
茶树菇B交配型位点结构分析
2021-01-15 08:27陈卫民柴红梅赵永昌陈玉惠
中国农学通报 2021年2期
钱 蓉,陈卫民,柴红梅,陶 南,赵永昌,陈玉惠
(1西南林业大学生命科学学院,昆明 650224;2云南省农业科学院生物技术与种质资源研究所,昆明 650221;3云南省农业生物技术重点实验室,昆明 650223;4农业部西南作物基因资源与种质创制重点实验室,昆明 650223)
0 引言
担子菌中存在多种有性孢子,包括同核体和异核体,具有两个或多个异核体萌发的菌丝体能完成正常的有性生殖过程,而同核体结实是一种比较异类的生殖方式,曾在多种担子菌中此现象[1],包括暗绒盖多孔菌(Polyporus ciliatus)[2],光滑环锈伞(Pholiota nameko)[3],和裂褶菌(Schizophyllum commune)[4]等,金针菇(Flammulina velutipes)[5]产生的子实体多为流产型结实(Abortive homokaryotic fruiting,AHF)和真同核体结实(True homokaryotic fruiting,THF),前者(AHF)子实体不开伞也不产生孢子,而后者(THF)产生的子实体发育完全但较小,孢子梗上多着生2个孢子。有趣的是茶树菇(Agrocybe aegerita)中单核体能产生第3种子实体类型即假同核体结实(Pseudo homokaryotic fruiting,PHF),其初生菌丝具锁状联合,而且子实体发育完全,担子上着生4种交配型的孢子[6]。由此可见,担子菌产孢类型多变,容易混淆研究者对于物种遗传模式的认识。
根据已有的遗传模式研究,茶树菇有性生殖遗传分为两种类型。(1)典型四极性异宗结合型:研究分析了欧洲和日本的野生菌株交配型后认为茶树菇为多等位基因控制的四极性交配型[7-8]。(2)同核体交配极性转换型:从茶树菇中分离了4株野生型菌株子代,经锁状联合观察和交配极性测试,确认后的同核体菌株进行出菇分析,结果显示同核体菌株能产生3种类型的子实 体 AHF(Abortive homokaryotic fruiting)、THF(True homokaryotic fruiting)和PHF(Pseudo-homokaryotic fruiting)型,其中PHF型子实体的后代中产生两种新的交配型,后期的验证表明单核体菌株中进行了自发的极性转换[6]。单核体产生子实体的比例超过50%,个别菌株达到100%,THF和PHF的比率约为40%。经过子代交配型分析,研究者推断茶树菇交配极性转换菌株中A和B交配位点分别有3个,其中2个为沉默表达位点[6]。当二者通过位点转移转移,交配基因具有活性时,就会出现新的交配极性。由此可见,茶树菇单核体生殖是一种普遍现象,揭示其交配位点的基因结构组成及交配特征对于分析茶树菇交配极性转化具有重要的作用。
真菌的交配系统中,由单个遗传因子A决定的称为二极性交配系统,四极性交配系统则由不连锁的A、B双因子控制[11-12]。其中B因子编码信息素及其受体,控制着锁状细胞的融合和核迁移过程[13-14]。信息素及其受体蛋白,通过细胞膜上的G-蛋白偶联受体触发细胞内的多个生化反应,进而完成整个的生活史[15-17]。多级联的磷酸化反应最终将信号传递到末级的功能蛋白,控制着细胞融合等生理过程[18]。前期研究显示茶树菇近缘种杨柳田头菇Agrocybe salicacola中B交配位点具有5个受体基因和2~3个信息素前体编码基因,并且该位点被完整遗传给子代[9],该结果与前期研究中推测茶树菇中具有3个B交配位点的结果不同。由于前期研究中,研究者仅对茶树菇交配特点进行了系统研究,没有从交配位点分子组成进行进一步阐述。因而,本研究拟以茶树菇YSG菌株为研究材料,通过分离同核体菌株,进行基因组测序,验证分析其B交配位点,结合子代同核体交配基因分布和交配型分析,为茶树菇的有性生殖研究提供参考。
1 材料与方法
1.1 试验时间、地点
试验于2017年9月—2018年10月在云南省农业科学院生物技术与种质资源研究所进行。
1.2 试验材料
实验菌株茶树菇YSG菌株来源于江西省农业科学院,菌株属性见前文描述[10]。
1.3 同核菌株分离及菌株配对分析
单孢菌株分离采用孢子稀释涂布法,挑取的菌株转移到YPD培养基(酵母提取物2 g,蛋白胨2 g,葡萄糖20 g,琼脂15 g,蒸馏水1000 mL,pH值自然)上。培养2周根据菌落的整齐度初步筛选,选取边缘整齐菌株进行显微镜观察,并依据锁状结构判断菌株是否为同核体。获得的同核体菌株采用采用对峙培养的方法分析菌株交配极性。子实体形态切片后在显微镜下进行显微形态观察。
1.4 菌株培养及DNA提取
菌丝体置于YPD培养基上25℃培养2周后,刮取1 g菌丝液氮研磨后使用真菌DNA提取试剂盒(Biomega)提取基因组DNA。菌丝块接到无菌的木屑培养基(木屑80%,麦麸19%,石灰1%),置于22℃培养,用于出菇观察。
1.5 引物特征及PCR扩增
依据前期获得YSG同核体子代菌株YS8基因组注释结果,获得B交配位点,根据序列特征设计引物,用于验证序列。本试验所用引物的合成及PCR产物序列测定均在昆明硕擎生物科技有限公司完成。
2 结果与分析
2.1 YSG菌株子代同核出菇特征
同核体菌株接到木屑培养基后,进行了子代同核体出菇实验,结果显示,同核体能够形成不同类型的子实体,包括菌丝型(只有浓密的菌丝)、无盖型(无菌盖)和菌柄型(经过显微观察,菌盖不发育,不产生孢子)(图1)。其中的无盖型和菌柄型子实体在形成小的菇体后不再继续生长,之后就被菌丝覆盖后凋亡。
2.2 B交配位点结构组成分析
选择YSG菌株子代同核体菌株YS8(交配型A1B1)进行基因组测序及组装。根据基因组注释信息,并设计5对嵌套引物YS8RCB1f/1r-YS8RCB5f/5r进行扩增,测序验证B交配位点序列的准确性(表1)。结果显示,YS8菌株的B交配位点分别被组装到2个Scaffold上,包括4个完整的信息素受体和1个信息素前体编码基因(图2A)。
进一步的序列分析发现,以上2个Scaffold上靠近信息素受体基因B2-rcb2和B2-rcb4区域,分别有一个片段被注释为信息素受体编码基因,二者(分别命名为B2-rcb3a和B2-rcb3b)的下游和上游分别约有30 kb和20 kb以上的区域内没有信息素受体基因存在。并且紧邻区域部分序列反向互补(图2B)。将2个片段拼接到一起后,能够组成完整的信息素受体编码基因(图2C)。
2.3 同核体子代基因型及交配型分析
根据2个片段B2-rcb3a和B2-rcb3b序列,分别设计引物对P3af/r和P3bf/r(表2),用来分析同核体群体子代基因型。扩增结果现实,2对引物将所有的同核体菌株分为2种类型,并且二者的扩增谱相同(图3A),推测连接B2-rcb3a和B2-rcb3b的大片段区域,在遗传到子代的过程中没有发生重组。
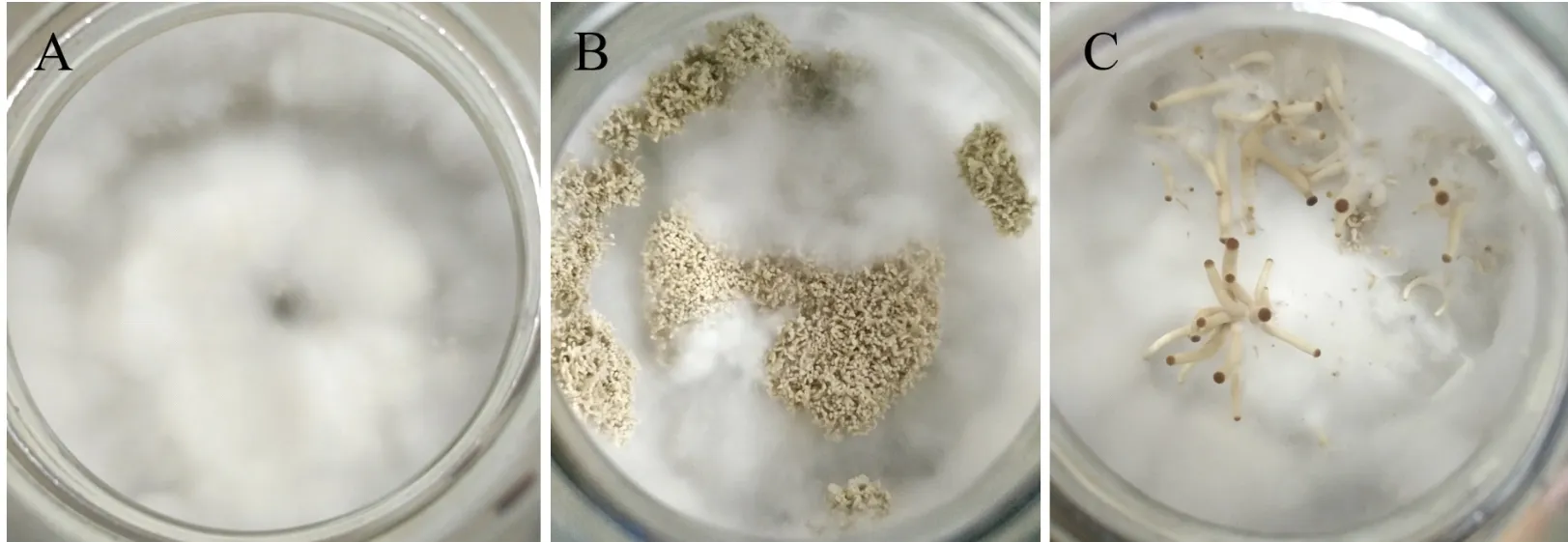
图1 同核体菌株出菇特征
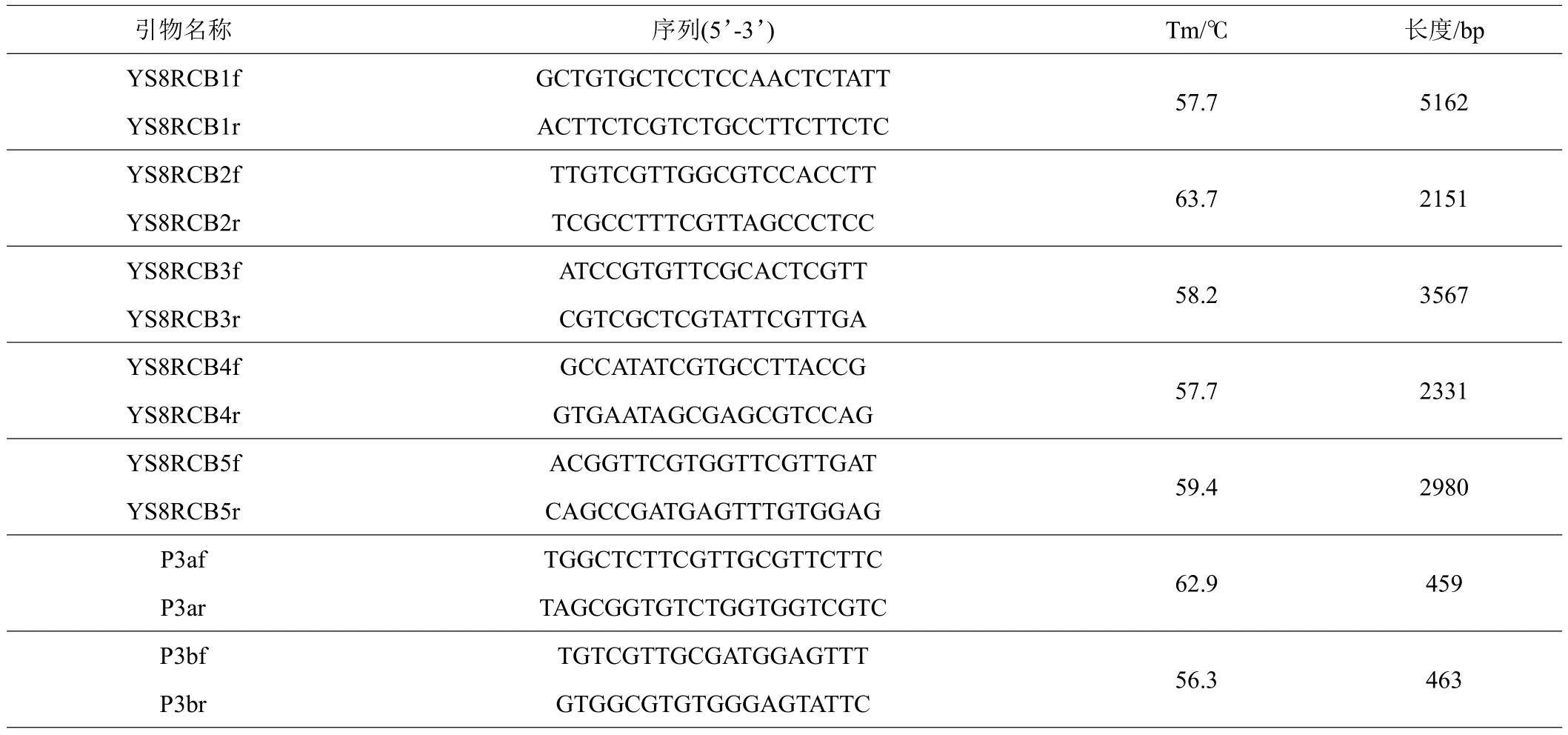
表1 YS8菌株B交配位点验证及交配型鉴定引物特征
交配型分析结果显示,所用的同核体菌株群体只有2种B交配型,分别为b1和b2型(图2B)。交配型与B交配基因的基因型呈现完全对应关系,从而也进一步表明了B交配位点决定了菌株的交配型,并且该位点被完整分配到子代中。
3 讨论
明确真菌的有性生殖模式和交配因子组成,对于其生殖遗传研究、子实体发育,以及育种研究都具有重要意义。交配位点的基因组成决定了菌株的交配特点,探究真菌物种的有性生殖特征,需要明确其交配位点结构组成。有关B位点基因遗传结构组成的研究较多,其组成也更为复杂,被认为是真菌在野生状态下提高交配成功率的一种生存策略[12,19]。灰盖鬼伞Coprinus cinereus B交配位点有3个基因簇,每个由3个基因组成,负责编码2个信息素和1个受体[19]。随着基因组学的广泛应用,更多真菌的基因组得以快速解析。香菇Lentinula edodes B交配位点内有4个受体编码基因及两个信息素前体,并且在单核体中B位点结构基本一致[21-22]。金针菇包含有两个亚位点,分别有一个信息素受体和1~2个信息素前体编码基因[23]。草菇Volvariella volvacea中具有4个信息素受体编码基因[24],但其B交配位点可能没有活性[25]。本研究中所确定的茶树菇YSG菌株B交配位点包含有5个信息素受体和1个信息素前体编码基因,与之前报道的近缘种杨柳田头菇相似[9]。由此可见,在明确不同物种中交配基因的组成,以及多个信息素及受体在同核体菌株交配过程中的作用,才能探究交配极性转换的机制。但是由于目前对交配基因功能的认识不足,因而对茶树菇B位点结构组成的命名比较模糊,如亚位点的划分只是根据信息素及其受体编码基因簇的聚集度,即它们在基因组物理图谱上的距离来进行,而不是依据其交配功能来划分。茶树菇B交配位点信息素和其受体在基因上距离较小,似乎只能认定为一个位点,判断比较困难。以前报道中所认为的茶树菇应有3个亚位点[6],笔者建议这些亚位点应该描述为功能基因,即它们决定交配功能角色的转变导致了新的交配极性的产生。另外,通过茶树菇YSG同核体子代B交配位点和交配型分析,明确了其决定菌株交配型,并且在遗传给子代的过程中,B位点内部没有重组事件发生。该结果表明了该位点组成发生改变的几率较小,即使如本研究中发现的大片段插入也没有改变该位点整体遗传到子代现象出现。在本研究中,分离了大量的同核体,培养结果只获得了部分畸形(或不产孢型)子实体,而并未得到正常子产孢型实体,这与文献报道中高比率产孢同核体相比明显不同,结果的差异是否由于类群差异造成,由于茶树菇类群较多[26-28],还需要收集更多的资源,并根据类群划分进行同核体出菇分析。
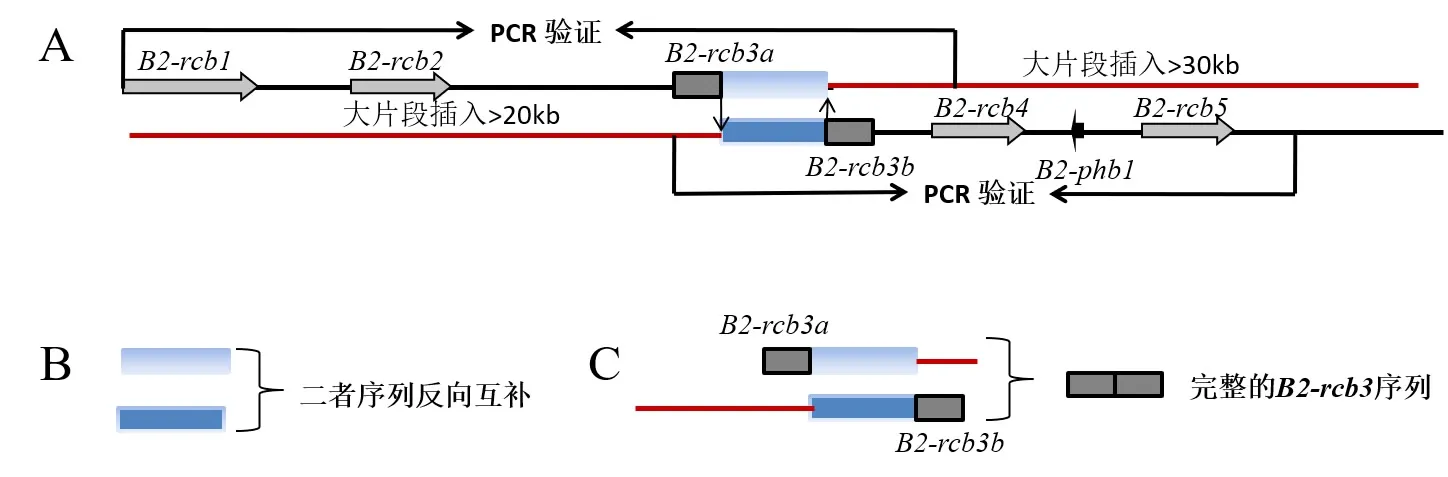
图2 B交配位点结构特征
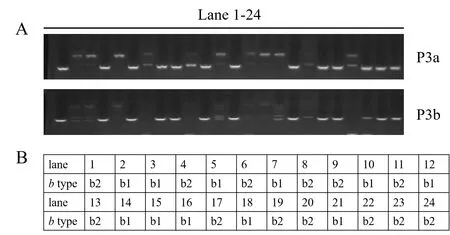
图3 同核体子代B交配位点基因型及交配型
4 结论
茶树菇YSG子代同核体培养后,部分菌株能产生子实体,但缺少菌盖或有菌盖但无担子,即产生的畸形子实体没有经过有性生殖过程。根据基因组序列,获得了YSG子代同核体YS8菌株B交配位点信息,经嵌套PCR验证确认B交配位点包含有5个信息素受体和一个信息素前体编码基因。其中B2-rcb3基因序列中有一个大于50 kb的插入片段,该片段中有部分序列为反向重复。经过子代同核体PCR扩增谱分析,表明B交配位点,包括该大的插入片段并未在有性生殖过程中发生重组,而是完整的遗传到子代中。交配极性分析结果显示,群体中只有2种B交配型,24个菌株中没有新的B交配型产生,并且也印证了B交配位点完整遗传到子代的特征。这些结果与以前报道的茶树菇交配极性转换结果不同,有助于茶树菇生殖遗传多样性及育种工作研究。
猜你喜欢
四川蚕业(2021年1期)2021-02-12
食用菌(2020年2期)2020-12-22
昆虫学报(2020年6期)2020-08-06
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
广西林业科学(2016年4期)2016-03-16
广西林业科学(2016年3期)2016-03-16
中学生物学(2016年8期)2016-01-18
中国马铃薯(2015年3期)2015-12-19
读者·原创版(2014年12期)2014-01-16
