01 g/L。对于S.cerevisiaeW5而言,当外源添加12 g/L乙偶姻时,2,3-BD的产量达到2.49±0.02 g/L(72 h);当外源添加14 g/L乙偶姻时,2,3-BD的产量达到2.75±0.03 g/L(72 h),但乙偶姻仍然没有完全被消耗,72 h时积累了2.55±0.04 g/L。因此,从乙偶姻积累量上考虑,向S.cerevisiaeW5/W141分别添加12 g/L和8 g/L乙偶姻更适合作为后续试验的最适添加浓度。
2.4 乙偶姻对菌体活性的影响,通过菌体活性辅助说明代谢流向改变的菌体作用
2.4.1S.cerevisiaeW141外源添加8 g/L乙偶姻对的菌体活性的影响 运用细胞活性染色试剂PI对S.cerevisiaeW141/W141-8 g/L菌体进行单染色。将不同时间段的荧光效果利用流式细胞仪对其检测,FCM图谱进行荧光差异比较(图4)。通过对横坐标(PI单染色)与纵坐标(侧向角散射)的对比来对菌体活性情况进行分析(图5)。
从图中可知,随着发酵时间的延长PI染色比例有所提高,并且所有数据都相对集中,S.cerevisiaeW141/W141-8 g/L的变化趋势大致相同,均在60 h时达到最大分别为1.3%和1.9%,然而,对于流式细胞仪每次所收集的一万个细胞而言,整体细胞活性几乎没有受到影响。
2.4.2S.cerevisiaeW5外源添加12 g/L乙偶姻对的菌体活性的影响 运用细胞活性染色试剂PI对S.cerevisiaeW5/W5-12 g/L菌体进行单染色。将不同时间段的荧光效果利用流式细胞仪对其检测,FCM图谱进行荧光差异比较(图6)。通过对比从而分析菌体活性情况(图7)。
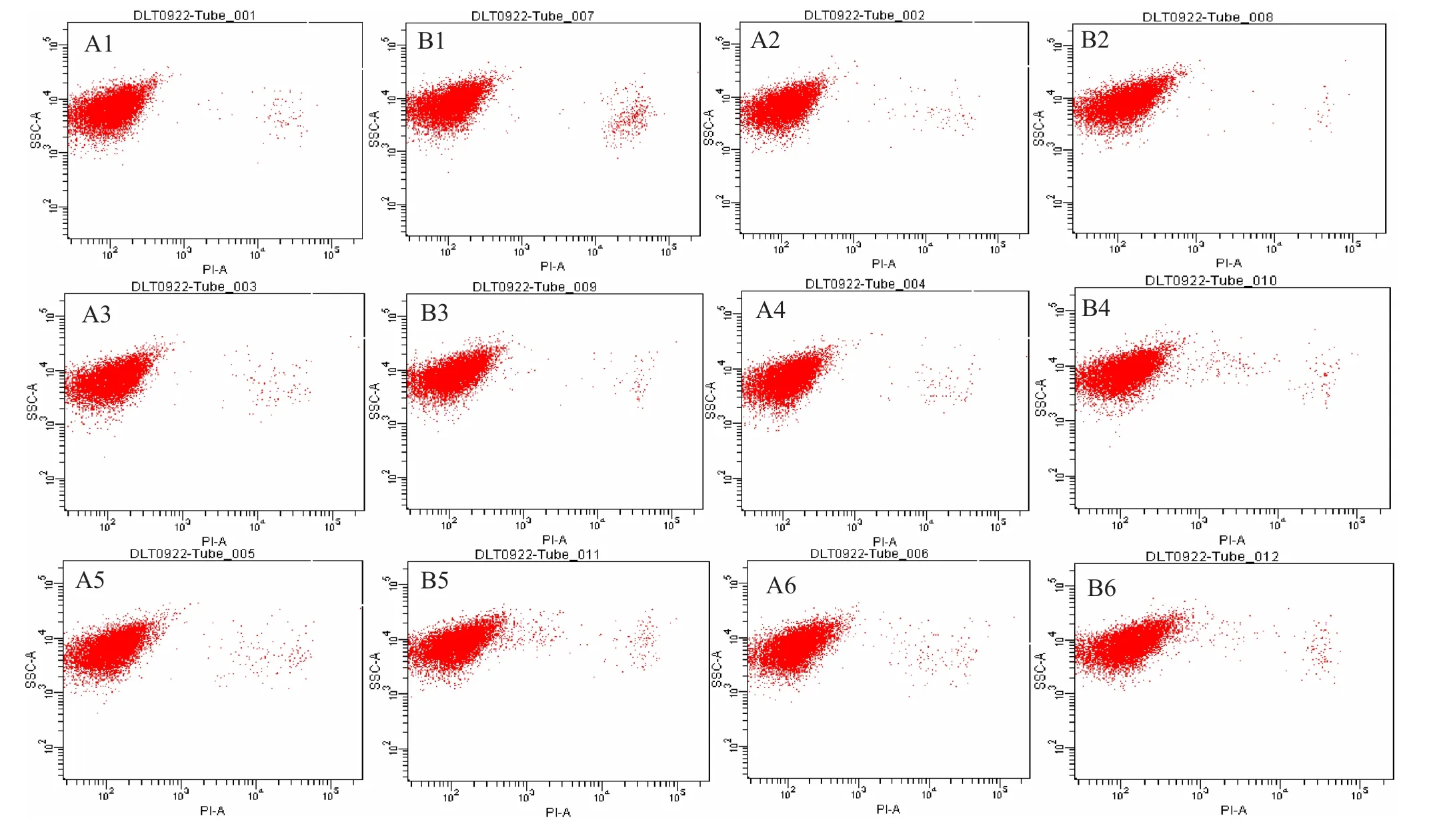
图4 不同时间段流式细胞术检测结果
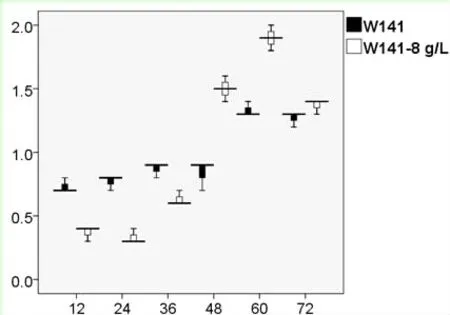
图5 流式细胞术检测结果
从图中可知,随着发酵时间的延长PI染色比例呈现先上升后下降再上升的趋势,所有数据都不太集中,与S.cerevisiaeW141/W141-8 g/L的变化趋势大致相同,均在24 h时达到最大分为0.6%和0.9%。
2.5 电镜观察菌体形态
将处理后的样品通过扫描电镜观察菌体外貌形态(图8)。可以看出,当S.cerevisiaeW141/W5在外源添加适量乙偶姻后,菌体表面圆滑平整,整体饱满,无明显变化。
3 结论与讨论
本研究前期通过尝试对丙酮酸脱羧酶的基因pdc1和pdc5进行敲除,在外源添加乙偶姻后,最终导致2,3-BD的产量不高。Yu等[19]敲除S.cerevisiae中的adh1、adh3和adh5基因后进行分批发酵,2,3-BD的产量达到2.29 g/L,与野生型S.cerevisiae相比,乙醇的产量降低31.25%,而2,3-BD的葡萄糖转化率显著提高,为0.113 g/g。但弊端是Δadh1、Δadh3、Δadh5会积累大量的毒性物质乙醛,且还会生成一定量的乙醇[20]。而前期试验结果2,3-BD的产量不高,可能是因为敲除了中心代谢的关键酶,所以,当2,3-BD代谢流增强时,菌株产能却减少了。因为在酿酒酵母中,乙醛和丙酮酸转化成乙偶姻过程中,丙酮酸脱羧酶在其中起关键作用,所以当丙酮酸脱羧酶基因被敲除后,导致关键酶的酶活降低,使乙醛和丙酮酸无法更多地转化成乙偶姻,最终导致菌株产2,3-BD的能力下降[21]。所以在2,3-BD合成过程中,关键酶起着至关重要的作用。
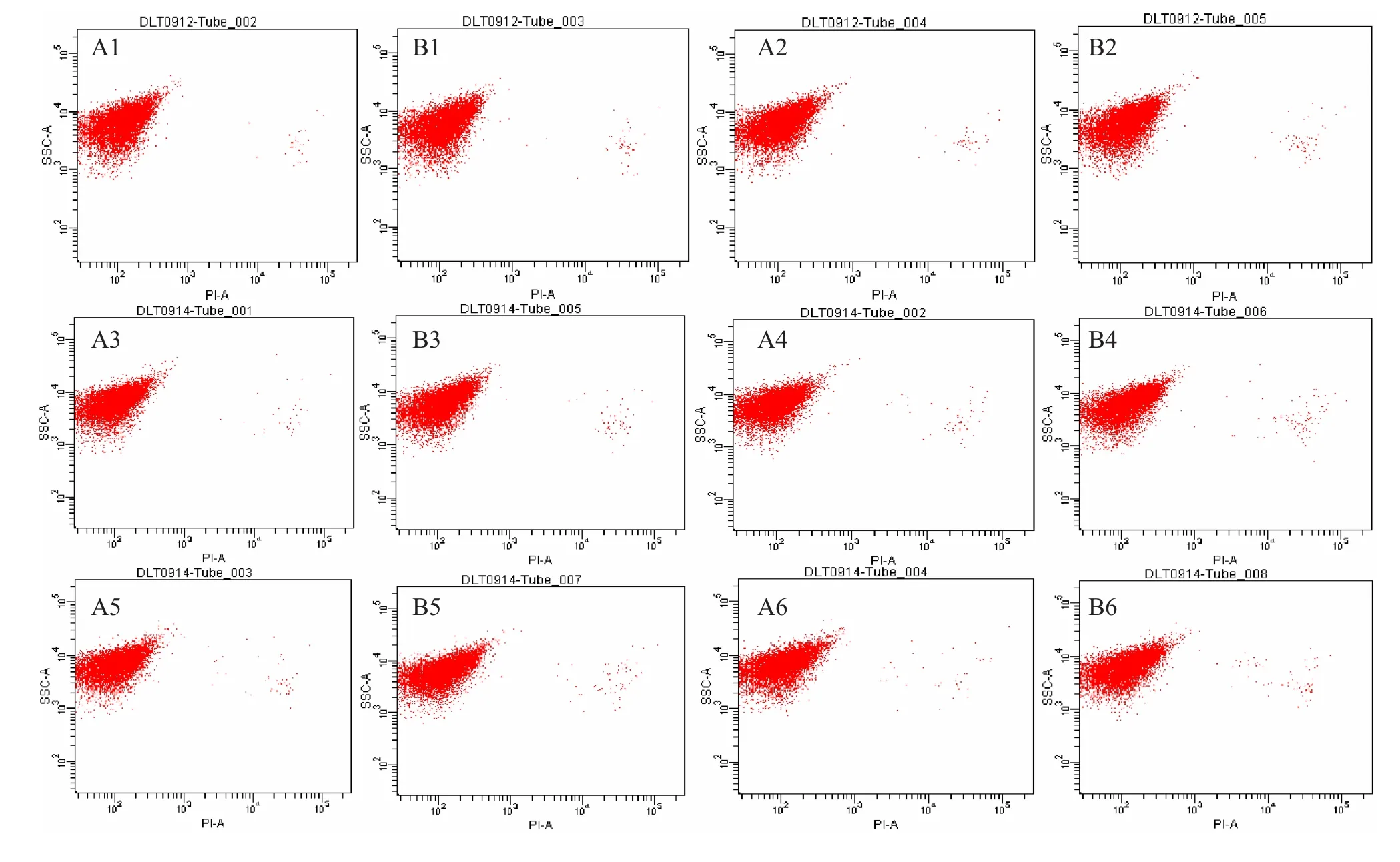
图6 不同时间段流式细胞术检测结果
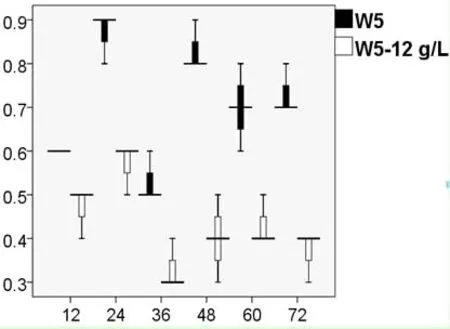
图7 流式细胞术检测结果
通过酿酒酵母代谢路径可知,乙醇、甘油和2,3-BD的合成途径成为酵母菌代谢的三个主要支路[22]。因此,外源添加适当乙偶姻后促使2,3-BD产量增加的同时,必定减少了乙醇或甘油合成过程中的碳源分配,这可能是因为添加适当乙偶姻后,促进了关键基因的表达,从而使2,3-BD的产量有所提高,由于乙偶姻和2,3-BD之间的反应是可逆的[23],所以当2,3-BD产量增加的同时也使丙酮酸到2,3-BD途径中相关酶的活性增加,所以,碳源在乙醇和甘油合成途径中减少的情况下,会导致最终的产量下降。同时,乙偶姻和2,3-BD的转化与NAD+与NADH之间的转化具有偶联关系[24]。Ehsani等[25]提出了增加供氧的策略,在敲除ilv2基因并过表达als、aldc和bdh1基因的S.cerevisiae中增加溶氧量,甘油积累减少的同时,葡萄糖合成2,3-BD的理论产量比厌氧条件下高20%。这说明通过改变溶氧调节氧化还原平衡对于生产2,3-BD具有重要意义。Kawai S等[26]将2-丁酮作为外源添加电子受体,使NADH的含量降低,使甘油的产量下降,但2,3-BD的产量明显提高。Kim S等[27]发现NADH氧化酶可以将NADH氧化成NAD+,为了降低NADH的浓度,在过表达als、aldc和bdh1的基因的S.cerevisiae过程菌株中外源表达乳酸乳球菌(Lactococcus lactis)中的NADH氧化酶,从而将甘油代谢流向2,3-BD,最终使2,3-BD的产量提高了23.8%。所以乙偶姻在被BDH催化生成2,3-BD的同时,也会受到NAD+与NADH含量的影响[28]。

图8 电镜观察结果
本研究将S.cerevisiaeW5/W5-Δpdc1/W5-Δpdc5/W141四株菌株进行试验对比,挑选出S.cerevisiaeW5/W141最适合作为出发菌株来生产2,3-BD,发现S.cerevisiaeW5和S.cerevisiaeW141外源添加乙偶姻的最适浓度为12 g/L和8 g/L,且当S.cerevisiaeW5最适添加量为12 g/L乙偶姻,且72 h时,2,3-BD的产量最大达到2.49±0.02 g/L,并且乙偶姻在发酵结束前消耗完全;S.cerevisiaeW141最适添加量为8 g/L乙偶姻,且72 h时,2,3-BD的产量最大达到1.56±0.04 g/L。当外源添加适量乙偶姻后,2,3-BD的产量之所以会提高,是因为在整个代谢过程中,乙偶姻与2,3-BD二者处于同一条代谢流中,且乙偶姻作为生产2,3-BD重要的前体物质,当乙偶姻的浓度升高时,同时也会伴随着最终产物的增加[29]。Brackman G等[30]利用异源过表达枯草芽孢杆菌的关键基因acoA和acoB,来增强乙醛转化为乙偶姻的能力,构建菌株进行分批补料发酵,最终2,3-BD的产量得到提升。Kim S等[31]在S.cerevisiae菌株的基础上,异源过表达B.subtilis中的alsS和alsD基因和内源bdh1基因后进行分批补料发酵,2,3-BD产量达到了29.1 g/L。理论上讲,在抑制乙醇代谢流向并过表达2,3-BD合成途径后,菌株可以利用葡萄糖经丙酮酸、α-乙酰乳酸、乙偶姻合成2,3-BD,或经丙酮酸和乙醛转化成乙偶姻,最终合成2,3-BD。因此,说明在代谢过程中,前体物质的增加对产物的产量具有一定促进作用。所以,本研究还可以做进一步的优化。通过代谢过程手段使工程菌株具备利用廉价原料(木质纤维素和藻类)来合成2,3-BD,从而提高生产2,3-BD的能力,但是,该方法是否会影响菌株正常生长,这些都是今后需要去研究的。综上所述,本研究中选用S.cerevisiaeW5/W141作为出发菌株,通过外源添加适量乙偶姻,使2,3-BD产量得到了一定的提升,从而解决了单一酿酒酵母发酵2,3-BD产量低的问题,也为微生物高产2,3-BD开辟了新的空间。
猜你喜欢
当代水产(2022年1期)2022-04-26
牡丹江医学院学报(2021年5期)2021-12-05
黑龙江大学自然科学学报(2021年4期)2021-11-19
食品安全导刊(2018年30期)2019-01-28
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
生物质化学工程(2014年4期)2014-07-05
食品工业科技(2014年13期)2014-03-11
食品科学(2013年24期)2013-03-11
食品与发酵工业(2011年2期)2011-12-18
